











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
The knowledge from the morphology of seeds allows the identification of different structures developed for geographic dispersion (Niembro, 1988). It has been documented that the individuals which allocate a greater energy investment into structures that maximize seed dispersal will be positively selected (Augspurger and Hogan 1983) and suitable for establishment (Sánchez-Salas et al. 2013).
One strategy of seed dispersion which is known as hydrochory, meaning that the dispersion of seeds occurs through water (Sánchez-Salas et al. 2012 y 2014). This process is endorsed by the presence of structures that have a low specific weight, due to the presence of slots filled with air, spongy tissue or special corky tissue (Moreno 2009; Hendry and Grime 1993). Structural characteristics, such as a large hilum (as is displayed in Astrophytum coahuilense seeds) plays an important role in the dispersion of seeds by means of water action (Bregman 1988).
In this way, is mention that there are seven main types of seed shapes that have been described for the subfamily Cactoideae, drawn from their structures and shape: Cereus-Type, Rhipsalis-Type, Arequipa-Type, Notocactus-Type, Thrixanthocereus-Type, Astrophytum-Type and Blossfeldia-Type, all of which have evolved from a Pereskioid-Type ancestor with a suborbicular-lenticular seed shape. Astrophytum-Type seeds are navicular and adapted especially for hydrochory, but can also be soared by wind (anemochory) which denotes an evolutionarily advanced form.
Astrophytum coahuilense (considered a different species to A. myriostigma según Vazquez-Lobo et al. 2015) is an endemic cactus species native to Mexico and it is currently in vulnerable (VU) category according to the IUCN (2011), mainly due to systematic extraction and habitat destruction in areas where it grows naturally. Various studies propose that the seed of this species is hydrochorous and that it is dispersed by means of water currents (Bregman 1988; Barthlott and Hunt 2000; Sánchez-Salas et al. 2012, 2013, 2014), which has probably provided a wide range of dispersion to the species along the northern Mexican Chihuahuan desert, considering that in the past (Pleistocene-Holocene) there were abundant paleo-lakes and paleo-rivers in the north of Mexico (Van Devender y Burgess, 1977, 1985; y Marín y Torres-Ruata, 1990; Metcalfe 2006; Czaja et al., 2014). However, there has not been any analysis carried out to show that the structure of the seed together with its density, as adaptations of its empty chambers and spongy tissue, confer buoyancy and to answer the question: why floats the seed of A. coahuilense? In this study, the buoyancy of the seed of this species was analyzed to identify the structures that confer the ability to float. The mass and volume of the whole seeds were calculated with the assumption that the seed density is less than that of water. This work is intended to contribute to future explication the of A. coahuilense distribution pattern with reference to the other five species of the genus.
Methods
During September of 2007, 40 individuals of A. coahuilense were georeferenced from "Las Palmas" locality at El Sarnoso, Gómez Palacio, Durango. 10 individuals were randomly selected, with the aim of disrupting at least the seed bank of this population, which has been highly impacted by anthropogenic activities. Two fallen fruits were collected from each selected individual, whereas only near the ground fruits, which were deposited in paper bag, taking into account the specifications from Gold et al. (2004) for its collection. Of 3000 seeds, 50 seeds were random selected of the fallen fruits colleted and stored in paper bags (Arredondo-Gómez and Camacho-Morfín, 1995) during one week approximately. In order to identify the structures that confer the buoyancy of the A. coahuilense seed, a transversal (freehand) gash using a cutter was performed to 25 randomly selected seeds. This treatment favored the separation of the large or empty chamber from the embryo chamber (Becerra López et al. 2010). We work with five structures like experimental unit and four repetitions (five repetitons in total by structures). Both chambers were divided into subgroups of five and placed in polyethylene vessels containing 30 ml of distilled water (five glasses per chamber type). Five subgroups of five complete seeds were also used. The vessels were shaken for 10 seconds every two hours for 24 hours (temperature 18°-25°C) to simulate the movement of water currents, because the natural current of region of La Comarca Lagunera its turbulents. Although, Van Den Broek et al. (2005) proved that the movement of water in buoyancy tests is not significant. At the end of this period seeds and structures that remained afloat and those sunk for each vessel were counted and thus we could able an error rate for each experimental unit. In order to identify associations between the type of structure and buoyancy a chi-square test was performed in a contingency table (2x3). The non-parametric Kruskal-Wallis variance analysis determined floated seed differences by type of structure employing a Post hoc test (ranks test) and difference between groups were identified. The statistical analyzes were executed in the program MedCalc® Version 12.5.0.0, 1993-2013. Statistical tests were assumed to be significant when P ≤ 0.05.
To determine the density of the seed, a sample of 50 randomly selected seeds mass and volume were calculated using a pycnometer and an analytical balance (ADVENTURE) with accurateness of 0.0001 g, and then the density of the seed was calculated. In order to calculate mass, volume and density of the seeds, Querelle y Cia Ltda (2015) procedure was used, where the density of a body (g/cm3) is given by:
Where: p = density; m = mass and V = volume
For which the following measures were calculated:
a) Weight of the seeds (Pc), b) Weight of the pycnometer filled with water (Pp) and c) Weight of the pycnometer containing submerged seeds (Pt).
To calculate the mass of each component (Pc, Pp, Pt), values were divided by the gravity force ≈ 9.8 m s2, to obtain their mass values, where:
m c = mass of the seeds, m p = mass of the pycnometer with water, m t = mass of the pycnometer containing the submerged seeds.
The Archimedes principle says that all body immersed in a fluid experiences a vertical and upwards thrust equal to the weight of fluid buoyed which is called buoyancy (Mohazzabi, 2017). So all body with a density less than water, floats (sufficient that the density of the body is less than the density of water so the body float).
Then by the principle of Archimedes we must:
Where:m t = mass of the pycnometer containing the submerged seeds.
m p = mass of the pycnometer with water
ρH2O = is the water density ≈ 1 g/cm3
V c = is the volume of the seeds
m c = mass of the seeds
Taking into account that:
Where ρc is the density of the seeds, we can substitute it in (2) and write:
Results
It was observed that the seed of A. coahuilense has two chambers, the large or empty chamber (hilum) and a small chamber or embryo chamber, which is the seed itself. The largest amount of spongy tissue was found in the large chamber (1). A significant association among type of structure and buoyancy capacity was determined (x2 = 31.59; D.F. = 2, P = 0.001). Most of the empty chambers (84%) and complete seeds (88%) floated, while most chambers of the embryo (80%) collapsed (table 1). The Kruskal-Wallis test showed a highly significant difference (Ht = 16.00, D.F. = 2, P = 0.0048) among floatability of the empty chamber, whole seed and embryo chamber (table 2). Multiple median comparison tests showed that the embryo chamber has a lower capacity to float than the empty chamber and complete seed (figure 1, 2 and table 2). A. coahuilense seeds displayed an average density of (0.213 g/cm3), mass of 0.0143 g and volume of 0.0672 mm3.
Table 1 Frequency and percent for floatability and sinking of A. coahuilense seeds are displayed. Small chamber seeds (SCS), large chamber seeds (LCS) and complete seeds (CS) were considered. Complete seeds (CS) showed the highest floatability and the lowest sinking proportion.
| SCS | LCS | CS | |
|---|---|---|---|
| A float seeds | 20% (5) | 84% (21) | 88% (22) |
| Sunk seeds | 80% (20) | 16% (4) | 12% (3) |
Table 2 Test for Floatability of A. coahuilense seeds are displayed. Empty chamber, Embryo chamber and Complete Seeds were considered. Complete seeds showed the highest flotability and the embryo chamber the highest sinking proportion.
| Data | Floatability | ||
| Factor codes | Groups | ||
| Test statistic | 9.9950 | ||
| Corrected for ties Ht | 10.6613 | ||
| Degrees of Freedom (DF) | 2 | ||
| Significance level | P = 0.0048 | ||
| Posthoc test analysis Factor | n | Average Rank | Different (P<0.05) from factor nr |
| (1) Complete seed | 5 | 12.70 | (2)(3) |
| (2) Embryo chamber | 5 | 3.80 | (1)(3) |
| (3) Empty chamber | 5 | 7.50 | (1)(2) |
| Sample size | 15 (five repetition for treatment) | ||

Fig. 1 Statistical median dispersion values for buoyancy (understood as the number of floating seeds) of the different seed structures evaluated.
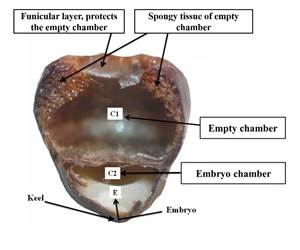
Fig. 2 Freehand seed cutting, where the double chamber of the A. coahuilense seed is observed. The upper part shows the empty chamber (C1) surrounded by spongy tissue, while in the lower part of the seed is the embryo chamber (C2), the whitish structure is the endosperm and in the lower part of the endosperm is the region called keel, where the embryo (E) is located forming part of the embryo chamber (C2). Photo Jaime Sánchez.
Discussion
It has previously been denoted that the seeds from many plants are adapted to float because they contain spongy tissue that include large compartments which harbor air (Raven and Evert, 1999; Hendry and Grime, 1993). These characteristics fall in agreement with the spongy tissue features from the seminal cover found in the empty chamber (hilum) from A. coahuilense seed and the association between the type of structure and the buoyancy capacity. In the present study it is suggested that the presence of both spongy tissue and empty chamber could perform as a buoy and their conjointment is what gives the seed the ability to float and consequently the hydrochoric capacity. Bregman's work (1988) confirms that the seeds with superior buoyancy abilities are those with a considerably large hilum and a little or absent funicular tissue, a very thin covering and a small embryo as in the seeds from the Astrophytum genus. Barthlott and Hunt (2000) support this mentioning that Astrophytum seeds are navicular and specially adapted for hydrochory, but can also be dispersed by the wind (anemochoric). Sánchez-Salas et al. (2014) also found that 70% of A. capricorne seeds showed buoyancy for more than 30 days and that most of the seedlings that developed on the water during the evaluation, also floated. The probable adaptation for dispersion by water currents in A. coahuilense seeds could be considered to explain the geographical range of this plant, which in turn borders the margins of the great Irritila paleolake (Czaja et al., 2014). Taking into account that this area was covered by extensive paleolakes and paleorivers in the past (Van Devender, 1977; Van Devender y Burgess, 1985; Metcalfe, 2006, Marín y Torres-Ruata, 1990) that could have served as a mechanism of dispersion and probably as an agent for deposition of seeds on the limestone hills with rosette vegetation, characteristics where its main nurse Agave lecheguilla inhabits (Muro-Pérez et al., 2009). It is very important, because for Bregman (1988), the distribution pattern is precisely one of the plant characteristics that must be taken into account to support seed hydrochory. On the other hand, experimental designs that integrate water flows where the seeds are deposited are needed, and verify its dispersion. In the future, we also need to compare the buoyancy and geographical distribution of this seed with other species of the desert to have more evidence that a large percentage of buoyancy of seeds may be associated with certain structures and wide geographical distribution. Sánchez-Salas et al. (2014) analyzed seed buoyancy in A. capricorne and A. ornatum, which belong to the same genus and have a very similar to A. coahuilense seed. Hydrochory was demonstrated through buoyancy tests and morphological analysis, it was concluded that both seeds have adaptations for hydro-dispersion. In this work the hydrochorous evidences were focused on the buoyancy analysis and the density trial of A. coahuilense seed (0.213 g/cm3) in contrast to water density (1 g/cm3), on the basis of Archimedes' theory, in which a body can float if its density is inferior than the volume of water it displaces. Considering the morphological characteristics such as the spongy tissue found in the empty chamber and shape of an empty buoy itself, density as a concept closely related to the buoyancy of the seed in water, that the navicular shape (of a ship or boat) is also associated with seed buoyancy (Bregman 1988) and seed buoyancy being closely linked to the hydrochory (Farji-Brener et al., 2005).

