











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
El fuego es un elemento de perturbación presente en varios ecosistemas terrestres y sus efectos se relacionan directamente con la estructura de los ecosistemas y composición de especies (Agee, 1993; Pyne et al., 1996; Rodríguez-Trejo, 2008; Jardel-Peláez et al., 2009; Ávila-Flores et al., 2014). Dependiendo del régimen de incendios, las especies tienden a responder de manera diferente ante un evento de perturbación, esto es, las especies son adaptadas y sensibles al fuego. Parte de la adaptación es presentar características de resistencia al fuego y supervivencia al mismo (Agee, 1993; Rodríguez-Trejo, 2008). En México, a pesar de constituir la base para el manejo sustentable de los recursos naturales (Guariguata, 1998), han sido escasos los estudios que evalúen la diversidad y composición de especies del matorral espinoso tamaulipeco después de un incendio (Moreno, 2013).
Entre muchos otros aspectos, el régimen de fuego es referido a la frecuencia, extensión, intensidad y severidad de los incendios en un área específica (Heinselman, 1981; Agee, 1993). La frecuencia se refiere al tiempo que tarde en volver a presentarse un incendio en el mismo lugar, la extensión es el tamaño promedio de los incendios en dichas condiciones y la severidad es referida al daño propiciado al ecosistema, la intensidad se caracteriza por la cantidad de energía liberada, mientras que severidad se mide principalmente en proporción de daño al arbolado (mortalidad), al suelo y biodiversidad. (Agee, 1993; Rodríguez-Trejo, 1996). Se han propuesto algunas clasificaciones para el régimen potencial de fuego para México (Jardel-Peláez et al., 2009; Rodríguez Trejo, 2008). El matorral del noreste de México, de manera generalizada y por sus características bioclimáticas y físicas, potencialmente presenta incendios con baja frecuencia, severidad baja-moderada con tamaño pequeño-mediano (Jardel-Peláez et al., 2009).
El matorral espinoso tamaulipeco (MET) cubre una superficie de 200 000 km² del noreste de México y del sur de Texas, de la Llera de Canales y los límites sureños de la Sierra Azul en Tamaulipas (González, 1985) al altiplano Edwards en Texas (Diamond et al., 1987) y de las faldas de la Sierra Madre Oriental hasta la costa del Golfo de México (Jurado y Reid, 1989). Este ecosistema es bastante diverso respecto a las especies arbóreas, arbustivas y subarbustivas, densidad y altura. También es de gran importancia en la economía del noreste de México desde fines del siglo XVI (Alanís, 2006).
En el MET se han realizado diversos estudios científicos que evalúan la regeneración de las comunidades vegetales del matorral espinoso tamaulipeco después de diferentes perturbaciones antropogénicas (Pequeño et al., 2012; Alanís et al., 2013; Jiménez et al., 2013; Martínez et al., 2014). Sin embargo, se carece de información sobre el efecto del fuego sobre la regeneración de este ecosistema.
Este estudio tiene como objetivo principal, caracterizar la estructura y diversidad de especies después de un incendio en el matorral espinoso tamaulipeco (MET), de tal forma que contribuya al conocimiento de la dinámica “sucesional” de este tipo de ecosistema. En específico, las preguntas de la investigación son: 1) ¿Que especies están presentes después de un incendio? 2) ¿Cuáles son las especies que resisten el fuego? 3) ¿Qué capacidad de resistencia presenta el ecosistema? Y finalmente, 4) ¿Cuán diverso es el ecosistema ante un incendio?
Métodos
Área de estudio
El estudio se llevó a cabo en la localidad de Estación Huertas, Municipio de Montemorelos, Nuevo León (noreste de México) a 82 km al sureste de la ciudad de Monterrey (fig. 1). Se localiza en las coordenadas 25°1′48″ de latitud norte y 99°45′47″ de longitud oeste, con una altitud de 510 m s.n.m.

Toma de datos en campo
En el verano del año 2010 se presentó un incendio superficial y de copa que afectó a los elementos vegetales. El incendio fue de severidad media y ocurrió en una superficie de 3 600 hectáreas. Se incineró la parte aérea y se presentó escasa sobrevivencia de los árboles y arbustos, con excepción de la especie Yucca filifera, la cual por su altura no sufrió daño en las hojas. Después del incendio no se practicó ninguna actividad productiva ni se realizaron actividades de restauración ecológica, dando oportunidad a que la comunidad vegetal se regenerara naturalmente.
En el verano del 2014 se establecieron aleatoriamente cinco sitios de muestreo cuadrangulares para evaluar la vegetación, la dimensión de cada uno fue de 40 × 40 m (1 600 m2). Los sitios se establecieron en áreas con características ecológicas similares (altitud, exposición, tipo de suelo, asociación de especies, etc.), en un paraje de 50 hectáreas del ejido “Estación Huertas” y en donde el incendio se presentó de forma homogénea. El paraje se encuentra delimitado por un cerco perimetral, donde se tiene la certeza que no se han desarrollado actividades productivas ni de restauración. La forma cuadrangular de los sitios se utilizó debido a su facilidad de delimitación y medición en vegetación densa, con respecto a la forma circular (Alanís et al., 2008). En los sitios de muestro se realizó un censo de todas las especies arbóreas y arbustivas (sobrevivientes y regeneradas) con un diámetro mayor a cinco cm a una altura de 10 cm (d 0.10 m > 5 cm) sobre la base del suelo debido a las bifurcaciones que presentaron los individuos y para obtener mayor representatividad (Mora et al., 2014). Además, que de cada individuo se registró el diámetro de copa (d copa ) y fue identificado por personal del laboratorio de botánica de la Facultad de Ciencias Forestales de la UANL.
Análisis de los datos
Para cada especie se determinó su abundancia, de acuerdo con el número de árboles, su dominancia, en función del área de copa, y su frecuencia con base en su presencia en los sitios de muestreo. Estos valores se utilizaron para obtener un valor ponderado a nivel de especie denominado índice de valor de importancia (IVI), que adquiere valores porcentuales en una escala de 0 a 100 (Mostacedo y Fredericksen, 2000). Las fórmulas para estos cálculos se describen a continuación.
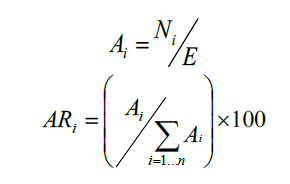
donde A i es la abundancia absoluta, AR i es la abundancia relativa de la especie i respecto a la abundancia total, N i es el número de individuos de la especie i, y E la superficie de muestreo (ha).
La cobertura relativa se evaluó mediante:

donde D i es la cobertura absoluta, DR i es cobertura relativa de la especie i respecto a la cobertura, Ab el área de copa de la especie i y E la superficie (ha).
La frecuencia relativa se obtuvo con la siguiente ecuación:

donde F i es la frecuencia absoluta, FR i es la frecuencia relativa de la especie i respecto a la suma de las frecuencias, P i es el número de sitios en el que está presente la especie i y NS el número total de sitios de muestreo.
El índice de valor de importancia (IVI) se define como (Whittaker, 1972; Moreno, 2001):

Para estimar la riqueza de especies se utilizó el índice de Margalef (D Mg ) y para la diversidad alfa el índice de Shannon-Weaver (H´) mediante las ecuaciones (Shannon, 1948; Magurran, 2004):
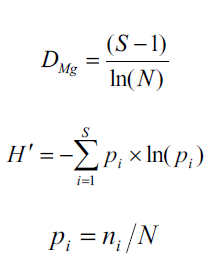
donde S es el número de especies presentes, N es el número total de individuos, n i es el número de individuos de la especie i y p i es la proporción de individuos de la especie i respecto al total de individuos (es decir la abundancia relativa de la especiei).
Resultados y discusión
Se registraron 1 448 individuos por hectárea (Naha-1) distribuidos en 14 familias, 22 géneros y 24 especies (cuadro 1). Las familias Fabaceae y Rutaceae son las que registraron más especies con seis y tres respectivamente, seguidas de las familias Euphorbiaceae, Rhamnaceae y Ebenaceae con dos especies cada una; para las familias restantes sólo se registró una especie. La mayor abundancia de las plantas de la familia Fabaceae se debe a que en el estado de Nuevo León algunas especies son abundantes sobre grandes extensiones, como Acacia rigidula, A. constricta, A. berlandieri y Prosopis glandulosa, asociados a diversas comunidades de matorral xerófilo (González et al., 1997; Estrada y Jurado, 2005) con vegetación secundaria. González et al. (2010) reportaron 10 especies de la familia Fabaceae al evaluar la composición y estructura de la vegetación en tres sitios del estado de Nuevo León, México. Según Domínguez et al. (2013) el desarrollo de la familia Fabaceae, se asocia a su capacidad de establecimiento en condiciones ambientales adversas.
Cuadro 1 Nombre científico y común, familias y forma de crecimiento de las especies registradas (ordenadas alfabéticamente).
| Nombre científico (TROPICOS®) | Nombre común | Familia | Forma de crecimiento |
|---|---|---|---|
| Acacia amentacea (L.) Sarg. | chaparro prieto | Fabaceae | arbustiva |
| Acacia farnesiana (L.) Willd | huizache | Fabaceae | arbustiva |
| Amyris texana (Buckley) P. Wilson | chapotillo | Rutaceae | arbustiva |
| Bernardia myricifolia (Sheele) Wats. | oreja de ratón | Euphrobiaceae | arbustiva |
| Celtis pallida Torr. | granjeno | Ulmaceae | arbustiva |
| Citharexylon berlandieri B.L. Robinson | encorvagallina | Verbenaceae | arbórea |
| Condalia hookeri M. C. Johnst. | brasil | Rhamnaceae | arbórea |
| Cordia boissieri A. D.C. | anacahuita | Boraginaceae | arbórea |
| Croton suaveolens Torr. | salvia | Euphorbiaceae | arbustiva |
| Diospyros palmeri Eastw | chapote amarillo | Ebenaceae | arbórea |
| Diospyros texana Scheele. | chapote blanco | Ebenaceae | arbórea |
| Ebenopsis ebano (Berl.) Britton et Rose | ébano | Fabaceae | arbórea |
| Forestiera angustifolia Torr. | panalero | Oleaceae | arbustiva |
| Havardia pallens (Benth) Britt y Rose | tenaza | Fabaceae | arbórea |
| Helietta parvifolia (Gray) Benth. | barreta | Rutaceae | arbustiva |
| karwinskia humboldtiana (Schult.) Zucc. | coyotillo | Rhamnaceae | arbustiva |
| Leucophyllum frutescens (Berl.) I. M. Johnst | cenizo | Scrophulariaceae | arbustiva |
| Mimosa biuncifera Benth. | uña de gato | Fabaceae | arbustiva |
| Neopringlea integrifolia (Hemsley) S. Watson | corvagallina | Flacourtiaceae | arbórea |
| Parkinsonia aculeata L. | retama | Caesalpiniaceae | arbórea |
| Prosopis laevigata (Humb. y Bonpl. ex Willd.) M. C. Johnston | mezquite | Fabaceae | arbórea |
| Randia rhagocarpa Standl. | cruceto | Rubiaceae | arbustiva |
| Rhus virens Lindh. ex A.Gray | lantristrisco | Anacardiaceae | arbustiva |
| Zanthoxylum fagara (L.) Sarg. | colima | Rutaceae | arbustiva |
Parámetros ecológicos
Abundancia
La abundancia se mide de acuerdo con el número de individuos por unidad de superficie, para el caso de esta comunidad vegetal se presentó una abundancia de 1 448 árboles y arbustos por hectárea (Naha-1). Esta abundancia es inferior a la reportada por Mora et al. (2013) y Jiménez et al. (2012), quienes registraron 1 792 Naha-1 y 2 370 Naha-1 respectivamente en un área de matorral espinoso tamaulipeco con historial pecuario. Alanís et al., (2011) registraron una abundancia de 2 876 Naha-1 al caracterizar de regeneración leñosa postincendio en una zona de transición entre el bosque de pino-encino y matorral en el Parque Ecológico Chipinque, México. Referente al número de individuos por hectárea, seis especies fueron las más abundantes, presentando 1 038 Naha-1 que representa el 72% de la comunidad. La especie Leucophyllum frutescens fue la más abundante con 248 Naha-1, seguida por Havardia pallens con 185 Naha-1 y Mimosa zygophilla con 160 Naha-1 (cuadro 2).
Cuadro 2 Abundancia, dominancia, frecuencia e índice de valor de importancia de las especies registradas.
| Especie | Abundancia | Dominancia | Frecuencia | IVI | |||
|---|---|---|---|---|---|---|---|
| Abs | Rel | Abs | Rel | Abs | Rel | ||
| Naha -1 | % | m 2 /ha | % | N/Sitios | % | ||
| Havardia pallens | 185 | 12.8 | 936.1 | 14.8 | 5 | 6.5 | 11.3 |
| Leucophyllum frutescens | 248 | 17.1 | 534.1 | 8.4 | 4 | 5.2 | 10.2 |
| Cordia boissieri | 96 | 6.6 | 1062.4 | 16.7 | 5 | 6.5 | 10.0 |
| Neopringlea integrifolia | 147 | 10.2 | 852.8 | 13.4 | 3 | 3.9 | 9.2 |
| Zanthoxylum fagara | 150 | 10.4 | 591.6 | 9.3 | 5 | 6.5 | 8.7 |
| Mimosa zygophilla | 160 | 11.0 | 542.6 | 8.6 | 5 | 6.5 | 8.7 |
| Ebenopsis ebano | 148 | 10.2 | 376.5 | 5.9 | 4 | 5.2 | 7.1 |
| Celtis pallida | 57 | 3.9 | 340.6 | 5.4 | 5 | 6.5 | 5.3 |
| Croton suaveolens | 74 | 5.1 | 263.9 | 4.2 | 5 | 6.5 | 5.3 |
| Forestiera angustifolia | 36 | 2.5 | 110.1 | 1.7 | 5 | 6.5 | 3.6 |
| Acacia amentacea | 21 | 1.5 | 214.6 | 3.4 | 4 | 5.2 | 3.3 |
| karwinskia humboldtiana | 35 | 2.4 | 104.0 | 1.6 | 3 | 3.9 | 2.7 |
| Bernardia myricifolia | 17 | 1.2 | 40.9 | 0.6 | 4 | 5.2 | 2.3 |
| Diospyros palmeri | 10 | 0.7 | 112.1 | 1.8 | 3 | 3.9 | 2.1 |
| Helietta parvifolia | 14 | 1.0 | 89.1 | 1.4 | 3 | 3.9 | 2.1 |
| Parkinsonia aculeata | 9 | 0.6 | 21.8 | 0.3 | 3 | 3.9 | 1.6 |
| Randia rhagocarpa | 8 | 0.6 | 19.4 | 0.3 | 3 | 3.9 | 1.6 |
| Amyris texana | 4 | 0.3 | 32.7 | 0.5 | 2 | 2.6 | 1.1 |
| Acacia farnesiana | 17 | 1.2 | 35.0 | 0.6 | 1 | 1.3 | 1.0 |
| Diospyros texana | 4 | 0.3 | 39.0 | 0.6 | 1 | 1.3 | 0.7 |
| Rhus virens | 4 | 0.3 | 9.5 | 0.1 | 1 | 1.3 | 0.6 |
| Citharexylon berlandieri | 1 | 0.1 | 9.4 | 0.1 | 1 | 1.3 | 0.5 |
| Condalia hookeri | 2 | 0.1 | 4.0 | 0.1 | 1 | 1.3 | 0.5 |
| Prosopis laevigata | 1 | 0.1 | 1.53 | 0.02 | 1 | 1.3 | 0.5 |
| 1448 | 100 | 6343.62 | 100 | 77 | 100 | 100 | |
Dónde: Abs = valores absolutos; Rel = valores relativos (%); Naha -1 = número de árboles por hectárea; IVIrel = índice de valores de importancia relativos (%).
La curva de dominancia-diversidad describe de forma gráfica la relación entre la abundancia y las especies ordenadas en categorías de la más a la menos abundante (Villareal et al., 2006). La línea de tendencia de la distribución de dominancia-diversidad del estudio corresponde a una distribución decreciente, donde se aprecia que hay un pequeño número de especies abundantes y una gran proporción de especies poco abundantes, lo que determina que las curvas sean como una “J” invertida (fig. 2).

Dominancia
El área evaluada presenta una cobertura de copa de 6 343.62 m2/ha, lo que representa una cobertura similar a lo reportado por Pequeño et al. (2012) con 6 844.87 m2/ha en un área de restauración pasiva postpecuaria en el matorral espinoso tamaulipeco, pero inferior al valor de 12 526.4 reportado por Alanís et al. (2011) al caracterizar de regeneración leñosa postincendio en una zona de transición entre el bosque de pino-encino y matorral. Cordia boissieri es la especie más dominante con 1 062.4 m²/ha, que es el 16.7 % de la cobertura del área de estudio. La especie que le continúa es Havardia pallens con 936.1 m2/ha correspondientes al 14.8% de la cobertura del área de estudio, y una tercera especie es Neopringlea integrifolia con 852.8m2/ha y una dominancia relativa de 13.4%. Por otro lado, la especie menos dominante es Prosopis laevigata, la cual presenta 1.53 m2/ha de cobertura, seguida por Condalia hookeri con 4.0 m2/ha y Citharexylon berlandieri con 9.4 m2/ha respectivamente (cuadro 2). Es importante mencionar que los mayores valores reportados para este parámetro en el MET en diferentes áreas con historial pecuario y agrícola los sustentan frecuentemente las especies Acacia farnesiana y Acacia amentacea (Alanís et al., 2008; Pequeño et al., 2012; Jiménez et al., 2012). Diferente a lo que reporta Mora et al., (2013) para un área sin disturbios, donde las especies dominantes son Acacia amentacea, Diospyros texana y Cordia boissieri, esta última se presenta en el área incendiada y con altos valores de importancia (cuadro 2).
Frecuencia
La frecuencia por especie está dominada por Havardia pallens, Cordia boissieri, Zanthoxylum fagara, Mimosa zygophilla, Celtis pallida, Croton suaveolens y Forestiera angustifolia con presencia en todos los sitios de muestreo, correspondientes a 6.5% de frecuencia relativa, las especies que continúan son Leucophyllum frutescens, Ebenopsis ebano, Acacia amentacea y Bernardia myricifolia con presencia en cuatro de los cinco sitios. Las especies menos frecuentes fueron Acacia farnesiana, Diospyros texana, Rhus virens, Citharexylon berlandieri, Condalia hookeri y Prosopis laevigatacon presencia en uno de los cinco sitios de muestreo (cuadro 2). Roque et al. (2013) reportó a Havardia pallens como la especie más frecuente al caracterizar un área del matorral espinoso tamaulipeco. Sin embargo, esta misma especie fue reportada con una frecuencia menor en un área postincendio de una zona de transición entre el bosque de pino-encino y matorral (Alanís et al., 2011).
Índice de valores de importancia (IVI)
El mayor peso ecológico obtenido mediante el índice de valor de importancia, lo tuvo la especie Havardia pallenscon 11.3% de IVI rel , las especies que le siguen en importancia son Leucophyllum frutescens con 10.2% y Cordia boissieri con 10.0%. Las especies con menor peso ecológico son Prosopis laevigata, Condalia hookeri y Citharexylon berlandieri con 0.5% (cuadro 2). Moreno (2013) reportó a Cordia boissieri, Havardia pallens y Leucophyllum frutescens como unas de las especies más importantes al caracterizar la vegetación de un área postincendio en el matorral espinoso tamaulipeco, coincidiendo con lo obtenido en este estudio, esto es de gran relevancia, ya que hace suponer que éstas son especies pioneras postincendio del matorral espinoso tamaulipeco. Además de que Havardia pallens se asocia a los incendios, ha sido reportada como una especie invasora en México (UNIBIO, 2009), por lo que se considera que su establecimiento después de un incendio se facilita (Pausas, 2004; Calvo et al., 2008; Weiguo et al., 2008).
Índices de diversidad de especies
La riqueza específica fue de 24 especies, superior a lo reportado por Mora et al. (2013) y por Pequeño et al. (2012), quienes evaluaron comunidades con perturbación diferente a los incendios, los cuales obtuvieron 21 y 12 especies respectivamente. Esto coincide con Beguet et al. (1987) quienes observaron que las áreas quemadas se van haciendo florísticamente más disímiles de las áreas sin quemar, a medida que transcurre el tiempo. Para el índice de Margalef el valor fue de D Mg = 3.16, lo cual indica una alta riqueza de especies si se compara con otras áreas regeneradas del matorral con perturbación antropogénica (Jiménez et al., 2012; Pequeño et al., 2012; Mora et al., 2013). En el índice de Shannon fue de H´= 2.52, el cual es un valor alto comparado con Alanís (2006) el cual obtuvo un índice de Shannon de 2.02 en una evaluación con diferente historial antropogénico. Mora et al. (2013) registró una diversidad H´= 1.95 evaluando el efecto de la ganadería en el MET. Pequeño et al. (2012), reporta valores de H´=1.27, en un análisis de la restauración, postpecuaria en el MET. Todo lo anterior concordando con Trabaud (1998), quien observo que la riqueza florística aumenta tras un incendio en el mediterráneo.
Conclusiones
De acuerdo con los resultados obtenidos en la presente investigación, se destacan las siguientes conclusiones: 1) la comunidad estudiada presenta una riqueza específica de 24 especies, lo cual es un valor alto si se compara con otras comunidades vegetales del MET regeneradas post-perturbación; 2) la especie Havardia pallens registró un 11.3% de IVI siendo la especie que presenta una mayor importancia ecológica en el área de estudio; por lo que se considera una especie resistente y que le benefician los incendios, 3) los índices de riqueza y diversidad (DMg = 3.16; H´= 2.52) registraron altos valores, esto demuestra que el área post-incendio cuenta con una buena regeneración y con alta diversidad de especies. La presente investigación aporta elementos cuantitativos de la vegetación arbórea y arbustiva de una comunidad incendiada, sugiriendo que este tipo de vegetación presenta alta resistencia a este disturbio.

