











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
México es considerado uno de los seis países que posee mayor diversidad biológica en el mundo junto con China, India, Colombia, Perú y Brasil (Mittermeier & Goettsch, 1997). Sin embargo, los ecosistemas han sufrido cambios en su estructura y composición, causando la pérdida de la diversidad biológica y propiciando un déficit en su funcionamiento (Hooper et al., 2005; Loydi & Distel, 2010). Estas alteraciones en los ecosistemas han sido provocadas por múltiples actividades humanas, debido a la necesidad de usos de suelos y desconocimiento de la utilidad potencial del sitio, la cual constituye una fuente de riqueza ecológica, genética, social, económica, educativa, cultural, recreativa y estética (Hooper et al., 2005; Santos & Tellería, 2006).
El presente estudio se enfoca al ecosistema denominado matorral espinoso tamaulipeco (MET) que cubre una superficie de 200 000 km² del noreste de México y sur de Texas (Estados Unidos de América). Esta región tiene una vegetación denominada semiárido-subtropical, localizado dentro de la Provincia de la Llanura Costera del Golfo Norte. El matorral está constituido por especies deciduas y siempre verdes o perennes, de contrastantes desarrollos fenológicos, con un amplio rango de patrones y dinámicas de crecimiento (González-Rodríguez & Cantú, 2001). Presenta una alta diversidad de especies arbóreas y arbustivas, cuyos rangos oscilan entre 15 000 a 21 000 individuos/ha (Alanís et al., 2008; González-Rodríguez et al., 2010; Jiménez-Pérez et al., 2013; Mora et al., 2013a ) Las especies de flora presentes en el MET se caracterizan por un amplio rango de grupos taxonómicos, siendo evidente las diferencias entre los patrones de crecimiento, diversidad en la longevidad foliar, dinámicas de crecimiento y desarrollo fenológico (Estrada et al., 2004).
Especies de árboles y arbustos constituyen la materia prima para las actividades productivas de uso tradicional silvoagropecuario, así como fuente de alimentación, combustible, herbolaria, construcción de muebles rústicos, productos artesanales y construcción de casas de habitación rural (Foroughbakhch et al., 2009). Sin embargo, el uso de la vegetación como alimento para animales domésticos (ganado bovino) en las áreas ha provocado modificaciones en la composición florística y la estructura de la formación vegetal (Turner, 1999).
La vegetación arbórea y arbustiva del MET en las últimas décadas ha sido expuesta a diferentes actividades silvoagropecuarias, causando la disminución de la capacidad de producción vegetal en la cubertura del suelo en áreas que se encontraban sujetas al aprovechamiento agrícola y pecuario (Pequeño-Ledezma et al., 2012). Estas actividades han provocado el abandono de tierras que actualmente se definen como la superficie de suelo conformada por espacios abiertos por la eliminación o disminución de la vegetación nativa, las cuales favorecen la germinación y rebrote de individuos de especies preexistentes o presentes en el banco de semillas del suelo (Sang et al., 2008).
A consecuencia, el paisaje se presenta como un mosaico de áreas en regeneración con un historial diferente de uso productivo (Alanís et al., 2008; Jiménez-Pérez et al., 2009). Las comunidades de vegetación secundaria resultantes presentan diferentes dinámicas sucesionales, dependiendo de la severidad de la perturbación, la dispersión de semillas de árboles padres en localidades cercanas y la capacidad de rebrote de especies preexistentes antes de la pertur-bación, y dan como resultado áreas con distinta composición vegetal (Boccanelli, 2011; Pincheira-Ulbrich & Hauenstein, 2008; Ramírez-Lozano et al., 2013). El objetivo de la presente investigación es evaluar la estructura y diversidad de la comunidad vegetal con historial de uso pecuario en el Matorral Espinoso Tamaulipeco (MET) del noreste de México.
Métodos
Área de estudio
El presente estudio se desarrolló en un área con historial pecuario del ejido Belisario Domínguez en el municipio de Mainero, Tamaulipas (Noreste de México: fig. 1), ubicadas a los 24°39’19.66” S, y 99°29’05.57” W, a una altitud promedio de 339 m s.n.m., en una fracción del matorral espinoso tamaulipeco de la Llanura Costera del Golfo Norte. El clima dominante en esta región es semicálido subhúmedo, en el cual registran precipitaciones en verano entre 700-1000 mm. El sistema de topoformas pertenece a lomeríos con llanuras, en donde los suelos predominantes son Leptosoles y las posibilidades de uso pecuario son el aprovechamiento de la vegetación natural únicamente por el ganado caprino y el establecimiento de praderas cultivadas con maquinaria agrícola, siendo el matorral la vegetación predominante (INEGI, 2009).

Análisis de la vegetación
Para cumplir con el objetivo propuesto, se seleccionó un área del matorral espinoso tamaulipeco que contara con un registro histórico de actividad pecuaria que implicara la remoción total de la vegetación (matarrasa), así como el establecimiento de pasto para el consumo del ganado y que por la pérdida de productividad de suelo tuviera un largo periodo de abandono.
En la década de los ochentas se realizó una modificación al ecosistema natural, realizando un desmonte con maquinaria agrícola, dicha actividad dejo desprovisto el suelo de vegetación arbórea y arbustiva. Bajo interés de mejorar las pasturas, se realizaron siembras de pastos exóticos, en este caso zacate buffel (Cenchrus ciliaris) siendo esta especie un reemplazo de las leguminosas nativas del área. Posteriormente en el año de 1992 después de ser utilizada para el pastoreo intensivo de ganado bovino se llevó a cabo el abandono de la práctica y la exclusión del área.
En verano de 2012, 20 años después de que se suspendiera la actividad pecuaria, se establecieron cuatro sitios de muestreo de 1 600 m2 (40 x 40 m) en una superficie de 18 hectáreas, para evaluar la estructura vegetal de las especies arbóreas y arbustivas mediante la determinación de la composición y diversidad florística (Canizales-Velázquez et al., 2009).
En los sitios de muestreo se realizó un censo de todas las especias leñosas, además se registró información dasométrica de altura total (h), diámetro basal (d 0.10 m) y diámetro de copa (d copa)en sentido norte-sur y este-oeste. La medición del diámetro basal se efectuó a 0.10 m sobre el suelo, siendo una medida estándar empleada para especies arbóreas y arbustivas del MET (Alanís et al., 2008; Jiménez et al., 2012), debido a las bifurcaciones que presentan los individuos a mayor altura, resultando más viable tanto práctico como económicamente para realizar una sola medición (Alanís et al., 2008; Jiménez et al., 2012; Marroquín-Castillo et al., 2016).
Para cumplir con los objetivos del estudio se determinaron parámetros ecológicos o variables estructurales relativas de abundancia (A i ), dominancia (D i ), frecuencia (F i ). Los resultados se utilizaron para obtener un valor ponderado a nivel de taxón denominado índice de valor de importancia (IVI) que adquiere valores porcentuales en una escala del 0 al 100 (Magurran, 2004; Mostacedo & Fredericksen, 2000; Mueller-Dombois & Ellenberg, 1974), así como el volumen (v) y área basal (g). Para estimar la diversidad alfa se utilizaron el índice de Shannon & Wiener (H´) e índice de riqueza de Margalef (D Mg ).
Análisis de la información
Las ecuaciones empleadas para realizar los cálculos se describen a continuación:
Para estimar la abundancia relativa se aplicó la siguiente ecuación:

Donde A i es la abundancia absoluta, AR i es la abundancia relativa de la especie i respecto a la abundancia total, N i es el número de individuos de la especie i, y S la superficie de muestreo (ha).
La dominancia relativa se evaluó mediante:

Donde D i es la dominancia absoluta, DR i es la dominancia relativa de la especie i respecto a la abundancia total, Ab i es el área de copa de la especie i, y S la superficie de muestreo (ha).
La frecuencia relativa se obtuvo con la siguiente fórmula:

Donde F i es la frecuencia absoluta, FR i es la frecuencia relativa de la especie i respecto a la abundancia total, P i es el número de sitios en el que está presente la especie i, y NS número total de sitios de muestreo (ha).
El índice de valor de importancia (IVI) se define como:

Para estimar la diversidad alfa se implementó el índice se Shannon & Wiener (Shannon, 1948) mediante la siguiente ecuación:

La riqueza de especies se estimó mediante el índice de Margalef (Clifford & Stephenson, 1975) con la ecuación:

Donde S es el número de especies presentes, In es el logaritmo natural y pi es la proporción de las especies, n i es el número de individuos de la especie i y N es el número total de individuos. Ambos índices cuentan con el mismo significado de variables.
Se estimó el volumen (V) de cada individuo mediante la fórmula:

Donde g es la superficie supuesta circular del diámetro basal (d 0.10 m), h la altura todal de individuo y CM el coeficiente mórfico, que para el caso del matorral espinoso tamaulipeco es 0.5 (Jiménez, 1990). Para describir la estructura de la comunidad se generó una gráfica de rango-abundancia (Brower et al., 1998) la cual describe la relación de la abundancia de las especies en función de un arreglo secuencial de especies, de la más a la menos importante (Martella et al., 2012).
Resultados
Se registraron 12 especies pertenecientes a 12 géneros y siete familias. La familia más representativa fue Fabaceae con seis especies, las restantes sólo se registró una especie. De acuerdo a la forma de vida cinco especies fueron arbustivas y siete arbóreas (cuadro 1).
Cuadro 1 Nombre científico y común, familia y forma de vida de las especies arbustivas y arbóreas registradas en el matorral espinoso tamaulipeco del área de estudio, ordenadas alfabéticamente.
| Nombre científico | Nombre común | Familia | Forma de vida |
|---|---|---|---|
| Caesalpinia mexicana A. Gray. | árbol del potro | Fabaceae | arbórea |
| Celtis pallida Torr. | granjeno | Cannabaceae | arbustiva |
| Cordia boissieri A. DC. | anacahuita | Boraginaceae | arbórea |
| Ebenopsis ebano (Berland.) Barneby & J.W. Grimes. | ébano | Fabaceae | arbórea |
| Forestiera angustifolia Torr. | panalero | Oleaceae | arbustiva |
| Havardia pallens (Benth.) Britton & Rose. | tenaza | Fabaceae | arbórea |
| Helietta parvifolia (A. Gray) Benth. | barreta | Rutaceae | arbustiva |
| Mimosa monancistra Benth. | charrasquillo | Fabaceae | arbustiva |
| Prosopis laevigata (Humb. & Bonpl. ex Willd.) M.C. Johnst. | mezquite | Fabaceae | arbórea |
| Randia obcordata S. Watson. | cruceto | Rubiaceae | arbustiva |
| Sideroxylon celastrinum (Kunth) T.D. Penn. | coma | Sapotaceae | arbórea |
| Vachellia farnesiana (L.) Wight &Arn. | huizache | Fabaceae | arbórea |
Parámetros ecológicos
La abundancia total fue de 1 156 N/ha. La especie que presentó mayor abundancia fue Vachellia farnesiana con 930 N/ha, que representa el 80.53% de la abundancia total. Las dos especies que le siguen fueron Prosopis laevigata y Celtis pallida, con 7.44% y 2.60% respectivamente. Las nueve especies restantes conforman el 9.43% faltante de dicho valor (cuadro 2).
Cuadro 2 Características de composición y estructura del matorral espinoso tamaulipeco postpecuario. Abundancia (N/ha), dominancia (m2/ha), frecuencia, índice de valor de importancia, área basal (m2/ha) y volumen (m3) de las especies arbustivas y arbóreas registradas en el matorral espinoso tamaulipeco del área de estudio, ordenadas de acuerdo al índice de valor de importancia.
| Núm. | Especie | Abundancia | Dominancia | Frecuencia | Valores de importancia | Área basal | Volumen | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| N/ha | % | m 2 /ha | % | N/Sitio | % | IVI | IVI rel | m 2 /ha | m 3 /ha | ||
| 1 | Vachellia farnesiana | 930 | 80.41 | 4 274.26 | 81.08 | 4 | 20 | 181.48 | 60.49 | 11.38 | 22.09 |
| 2 | Prosopis laevigata | 86 | 7.43 | 499.81 | 9.48 | 4 | 20 | 36.91 | 12.30 | 1.31 | 2.70 |
| 3 | Cordia boissieri | 20 | 1.76 | 145.87 | 2.77 | 2 | 10 | 14.52 | 4.84 | 0.38 | 0.58 |
| 4 | Havardia pallens | 28 | 2.43 | 58.56 | 1.11 | 2 | 10 | 13.54 | 4.51 | 0.17 | 0.43 |
| 5 | Celtis pallida | 30 | 2.57 | 109.50 | 2.08 | 1 | 5 | 9.64 | 3.21 | 0.08 | 0.15 |
| 6 | Caesalpinia mexicana | 23 | 2.03 | 120.05 | 2.28 | 1 | 5 | 9.30 | 3.10 | 0.30 | 0.55 |
| 7 | Forestiera angustifolia | 17 | 1.49 | 13.73 | 0.26 | 1 | 5 | 6.75 | 2.25 | 0.04 | 0.05 |
| 8 | Sideroxylon celastrinum | 8 | 0.68 | 17.12 | 0.32 | 1 | 5 | 6.00 | 2.00 | 0.02 | 0.04 |
| 9 | Ebenopsis ebano | 5 | 0.41 | 20.08 | 0.38 | 1 | 5 | 5.79 | 1.93 | 0.07 | 0.14 |
| 10 | Randia obcordata | 5 | 0.41 | 4.14 | 0.08 | 1 | 5 | 5.48 | 1.83 | 0.00 | 0.00 |
| 11 | Helietta parvifolia | 3 | 0.27 | 3.93 | 0.07 | 1 | 5 | 5.34 | 1.78 | 0.01 | 0.01 |
| 12 | Mimosa monancistra | 2 | 0.14 | 4.91 | 0.09 | 1 | 5 | 5.23 | 1.74 | 0.00 | 0.00 |
| 1 156 | 100 | 5 271.96 | 100 | 4 | 100 | 300 | 100 | 13.76 | 26.74 | ||
La dominancia de una especie se estimó mediante la cobertura de copa. En general, la cobertura de copa del total de las especies evaluadas presentó 5 271.96 m2/ha, es decir un 52.72% del área está ocupada por el dosel de los árboles y arbustos. Sólo el 47.28% de la superficie se encuentra desprovista de vegetación. La especie altamente dominante es Vachellia farnesiana, presentando el 81.08% de la cobertura total de la vegetación, seguida por Prosopis laevigata con el 9.48%. Las restantes 10 especies sólo representan el 9.44%.
Se registró la presencia de dos especies en todos los sitios de muestreo, Vachellia farnesiana y Prosopis laevigata con una representatividad de 40% de frecuencia relativa, le siguen Cordia boissieri y Havardia pallens con presencia en dos de los cuatro sitios de muestreo. Las ocho especies restantes sólo se presentaron en un sitio de muestreo, teniendo en conjunto la misma representatividad que Vachellia farnesiana y Prosopis laevigata.
La especie con mayor peso ecológico, según el índice de valor de importancia es Vachellia farnesiana con 60.49%, seguido por Prosopis laevigata y Cordia boissieri con el 12.30% y 4.84%, respectivamente. Estas tres especies representan el 77.63%. Las especies con menor peso ecológico son Hellieta parvifolia y Mimosa monancistra con 1.78% y 1.74% respectivamente.
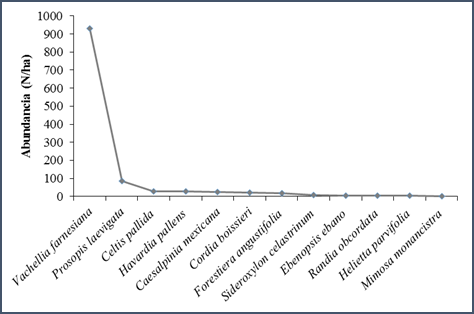
Según el rango-abundancia, las especies en estudio presentan una distribución tipo geométrico, de acuerdo a este modelo, la comunidad está compuesta por pocas especies muy abundantes y las restantes prácticamente escasas. El modelo supone que las especies difieren marcadamente en su dominancia competitiva, de manera que la especie más dominante ocupa una fracción grande del nicho y las demás especies deben distribuirse lo que queda. Este patrón se observa básicamente en estadios tempranos de una sucesión o en ambientes desfavorables (Martella et al., 2012). Para este estudio la única especie que presentó una abundancia alta fue Vachellia farnesiana con 930 N/ha (fig. 2).
La figura 3 indica la densidad de individuos por hectárea de acuerdo con las clases diamétricas registradas en el estudio. Principalmente se observa una línea de tendencia exponencial negativa en la densidad de individuos conforme aumentan sus diámetros, siendo la clase I (0-6 cm de diámetro) la que presentó mayor número de individuos (424 N/ha).

La figura 4 indica la densidad de individuos por hectárea de la especie Vachellia farnesiana, la cual es dominante en el área de estudio. Se aprecia que el 55% (330 N/ha) se encuentra en un rango diamétrico de 0 a 6 cm.

Discusión
De acuerdo a nuestros análisis, la familia Fabaceae es la más representativa en el área de estudio. Este resultado concuerda con los obtenidos en otras investigaciones, por ejemplo de Alanís et al. (2013), Molina-Guerra et al. (2013) , Mora-Donjuán et al. (2014) , Jiménez et al. (2012) y Mora et al. (2013b). Estos estudios sugieren que las comunidades vegetales del MET tanto maduras (Mora et al., 2013a) como regeneradas después alteraciones antropogénicas presentan a las Fabaceaes como las más representativas.
Abundancia. En la presente investigación se registraron 12 especies, número similar a lo registrado por Pequeño-Ledezma et al. (2012) y Yerena et al. (2014) quienes también evaluaron comunidades vegetales regeneradas del matorral espinoso tamaulipeco. Las comunidades vegetales maduras como la evaluada por Mora et al. (2013a) presentan 21 especies, casi el doble de lo registrado en este estudio. Esto podría deberse a que la comunidad vegetal evaluada en el presente estudio aún está en estadíos iniciales o intermedios de sucesión ecológica y algunas especies de estadíos tardíos aún no se presentan como el caso de Yucca filiffera (Alanís et al., 2013 ).
Asimismo, se observó que Vachellia farnesiana y Prosopis laevigata fueron las especies que presentaron mayor densidad poblacional. Esta información concuerda con los resultados obtenidos por Alanís et al. (2013) y de Jiménez-Pérez et al. (2013), quienes también evaluaron comunidades vegetales regeneradas del MET. La alta presencia de estas especies se podría considerar como indicadoras de disturbio, ya que son altamente abundantes en áreas regeneradas y poseen una abundancia baja en comunidades maduras.
Con respecto a las especies con mayor número de individuos por hectárea, nuestros resultados concuerdan con los de Yerena et al. (2014), quienes evaluaron un área del matorral espinoso tamaulipeco. En su estudio evaluaron una comunidad vegetal con un registro histórico de actividad pecuaria con un periodo de 20 años de abandono, registrando a las especies Vachellia farnesiana, Prosopis laevigata y Celtis pallida con los valores más altos con 930, 86 y 30 respectivamente. Por otro lado, también son similares a los resultados obtenidos por Pequeño-Ledezma et al. (2012) en un área regenerada postpecuaria del matorral espinoso tamaulipeco situado al noreste de México, donde registraron a Vachellia farnesiana, Prosopis laevigata en primero y segundo lugar respectivamente y Celtis pallida en cuarto lugar.
Según Yerena et al. (2014), Vachellia farnesiana presenta un patrón de disminución de individuos por hectárea según la edad de abandono, limitando el restablecimiento de otras especies, comportándose como dominante en los primeros años de la sucesión ecológica. Nuestros resultados también concuerdan con los de Estrada et al. (2004) y de Jiménez et al. (2012), donde mencionan que las aéreas abandonadas que han sufrido algún tipo de disturbio por actividad antropogénica tienden a reflejar una baja disponibilidad de nitrógeno, resultando el establecimiento de esta especie leguminosa. La dominancia de esta especie en la restauración pasiva postpecuaria se debe a su plasticidad fenotípica, dominando en los primeros estadíos sucesionales, y gradualmente de manera natural con el paso de los años permite el establecimiento de otras especies.
Dominancia.Mora et al. (2013a) indican que la cobertura de copa del matorral espinoso tamaulipeco sin registro de actividades productivas (comunidad madura) presenta valores de 13 973 m2/ha, lo cual muestra que el valor obtenido en esta investigación (5 271 m2/ha) es bajo y el área no se encuentra cubierta completamente por el dosel. No obstante, la cantidad registrada en este estudio se aproxima a los obtenidos por Pequeño-Ledezma et al. (2012) con un valor de 6 844.87 m2/ha para un área de restauración pasiva postpecuaria en un área con 30 años de abandono. Las especies dominantes en el área de estudio son Vachellia farnesiana y Prosopis laevigata presentando el 81.07% y el 9.48% de la cobertura total de la vegetación, siendo las mismas especies registradas por Pequeño-Ledezma et al. (2012) pero con valores menores 61.77% y 12.80% respectivamente. Esta similitud en las especies y porcentajes podría deberse a que ambos estudios evaluaron comunidades vegetales con historial pecuario y se encuentran en estadíos sucesionales intermedios o iniciales.
Frecuencia. En cuanto a la frecuencia, nuestros resultados concuerdan con otros estudios, principalmente por aquellos observados por Pequeño-Ledezma et al. (2012) , que evaluaron un área de restauración pasiva post-pecuaria del MET y encontraron a las especies Vachellia farnesiana y Prosopis laevigata en todos los sitios de muestreo.
Índice de valor de importancia (IVI). La especie con mayor peso ecológico observada en este estudio fue Vachellia farnesiana, este resultado es similar al registrado por Pequeño-Ledezma et al. (2012) para un área de matorral espinoso tamaulipeco con impacto de ganadería con 30 años de abandono. A su vez, esta especie es la que presenta una mayor presencia en un área de ganadería extensiva según lo registrado por Alanís et al. (2008).
Índice de Shannon-Wiener (H’). El índice de Shannon obtenido en este estudio fue de 0.86, este valor es bajo comparado con comunidades vegetales maduras del MET, las cuales presentan valores superiores a los 1.90 (Mora et al., 2013a). También este valor es menor si se compara con resultados obtenidos en otros matorrales del noreste de México (Alanís-Rodríguez et al., 2015; Canizales-Velázquez et al., 2009; Mora-Donjuán et al., 2014).
Conclusiones
Con los resultados de la presente investigación se concluye que 1) después de abandonar la ganadería extensiva con zacate buffel (Cenchrus ciliaris) en el matorral espinoso tamaulipeco de Linares N.L se generan condiciones favorables para que restablezca las especies leñosas nativas, específicamente las de la familia Fabaceae ya que se ha demostrado que esta familia presentan tolerancia a suelos con escaso nitrógeno, y presentan características fisiológicas que le permiten colonizar, esta capacidad de regeneración se ve reflejada en los altos valores de densidad, cobertura, riqueza y diversidad registrados en este estudio. 2) La especie de mayor importancia por su contribución estructural a la comunidad fue Vachellia farnesiana. 3) Dicha comunidad de acuerdo a la gráfica de rango-abundancia presenta una distribución de modelo geométrico, ya que está dominada por la alta presencia de la especie V. farnesiana. 4) Existe alta presencia de individuos en las clases diamétricas menores, lo cual muestra un estado de regeneración activo. El estudio proporcionó información cuantitativa sobre la restauración pasiva postpecuario del MET, generando información fitosociológica de la comunidad vegetal.

