











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Limnobium laevigatum (Humb. & Bonpl. ex Willd.) Heine. es una planta acuática libremente flotante que pertenece a la familia Hydrocharitaceae. Se caracteriza por tener rápido crecimiento, invadiendo ecosistemas acuáticos y transformándose en una especie invasora en países de Norte y Sud América (San Martín y Boetscher, 2003; USDA, ARS, National Genetic Resources Program, 2014). En el Perú, esta especie se distribuye entre los 0 y 500 metros sobre el nivel del mar, habiéndose registrado para ambientes lacustres y ribereños en los departamentos de Lima, Ucayali y Loreto (Brako y Zarucchi, 1993; Ramirez y Cano, 2010).
A pesar de ser altamente invasiva, la función de L. laevigatum en los humedales es muy importante, ya que esta especie forma parte de la dieta de algunas aves sudamericanas como Cygnus melancoryphus Molina, 1782 y Gallinula chloropus Linnaeus, 1758 (Beltzer et al., 1991; Corti y Schlatter, 2002), así como de algunos invertebrados como Paulinia acuminata (Carbonell et al., 2006). Corti y Schatteler (2002) hacen un estudio químico proximal de L. laevigatum, mostrando que, en condiciones naturales, puede llegar a tener hasta un 16% de proteínas (% en base seca). Posteriormente Aponte et al. (2013) indican que, en condiciones de laboratorio, las plantas tienen un valor proteico más alto (entre 26 y 30%) y bajo contenido en fibras (7%) en la materia seca; e indican que esta especie es un recurso potencial para su uso como forraje.
Su tasa de crecimiento relativo en condiciones de laboratorio (TCR = 0.124) es más alta que su similar reportada para otras macrófitas acuáticas como Egeria densa Planch. y Eichhornia crassipes (Mart.) Solms. (Reddy y DeBusk, 1984; Henry-Silva et al., 2002; Pistori et al., 2004; Aponte y Pacherres, 2013). Plantear modelos de crecimiento adecuados para esta especie, permitiría estimar el potencial de la planta para su uso en sistemas forrajeros a gran escala.
El objetivo del presente estudio fue estimar la productividad de Limnobium laevigatum (producción de biomasa, de carbono y de proteínas) en laboratorio, estableciendo un modelo matemático que describa este crecimiento bajo condiciones controladas.
Materiales y métodos
Propagación de las plantas y estimación de la capacidad de carga (k)
A fin de tener un modelo de crecimiento poblacional con recursos limitados, se optó por realizar una estimación de K. Para ello se utilizaron 30 rametos de L. laevigatum pertenecientes a un cultivo constante de esta especie en las instalaciones de la Universidad Científica del Sur (Lima, Perú). Cada rameto se colocó en un envase de vidrio de 600 ml (50 cm2 de área superficial), con 300 ml de solución nutritiva, haciendo un experimento con 30 repeticiones simultaneas. La solución nutritiva que se utilizó fue aquella que permite obtener las tasas de crecimiento más altas y alta concentración de proteínas en los tejidos de la planta (25X en Aponte y Pacherres, 2013; 2X en Aponte et al., 2013). Cada envase de vidrio estuvo protegido por un plástico negro que cubría la exposición de la solución a la luz, evitando el crecimiento de algas. Cada siete días los envases de vidrio fueron limpiados (se retiró la materia muerta) y la solución de cada envase fue cambiada por una nueva a la misma concentración a fin de mantener las mismas condiciones de nutrientes a lo largo del experimento (Aponte y Pacherres, 2013).
Las condiciones de iluminación, fotoperiodo, temperatura y humedad durante el experimento se muestran en la tabla 1. Se registró la biomasa cada siete días utilizando una balanza eléctrica (0.01 g de precisión). Se midió la tasa de crecimiento relativo semanal (TCRs) siguiendo la siguiente fórmula:
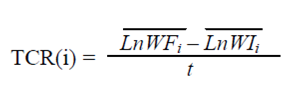 (1)
(1)
Donde WF y WI son el peso de cada planta al inicio y al final de cada semana; el tiempo (t) fue de siete días. Se consideró que las plantas habían alcanzado K cuando la TCRs promedio se hizo cero. Posterior a ello se continuó con el experimento durante tres semanas más a fin de analizar el comportamiento de la población.
Tabla 1. Condiciones de iluminación, fotoperiodo, temperatura y humedad durante el primer experimento de capacidad de carga. Se muestra el promedio ± DE (máximo-mínimo).
| Condición | Promedio ± DE (máximo-mínimo) |
|---|---|
| Luz (µmol/m2/seg) | 142.6+/-141.61 (0.34-764) |
| T Ambiental (°C) | 24.61+/-1.25 (22.4-27.3) |
| T H2O (°C) | 26.38+/-2.49 (22.7-32.1) |
Modelamiento del crecimiento poblacional
Terminado el experimento para hallar K se analizó la totalidad de los datos en función del tiempo y se realizaron regresiones con la finalidad de conocer el modelo que mejor se adapta a esos datos. Las regresiones probadas fueron las logísticas y de la familia del crecimiento logístico (Logístico, Sinusoidal y Grompetz). Por cada modelo se halló el Criterio de Información de Akaike (AIC), el coeficiente de determinación (R2), el coeficiente de correlación y el error estándar, de tal manera que se pudo elegir el mejor modelo (aquel que muestra mayor correlación, mayor coeficiente de determinación, menor AIC y que menor error). El uso de estos modelos y la evaluación de estos parámetros son recomendados para la selección de modelos en organismos vegetales, dado que su crecimiento en condiciones de recursos limitados tiene como particularidad la disminución de la TCR conforme va creciendo la población y/o tamaño de la planta (Paine et al., 2012). Las regresiones no lineales y obtención de parámetros de selección de modelos fueron obtenidas utilizando los softwares PAST 2.17c (Hammer et al., 2001) y Curve Expert profesional 2.2 (Hyams, 2010).
Una vez seleccionado el mejor modelo, se aplicó la regresión correspondiente a cada una de las 30 repeticiones del experimento. De esta manera se obtuvieron 30 valores de los parámetros del modelo, pudiendo así, tener datos estadísticos descriptivos de los mismos.
El momento exacto en que la TCRs se volvió cero, se calculó mediante una regresión logarítmica utilizando Microsoft Excel (©Microsoft bajo licencia).
Porcentaje de humedad, proteínas y carbono en el tejido vegetal
El porcentaje de carbono se halló mediante el análisis del tejido siguiendo la técnica descrita por Walkley y Black (1934) por triplicado. El porcentaje de humedad (95.2%) y proteínas (30.76%) fueron obtenidos de estudios previos donde las plantas crecieron bajo condiciones similares (Aponte et al., 2013).
Estimación de la productividad
Habiendo seleccionado el mejor modelo, se utilizó el valor de K del modelo (Kmod) para calcular el potencial de biomasa producible por área en condiciones de laboratorio. Este valor se multiplicó por los factores de conversión en biomasa seca, de proteínas y en carbono, obtenidos previamente (Aponte et al., 2013 y explicados en la sección anterior). Las fórmulas para el cálculo de la productividad en unidades de carbono (PC) y proteínas (PP) se indican en las fórmulas (2) y (3):
 (2)
(2)
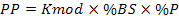 (3)
(3)
Donde Kmod es la capacidad de carga obtenida con el modelo, %BS es el porcentaje de materia seca, %C es el porcentaje de carbono en la materia seca y %P es el porcentaje de proteínas en la materia seca. Se hallaron los valores máximos y mínimos de la productividad utilizando los valores máximos y mínimos de Kmod.
Resultados
Crecimiento en laboratorio y análisis de k
La figura 1 muestra el crecimiento promedio de las plantas en el tiempo. La figura 2 muestra la evolución de la TCRs a lo largo del experimento; asimismo muestra los resultados de la regresión. Según este último resultado, TCRs se hizo cero a los 85 días del experimento (R2=0.87), cuando la biomasa era de 8.9 gramos. Este valor puede dividirse entre el área superficial de los beakers para tener el dato de la densidad promedio por cm2 a la cual la curva deja de crecer (K) la cual es equivalente a 0.178 g/cm2.

Fig. 1. Crecimiento de las plantas en el experimento para hallar K. Se muestra el peso promedio durante el experimento.

Modelamiento del crecimiento poblacional
Se observó que el modelo que mejor representa el crecimiento de la especie es el modelo logístico ya que los valores de AIC fueron los más bajos (-25.79), el coeficiente de correlación fue cercano a 1 (0.99), tuvo el menor error estándar (0.37) y el coeficiente de determinación también fue cercano a 1 (0.99) (tabla 2).
Tabla 2. Criterio de Información de Akaike (AIC), coeficiente de determinación (R2), coeficiente de correlación (Coef. Correl) y error estándar (ES) para los cuatro modelos evaluados.
| AIC | R2 | COEF. CORREL | ES | |
|---|---|---|---|---|
| Sinusoidal | 40.83 | 0.01 | 0.11 | 3.76 |
| Logístico | -25.79 | 0.99 | 0.99 | 0.37 |
| Grompetz | -17.12 | 0.98 | 0.99 | 0.51 |
Los estadísticos descriptivos para los parámetros obtenidos de la regresión logística obtenida se indican en la tabla 3. Utilizando el promedio de los valores obtenidos, el modelo del crecimiento poblacional para la especie en estudio es:
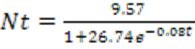 (4)
(4)Este modelo considera que la población en t = 0 tiene una densidad de 6x10-3g/cm2. El parámetro a (9.57) que dividido entre el área de beaker corresponde al K del modelo (Kmod) que en este caso es 0.1914 g/cm2.
Tabla 3. Estadísticos descriptivos para los parámetros obtenidos de la regresión logística.
| a | b | c | |
|---|---|---|---|
| N | 30.00 | 30.00 | 30.00 |
| Min | 7.56 | 3.56 | 0.05 |
| Max | 11.88 | 82.83 | 0.10 |
| Pro | 9.57 | 26.74 | 0.08 |
| ES | 0.20 | 3.27 | 0.00 |
| Var | 1.18 | 320.67 | 0.00 |
| DS | 1.09 | 17.91 | 0.01 |
| CV | 11.34 | 66.96 | 15.32 |
N = número de datos; Mín = mínimo; Mx = máximo; Pro = promedio; ES = error estándar; Var = varianza; DS = desviación estándar; CV = coeficiente de variación. Los valores a, b, y c son parámetros del modelo logístico, donde a es la capacidad de carga (K) y b y c son constantes del denominador (para más detalle ver ecuación 4).
Conversión a unidades de productividad
Utilizando Kmod en las ecuaciones 2 y 3, se calculó la productividad de Limnobium laevigatum. Esta especie, puede producir 2.83 x 10-3 g/cm2 de proteínas. En el caso de carbono, el resultado del análisis de contenido de carbono fue de 43%, por lo que su productividad en unidades de carbono equivale a 3.96 x 10-3 g/cm2 de carbono.
Discusión
A diferencia de las ecuaciones exponenciales, los modelos logísticos son más realistas y se adaptan mejor a la forma de crecimiento de las plantas en general, cada vez que se considere los recursos como un factor limitante (Birch, 1999; Paine et al., 2012). Los parámetros de selección de modelo indican que la ecuación utilizada se adapta a los resultados originales dado que tiene un error bajo, coeficiente de determinación alto y un coeficiente de correlación alto. Estos resultados permiten tener confianza en el modelo y utilizarlo en la predicción de esta especie en condiciones de laboratorio. Los datos obtenidos en el presente estudio corresponden a los primeros que describen el crecimiento en laboratorio de esta especie considerando el límite de recursos y podrán ser utilizados como base para optimizar el uso de la especie en diversos propósitos, tales como la biorremediación (Murillo Castillo et al., 2012) o forraje (Aponte et al., 2013); aunque para ello es necesario realizar un mayor trabajo de campo y experimental, a fin de complementar el poco conocimiento que se tiene sobre esta especie para estas aplicaciones.
Uso del modelo para estimaciones a gran escala
El modelo propuesto describe un crecimiento bajo condiciones limitadas de nutrientes y espacio. Los datos obtenidos ofrecen un indicio de la producción de esta especie a gran escala bajo las condiciones de crecimiento del presente estudio. Por ejemplo, según los resultados, la especie crece durante 80.5 días (momento en el que la TCRs se hace 0); esto significa que, optimizando el proceso productivo a gran escala, podría repetirse esta misma operación al menos cuatro veces al año (4.53 veces exactamente). Convirtiendo los resultados obtenidos en el presente estudio en unidades mayores (en Toneladas por hectárea y por año), se obtendría la producción de 1.28 (1.01-1.59) T/ha/año de proteínas y 1.79 (1.41-2.23) T/ha/año de carbono.
La producción de proteínas de Limnobium laevigatum obtenida con los cálculos del presente estudio es mayor que la producción de otras especies acuáticas como Justicia americana (L.) Vahl, Sagittaria latifolia Wild., Nymphaea odorata Aiton, Jussiaea decurrens (Walter) DC., Polygonum sp., Nymphoides aquatica (J.F. Gmel.) Kuntze y Nelumbo lutea Wild., la cual se encuentra entre 0.1 T/ha y 0.5 T/ha de proteínas (Boyd, 1968). Las plantas acuáticas, como Limnobium laevigatum, pueden ser utilizadas como alimentos para animales de granja y peces dado a que son fuentes proteicas de alto valor nutricional (18 a 32% en base seca), aunque algunas tienen como deficiencia el bajo porcentaje de materia seca (5-6 %, Hang et al., 1997, Escamilla 1998), tal y como se aprecia en la especie estudiada. Por otro lado, Limbobium laevigatum posee valores de proteína bruta óptimos (entre 26 y 30% en base seca, Aponte et al., 2013), cosa que también ha sido encontrada en otras plantas acuáticas utilizadas en la alimentación de organismos acuáticos, como Lemna gibba L. (proteína bruta entre 20 y 30% BS, Dewanji, 1993; Gutiérrez et al., 2001; Ponce & Fitz, 2004) y Lemna perpusilla Torr. (entre 24 y 26%, González 2010). El presente trabajo sienta las bases para plantear la propagación de esta especie a mayor escala, teniendo en consideración su potencial de producción proteica.
Dado que la forma de calcular la capacidad captadora de carbono incluye medir cuánto carbono se almacena en la biomasa vegetal (IPCC, 2006), los valores de productividad de carbono para Limnobium laevigatum obtenidas en el presente estudio representan también la cantidad de carbono que puede almacenar la especie en un espacio determinado. Si se compara con otros valores de captación de carbono conocidos, los valores estimados para la especie en estudio se asemejan a los niveles de captación de carbono en sistemas silvícolas y agroforestales diseñados con fines de captura de carbono (0.93-7.23 T/ha/año de carbono; Soto-Pinto et al., 2001), y son mayores a los de la productividad lograda por las plantas en humedales de zonas templadas frías (0.08-0.6 T/ha/año; Roulet, 2000). Por otro lado, su capacidad de captación de carbono es menor que la obtenida en bosques semiáridos de pinos (1.6 T/ha/año; Dore et al., 2008) y en pastizales de zonas templadas (1.9-2.6 T/ha/año; Jaksic et al., 2006). Se conoce la productividad de algunas especies acuáticas en condiciones silvestres como Eichhornia azurea, Nymphea rudgeana (Coutinho, 1989; Camargo y Florentino, 2000), Pontederia cordata (Penha, 1994) y Nymphoides indica (Menezes, 1984), los cuales se encuentran entre 3.5 T/ha y 17.5 T/ha de carbono; lo que nos muestra, que, en las condiciones realizadas en el presente estudio, la especie tiene una menor producción (aunque en los trabajos mencionados no se incluye el tiempo de producción de dicha biomasa). Estudios posteriores deberán dirigirse para conocer si el crecimiento de Limnobium laevigatum podría mejorar en condiciones naturales u optimizando las condiciones de laboratorio.
Conclusiones
El modelo que mejor representa el crecimiento de Limnobium laevigatum bajo condiciones limitadas de nutrientes y espacio es el modelo Logístico. Los datos obtenidos pueden dar un indicio de la producción de esta especie a gran escala. La productividad de Limnobium laevigatum bajo condiciones controladas asciende a 0.19 gr/cm2 de biomasa fresca, que equivalen a 2.83 x 10-3 gr/cm2 de proteínas y 3.96 x 10-3 gr/cm2 de carbono.
Los resultados alcanzados describen el crecimiento en laboratorio de esta especie considerando el límite de recursos y podrán ser utilizados como base para optimizar el uso de Limnobium laevigatum, por ejemplo, en biorremediacíon o como forraje.

