











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
México es el octavo productor y el tercer exportador de miel en el mundo (FAOSTAT, 2014). Durante 2013 exportó 33 mil 476 toneladas de miel, que alcanzaron un valor de 112 millones de dólares (SAGARPA, 2014). Sin embargo, mucha de la producción de miel se concentra en las regiones del sureste, sur y occidente del país. En el estado de Baja California la actividad es más bien marginal; se cuenta con alrededor de 7 mil 600 colmenas, con rendimientos en miel por colmena inferiores a los 12 kilogramos por año (SIAP, 2014), lo cual es bajo en comparación con otros estados de México. Esta actividad es más frecuente en el área agrícola del municipio de Mexicali, donde se concentra el 81% de las colmenas y el 91% de la miel que se cosecha en la entidad (OEIDRUS, 2010).
La rentabilidad de la producción apícola y las poblaciones saludables de abejas dependen de paisajes ricos en flores productoras de néctar y polen abundante (Decourtye et al., 2010). Es claro que el conocimiento de la flora apícola, la distribución geográfica y épocas de floración y su contribución a la producción, constituyen la información básica para determinar el potencial productivo de la apicultura en una región. En el estado de Baja California, se carece de este tipo de información, no se conocen los principales recursos nectaríferos que contribuyen a la producción de miel, lo cual sería importante para establecer alternativas de repoblación artificial y conservación con plantas nativas melíferas, con la finalidad de incrementar la productividad apícola del estado.
La metodología más económica y común para determinar el origen del néctar y la contribución de los recursos néctar-poliníferos a la producción de miel consiste en el estudio melisopalinológico de la misma (Jones y Bryant, 2014; Sahinler et al., 2009). El análisis de los tipos polínicos presentes en la miel, permite conocer su origen botánico para darle valor agregado, identificar el hábitat y estado de conservación de la vegetación del entorno en que fue producida, determinar la proporción de uso de especies silvestres y cultivadas, y contribuir a la toma de decisiones que mejoren la producción apícola (Ramírez-Arriaga et al., 2011; Forcone y Ruppel, 2012; González-Porto et al., 2013; Jones y Bryant, 2014; Kamble et al., 2015).
En México se han realizado diversos estudios (Villanueva-Gutiérrez et al., 2009; Ramírez-Arriaga et al., 2011; Castellanos-Potenciano et al., 2012; Zavala-Olalde et al., 2013 y Villanueva-Guriérrez et al., 2014) relativos a la melisopalinología y la identificación de recursos nectaríferos y poliníferos, para la flora néctar-polinífera del sur y sureste mexicanos y no existen antecedentes para el estado de Baja California.
Por lo anterior, este análisis se planteó para determinar los recursos nectaríferos importantes en el Valle de Mexicali, a partir de la identificación y cuantificación de los tipos polínicos presentes en las mieles, y determinar si se producen mieles que puedan ser denominadas como monoflorales.
Material y métodos
El trabajo se llevó a cabo durante los ciclos 2010 y 2011, en el norte del estado de Baja California, en la región conocida como Valle de Mexicali, perteneciente al municipio del mismo nombre (fig. 1). El Valle de Mexicali se localiza entre los 31° 40’ a 32° 40’ N, 114° 45’ a 115° 40’ O y una altitud sobre el nivel del mar entre 5 m y 28 m. Limita al norte con California, Estados Unidos, al sur y sureste con el Golfo de California y la sierra El Mayor, al oeste con la Sierra Cucapá y al este con Arizona, Estados Unidos y con San Luis Río Colorado, Sonora (Camargo-Bravo y García-Cueto, 2012).
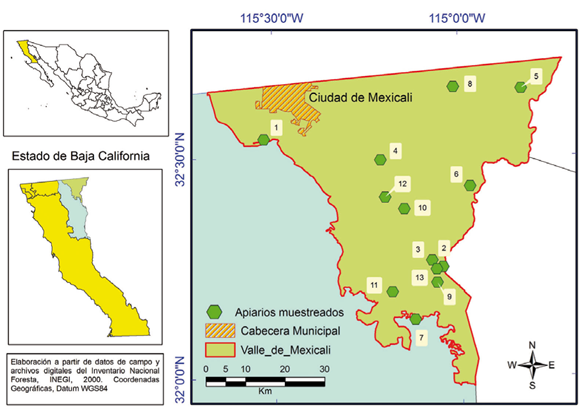
Fig. 1 Área de estudio y de apiarios muestreados. 1 Coronita, 2 Francisco Murguía, 3 Col. Hidalgo, 4 Col. Pólvora, 5 Cuervos, 6 Ejido Chiapas, 7 Col. Lerma, 8 Col. Borquez, 9 Ejido Plan de Ayala, 10 Ejido Guerrero, 11 Col. Carranza, 13 Ejido Nuevo León, 13 Francisco Murguía.
Su clima se caracteriza como extremadamente árido, con precipitaciones que raramente exceden los 100 mm al año (Brun et al., 2010). García (1988) lo clasifica como cálido seco, muy extremoso con gran oscilación anual de temperatura y precipitación escasa de régimen intermedio entre verano e invierno.
La zona de estudio tiene la peculiaridad de ser un área agrícola de riego, fuertemente intervenida por la acción humana y cuya cubierta vegetal original ha sido removida. La vegetación original corresponde al matorral desértico sonorense (Delgadillo, 1998), en la subdivisión del Valle Bajo del Río Colorado, donde dominan los matorrales de Larrea tridentata y Ambrosia dumosa (gobernadora). Además, existen comunidades riparias donde Typha dominguensis (tule) es la planta dominante, acompañada del pasto Distichlis palmeri; la vegetación dominante en áreas estables sin inundación frecuente son árboles con comportamiento ripario, como Salix goodingii (sauce), Populus fremontii (álamo) y Prosopis pubescens (tornillo), además de arbustos como Pluchea sericea (cachanilla), y árboles como Prosopis glandulosa var. torreyana (mezquite); también se han establecido otros como Pithecellobium dulce (guamúchil). No obstante, Tamarix spp. (pino salado) ha remplazado mucha de la vegetación riparia original (Delgadillo, 1998), y las áreas agrícolas donde se siembra algodón, alfalfa, gramíneas para forraje y grano representan el principal uso de la tierra.
Se recolectaron en campo 52 muestras de miel, de 13 apiarios localizados en el Valle de Mexicali (fig. 1), en cuatro fechas de recolecta, en los meses de junio y agosto de 2010 y 2011. Cada muestra consistió de cinco piezas de panal (5 x 5 cm) con miel en proceso de ser operculada, proveniente de un cuadro del alza melaria de cada una de cinco colmenas seleccionadas en forma aleatoria en cada apiario.
Para identificar los tipos polínicos presentes en la miel se recolectaron especímenes vegetales que fueron identificados y depositados en el herbario BCMEX. De cada planta recolectada se conservaron, por separado, flores en un sobre de papel, para la posterior extracción, acetólisis y montaje de su polen, con lo cual se conformó una colección palinológica de referencia que contiene 150 especies de plantas visitadas por las abejas en las inmediaciones de los apiarios muestreados.
El polen de las plantas para conformar la colección palinológica de referencia se obtuvo de macerar las anteras de cada muestra de flores en 10 ml de agua destilada y pasar el sedimento por una malla tipo organza, este residuo fue procesado por acetólisis y montado en dos portaobjetos usando una pequeña cantidad de gelatina glicerinada (Costa et al., 2013).
Las cinco piezas de panal procedentes de cada apiario fueron macerados y luego se separó la cera de la miel mediante filtrado a través de una malla de tela de 500 μm. Las muestras de miel filtrada fueron entonces almacenadas en contenedores de vidrio a temperatura ambiente. El polen se extrajo de una muestra de 15 g de miel, se colocó en un tubo de plástico de 50 ml, se agregaron 25 ml de agua destilada y se disolvieron a baño maría a 60°C por 10 min, posteriormente fueron sometidas al proceso de acetólisis (Erdtman, 1943).
Para el análisis cuantitativo se utilizó el método de Loveaux et al. (1978), actualizado por Von der Ohe et al. (2004). Consistió en diluir 10 ml de miel, pasarlo a través de un filtro de membrana con poros de 3 μm de diámetro (MF-Millipore SSWP02500), después se secó el filtro a una temperatura de 40°C, se transparentó con aceite de inmersión y se contabilizaron los elementos retenidos bajo un microscopio. El número de partículas por unidad de peso se calculó mediante la fórmula:
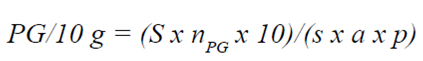
donde:
PG |
número absoluto de granos de polen en 10 g de miel. |
S |
área del filtro conteniendo el sedimento (mm2). |
s |
área del campo visual del microscopio (mm2). |
nPG |
número total de granos contados. |
a |
número de campos contados. |
p |
peso de la miel (g). |
Las muestras se clasificaron por su concentración de polen de acuerdo al esquema de Maurizio (1939), quién las agrupa en cinco clases de acuerdo al número total de granos de polen por cada 10 g de miel: Clase I (≤ 20 × 103), Clase II (21 × 103 ≤ N ≤ 100 × 103),ClaseIII(101×103≤N≤500×103), Clase IV (501 × 103 ≤ N ≤ 106), y Clase V (> 106). Esta clasificación, se toma en cuenta para interpretar los resultados cualitativos, pues permite identificar la sobre o subrepresentación de tipos polínicos en las muestras.
La identificación de los granos de polen se realizó con un microscopio compuesto (Micromaster, Fisher ScientificTM) con objetivo 40X y 100X, por comparación morfológica del polen con la colección palinológica de referencia y con apoyo de claves palinológicas (Kapp et al., 2000; Palacios-Chávez et al., 1991), y atlas polínicos (Reyes et al., 2009). Se identificaron y contaron al menos mil granos de polen por muestra, como lo establece Von der Ohe et al. (2004).
Los tipos polínicos fueron clasificados por su clase de frecuencia, como predominante (P > 45%), secundario (S = 16-45%), de importancia menor (I = 3-15%), polen menor (M < 3%), y polen presente (“+” ≤ 1%). Las mieles se caracterizaron como monoflorales cuando un tipo polínico fue predominante, y multiflorales cuando ningún tipo polínico representó más de 45% del contenido polínico en la muestra (Loveaux et al., 1978).
Resultados
Se identificaron un total de 78 tipos polínicos en las 52 muestras de miel analizadas. Uno de ellos correspondió al tipo morfológico compuesto por las familias Chenopodiaceae y Amaranthaceae [Chen-Am, (Martin, 1963)] y los 77 tipos polínicos restantes fueron de 33 familias botánicas diferentes; de los cuales 47 se identificaron a nivel de especie, 26 a nivel de género y cuatro a nivel de familia.
El número de tipos polínicos identificados varió de ocho a 38 por muestra de miel. La familia mejor representada en diversidad de tipos polínicos, fue Asteraceae (14 tipos), seguida de Fabaceae (10 tipos), Poaceae (cinco tipos) y Boraginaceae y Malvaceae (cuatro tipos cada una). Las familias Fabaceae y Tamaricaceae estuvieron presentes en el 100% de las muestras, la Asteraceae 98%, la Arecaceae y Poaceae 90%, el tipo morfológico Chen-Am 86%, Myrtaceae 80%, Brassicaceae y Salicaceae 76%.
De las 52 muestras de miel analizadas 34 (65%) se clasificaron como mieles monoflorales, de las cuales 20 fueron de Tamarix spp., siete de Prosopis spp., dos de Pluchea sericea, y una muestrade Sysimbrium irio, Myrtaceae, Phoenix dactylifera, Coriandrum sativum y Washingtonia filifera, respectivamente (fig. 2). Las mieles monoflorales de Tamarix spp. se obtuvieron en el segundo periodo de cosecha (agosto) y las mieles monoflorales de Prosopis spp. fueron más frecuentes en el primer periodo de cosecha (junio). Asimismo la mayoría de las mieles monoflorales corresponden a plantas naturalizadas (Tamarix spp., S. irio), o cultivadas (Myrtaceae, P. dactylifera, C. sativum, y W. filifera), y con menor frecuencia a especies nativas (Prosopis spp., y P. sericea). Las fotografías de algunos tipos polínicos registrados en las clases de frecuencias predominantes y secundarios se presentan en la figura 3.
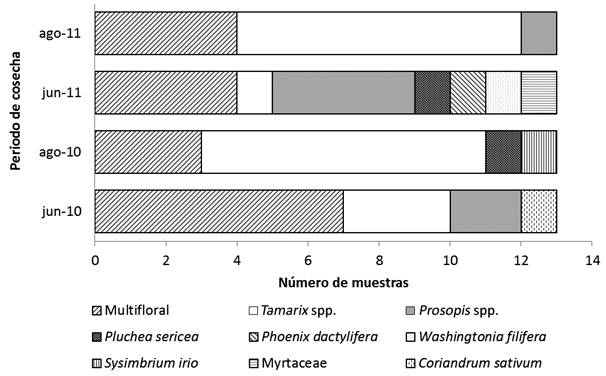
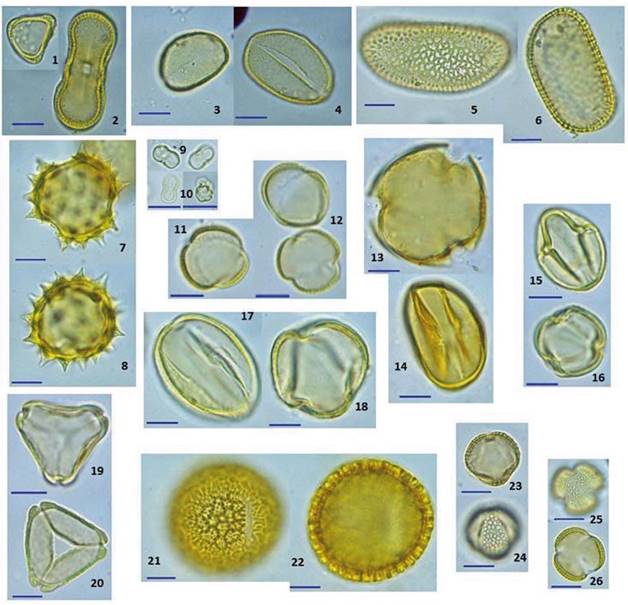
Fig. 3 Tipos polínicos dominantes y algunos secundarios identificados en las mieles del Valle de Mexicali. Apiaceae: 1-2. Coriandrum sativum, Arecaceae: 3-4. Phoenix dactylifera, Arecaceae: 5-6. Washingtonia filifera, Asteraceae: 7-8. Pluchea sericea, Boraginaceae: 9-10. Cryptantha micrantha, Brassicaceae: 11-12. Sysimbrium irio, Fabaceae: 13-14. Medicago sativa, Fabaceae: 15-16. Melilotus indica, Fabaceae: 17-18. Prosopis glandulosa, Myrtaceae: 19-20. Callistemon viminalis, Nyctaginaceae: 21-22. Abronia villosa, Salicaceae: 23-24. Salix gooddingi, Tamaricaceae: 25-26. Tamarix aphylla. La barra en cada foto representa una longitud de 10 micras.
De acuerdo al análisis cuantitativo, una muestra (2%) se ubicó en clase I, 18 (35%) corresponden a la clase II, 27 (52%) a la clase III, dos muestras (4%) a la clase IV, y cuatro (8%) a la clase V (cuadros 1 y 2). En el periodo de agosto 2010 las mieles fueron menos variables en riqueza polínica, presentándose solo dos clases; en cambio en agosto de 2011 fueron más variables presentándose mieles en las cinco clases.
Cuadro 1 Tipos polínicos presentes en muestras de miel del Valle de Mexicali, Baja California, durante dos periodos de cosecha de 2010. Clases de Frecuencia: P = Polen dominante (>45%), S = Polen secundario (16-45%), I = Polen de importancia menor (3-15%), M = Polen menor (>1 - <3%), y + = Polen presente (≤1%). FO = Frecuencia de ocurrencia en el periodo (%).

Cuadro 2 Tipos polínicos presentes en muestras de miel del Valle de Mexicali, Baja California, durante dos periodos de cosecha de 2011.Clases de Frecuencia: P = Polen dominante (> 45%), S = Polen secundario (16-45%), I = Polen de importancia menor (3-15%), M = Polen menor (> 1 - < 3%), y + = Polen presente (≤ 1%). FO = Frecuencia de ocurrencia en el periodo (%).
El análisis cualitativo reveló que 48 tipos polínicos, pertenecientes a 24 familias botánicas tuvieron una frecuencia ≥ 1% del total de granos contabilizados en al menos una de las muestras de miel (cuadros 1 y 2). En contraste, hubo 30 tipos polínicos, que corresponden a 17 familias cuya frecuencia fue < 1%.
Considerando la frecuencia de ocurrencia (porcentaje sobre el total de las muestras analizadas en la cual el tipo polínico aparece), sobresale el de Tamarix spp. (pino salado), que se encontró en el 100% de las mieles analizadas, seguido de Prosopis spp. (mezquite y tornillo), Medicago sativa (alfalfa), P. sericea (Cachanilla) y el tipo Chen-Am (Chamizos y quelites) presentes en el 92% (48), 90% (47) y 87% (45) de las muestras analizadas respectivamente. Otros tipos polínicos con alta frecuencia de ocurrencia fueron Myrtaceae (eucaliptos y cepillo rojo), en 81% (42 mieles), seguido por W. filifera (palma de abanico), S. irio (mostacilla), Poaceae 1 (pastos), y S. gooddingi (sauz), cada uno presente en 76% (38) de las muestras analizadas (cuadros 1 y 2). De los tipos polínicos hallados con valores bajos en las muestras (< 1%), los que tuvieron mayor frecuencia de ocurrencia fueron Gossypium hirsutum, Baccharis salicifolia, Acacia saligna, Cucumis melo, Malvella leprosa, Heterotheca sp., Heliotropium curassavicum, Opuntia sp., y Populus fremontii (cuadros 1 y 2).
Se encontró que el tipo polínico que resultó predominante en el mayor número de muestras fue Tamarix spp., además este tipo polínico fue el único que tuvo frecuencia de ocurrencia del 100% en las 52 muestras analizadas (cuadros 1 y 2).
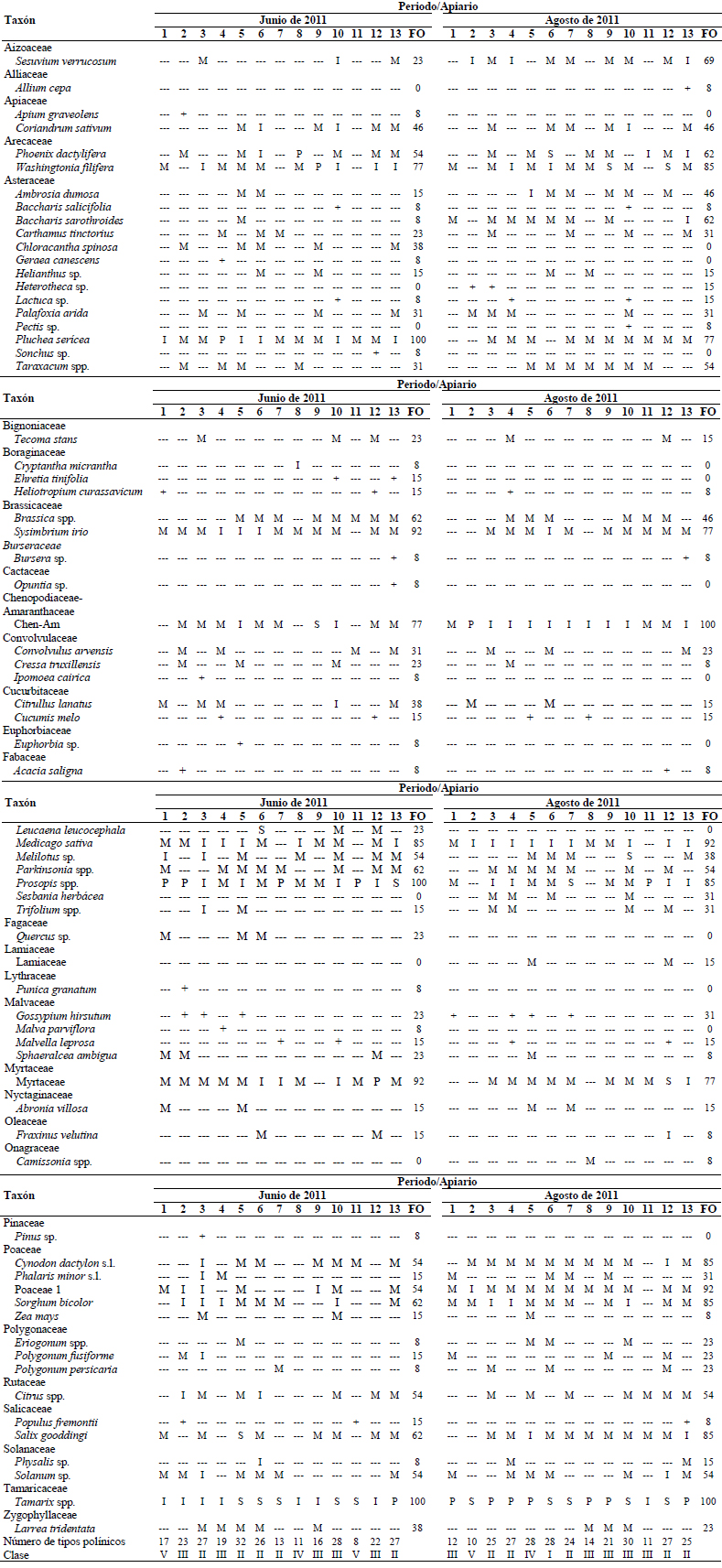
En las muestras de miel colectadas durante junio de 2010 se identificaron 62 tipos polínicos, pertenecientes a 25 familias botánicas. Seis de las trece muestras fueron clasificadas como monoflorales: tres de ellas de Tamarix spp., dos de Prosopis spp. y una de C. sativum. Otros taxones con clases de frecuencia altas ≥ 16% fueron C. sativum, P. sericea, C. micrantha, Melilotus sp., Prosopis spp., A. villosa y Tamarix spp. Los tipos polínicos con mayor frecuencia de ocurrencia fueron P. sericea, Prosopis spp. y Tamarix spp., presentes en las trece muestras del periodo, seguidos por P. dactylifera, M. sativa y Melilotus sp., que estuvieron presentes en 11 muestras.
Mediante el análisis de las muestras obtenidas el mes de agosto de 2010 fueron identificados 55 tipos polínicos correspondientes a 24 familias botánicas. Diez de las trece muestras se clasifican como monoflorales: ocho de ellas de Tamarix spp. una de S. irio y otra de P. sericea. Los taxones clasificados por clase de frecuencia como polen secundario (≥ 16%) fueron M. sativa, A. villosa, Tamarix spp., Prosopis spp., P. sericea y el tipo Chenopodiaceae-Amaranthaceae. Aquellos tipos polínicos con las más altas frecuencias de ocurrencia fueron Tamarix spp. y M. sativa, encontrados en las 13 muestras, Chen-Am en 12, y tanto Prosopis spp. como S. gooddingui estuvieron presentes en 11 muestras.
En las muestras correspondientes a junio de 2011 se identificaron 64 tipos polínicos, correspondientes a 26 familias botánicas. Nueve de las 13 muestras se clasifican como monoflorales, cuatro de ellas con polen predominante de Prosopis spp., mientras que las cinco muestras monoflorales restantes son de P. sericea, P. dactylifera, W. filifera, Tamarix spp., y Myrtaceae, respectivamente. Los taxones secundarios (≥ 16%) fueron Tamarix spp., Prosopis spp., L. leucocephala, S. gooddingi y el tipo polínico Chenopodiaceae-Amaranthaceae. Al igual que en mismo periodo del año anterior, los tres tipos polínicos con mayor frecuencia de ocurrencia fueron P. sericea, Prosopis spp. y Tamarix spp., estando presentes en las trece muestras; pero en esta ocasión Myrtaceae y S. irio estuvieron presentes en 12 de las muestras, mientras que M. sativa lo estuvo en 11.
Los tipos polínicos identificados en las muestras cosechadas en agosto de 2011 fueron 56, correspondientes a 25 familias botánicas. Nueve de las trece muestras se clasificaron como monoflorales, de las cuales en siete el polen predominante fue de Tamarix spp.; se obtuvo una muestra de Prosopis spp., y otra de Chenopodiaceae-Amaranthaceae. Los tipos polínicos secundarios (≥16%) fueron Tamarix spp., P. dactylifera, W. filifera, Melilotus sp., y Myrtaceae. Respecto a la frecuencia de ocurrencia, el tipo polínico Chen-Am, al igual que Tamarix spp. estuvieron presentes en 13 muestras del periodo, mientras que Poaceae y M. sativa fueron identificadas en 12 muestras; también se encontró con alta frecuencia de ocurrencia Prosopis spp., W. filifera, S. gooddingi, Sorghum bicolor y Cynodon dactylon s.l., cada uno de ellos presente en 11 muestras.
En las figuras 4, 5, 6 y 7 se muestran los valores porcentuales de los pólenes predominantes y secundarios en cada muestra de miel por periodo de cosecha. Se observa que Tamarix es el tipo mejor representado en todos los periodos, principalmente en las mieles del mes de agosto de ambos años, seguido por Prosopis, que se clasifica como polen predominante y secundario en las mieles cosechadas en junio, pero no así en las mieles de agosto, en tercer lugar se encuentra P. sericea que fue predominante en dos muestras, una de agosto de 2010 y otra de junio de 2011, sin embargo no fue predominante o secundario en las muestras recolectadas en agosto de 2011, este taxón se identificó en el 100% de las muestras obtenidas en junio de ambos años, y en el 87% del total. El polen de M. sativa sólo fue taxón secundario en cuatro muestras de agosto de 2011, aunque su frecuencia de ocurrencia es alta (90%) estando presente en 47 de las 52 muestras.
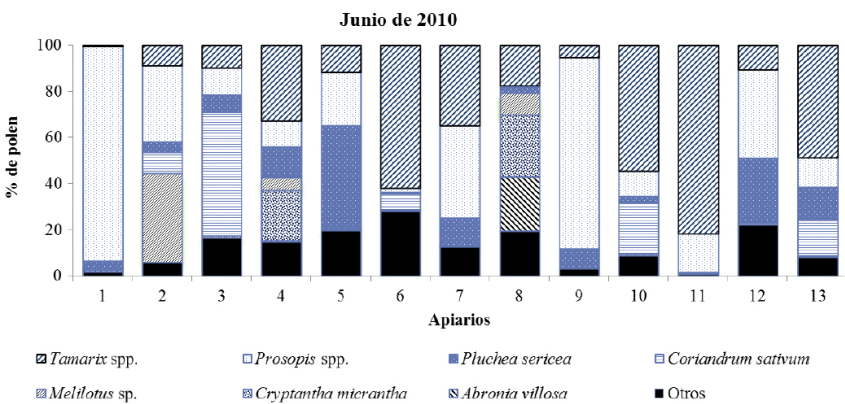
Fig. 4 Tipos polínicos predominantes y secundarios presentes en las muestras de miel cosechadas en junio de 2010 en el Valle de Mexicali (México).
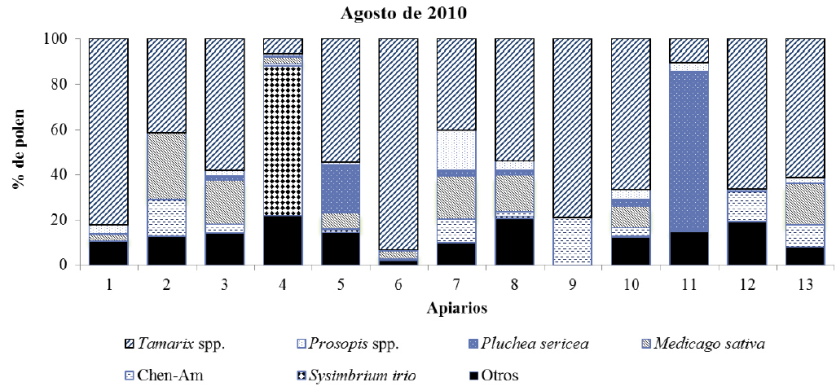
Fig. 5 Tipos polínicos predominantes y secundarios presentes en las muestras de miel cosechadas en agosto de 2010 en el Valle de Mexicali (México).
Fig. 6 Tipos polínicos predominantes y secundarios presentes en las muestras de miel cosechadas en junio de 2011 en el Valle de Mexicali (México).
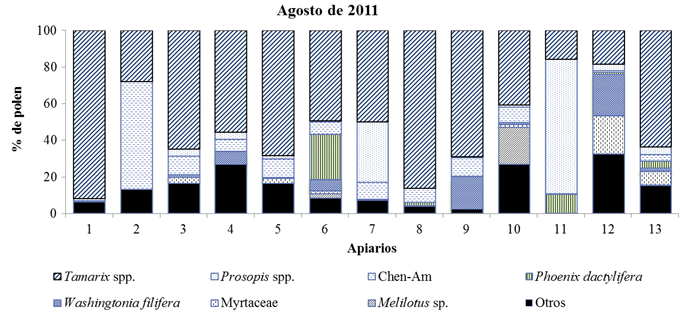
Fig. 7 Tipos polínicos predominantes y secundarios presentes en las muestras de miel cosechadas en agosto de 2011 en el Valle de Mexicali (México).
Otros tipos polínicos sobresalientes por su porcentaje en algunas de las muestras fueron: Chen-Am, que presentó polen predominante en una muestra de agosto de 2011 y secundario en una muestra de agosto de 2010 y en otra de junio de 2011; también P. dactylifera, fue predominante en una muestra de junio de 2011 y secundario en otra de agosto del mismo año; también en las mieles de 2011 W. filifera fue polen predominante en una muestra de junio y secundario en dos muestras de agosto, una de ellas en el mismo sitio donde previamente había resultado como predominante; lo mismo ocurrió con el polen de Myrtaceae, que siendo predominante en una muestra de junio 2011, para el mismo apiario resultó como secundario en la muestra de agosto del mismo año. El caso de C. sativum, fue secundario en una muestra y predominante en otra de las muestras obtenidas en junio de 2010, sin haber llegado a ser predominante o secundario en ninguna muestra de otro periodo. Melilotus sp. resultó polen secundario en dos muestras: una correspondiente a junio de 2010 y otra recolectada en agosto de 2011. S. irio fue hallado como polen predominante en una muestra de agosto de 2011. Finalmente, cuatro tipos polínicos aparecieron como secundarios en una sola de las muestras, éstos correspondieron a: S. gooddingui, C. micrantha, L. leucocephala, y A. villosa.
Discusion
Éste es el primer trabajo que describe los tipos polínicos presentes en las mieles de Mexicali, y en el estado de Baja California.
Aunque existen referencias bibliográficas sobre la importancia nectarífera de algunas especies vegetales presentes en el área de estudio (Pellet, 1947; Wiggins, 1980; Crane, 1990; Rebman y Roberts, 2012), en general estas referencias corresponden a observaciones de visitas de abejas a las flores, sin embargo mediante la identificación de los granos de polen de estos recursos nectaríferos en mieles, y otros de los cuales no existían referencias previas, se confirma su importancia para la apicultura en la región.
En las mieles de Mexicali se observa una diversidad de tipos polínicos relativamente baja comparada con otros estudios (Villanueva-Gutiérrez, 1994; Ramírez-Arriaga et al., 2011; Castellanos-Potenciano et al., 2012; Costa et al., 2013; Jones y Bryant, 2014), ello se explica, en parte, por la relativa homogeneidad del área de estudio y la baja diversidad del hábitat, ocasionada por la sustitución de la vegetación riparia por especies invasivas como Tamarix spp. (pino salado) y cultivos agrícolas (Delgadillo, 1998), ya que con frecuencia la diversidad del área donde fue producida la miel se refleja en la diversidad polínica que ésta contiene (González-Porto et al., 2013). También, el espectro polínico refleja un elevado nivel de intervención humana en el área de estudio (Fernandes y Barth, 2012).
Este estudio demostró la importancia de Tamarix spp. como fuente de néctar, sobre todo en el segundo periodo de cosecha (agosto), lo cual es congruente con lo observado en campo, ya que la floración de la planta ocurre durante toda la época de producción de miel (mayo a septiembre). Se observó que en agosto se reduce la floración de otras especies vegetales, como sucede en Prosopis spp., a medida que se aproximan los meses más cálidos del año en el Valle de Mexicali (julio y agosto). La dominancia de Tamarix spp. en muestras de miel de otras áreas geográficas ha sido reportada por Forcone et al. (2003), quienes encontraron que Tamarix gallica fue una de las principales fuentes de néctar en la producción de miel en el Valle inferior del Río Chubut (Argentina), tratándose también de una región donde la vegetación original había sido intervenida.
Las familias mejor representadas por su diversidad de tipos polínicos en las mieles analizadas fueron Asteraceae y Fabaceae, lo cual es consistente con lo reportado por Villanueva-Gutiérrez (1984), Crane (1991), Forcone et al. (2003) y Costa et al., (2013). Sin embargo en relación a las clases de frecuencia más elevadas, la familia mejor representada fue Tamaricaceae, seguida de Fabaceae y Asteraceae (cuadros 1 y 2).
Los resultados del análisis cuantitativo revelaron que el 52% de las mieles del Valle de Mexicali se ubicaron en el grupo III (27 muestras), lo que indica que proceden de plantas altamente productoras de polen. Además el 34% de las muestras se clasificaron en el grupo II (18 muestras), indicando que proceden de plantas que producen una cantidad normal de polen (Loveaux, et al., 1978; Jones y Bryant, 1996; Von der Ohe et al., 2004).
El 11.5% de las muestras de miel se clasificaron en los grupos IV y V, indicando que provienen de plantas con néctares extremadamente ricos en polen (Loveaux et al., 1978; Jones y Bryant, 1996; Von der Ohe et al., 2004). Sin embargo existe evidencia que las mieles pertenecientes a estos grupos pueden indicar contaminación con celdas de polen (Loveaux et al., 1978). Lo que explicaría los resultados obtenidos, ya que en esta investigación se buscó recolectar solamente trozos de panal que contuvieran miel, pero es posible que algunas celdas contuvieran polen, lo que pudo sobrerepresentar el contenido de polen en las muestras de miel estudiadas.
De acuerdo al origen botánico, la mayoría de las mieles producidas en Mexicali pueden considerarse como monoflorales, principalmente aquellas con dominancia de polen de Tamarix spp. que pese a tratarse de un grupo de especies introducidas e invasivas (Rebman y Roberts, 2012), resultan ser el recurso nectarífero más importante para el Valle de Mexicali. Pero desde el punto de vista comercial, resulta más atractiva la posibilidad de ofrecer mieles cuyo polen predominante proceda de la vegetación nativa (Prosopis spp. y P. sericea), y en segundo término las que proceden de cultivos regionales (C. sativum y posiblemente M. sativa).
En este estudio se encontraron contenidos elevados de polen de M. sativa, sin embargo las mieles monoflorales de M. sativa tienden a tener bajos contenidos totales de polen (Forcone, 2008), lo que difiere a nuestros resultados. Probablemente este alto contenido de polen se haya debido a una contaminación secundaria, mediante celdas con polen de este taxón por lo tanto no se consideraron mieles monoflorales de M. sativa.
Se conoce que no todos los taxa contribuyen en la misma proporción a la producción de miel (Jones y Bryant, 2014), ante lo cual debería considerarse el uso de los llamados “coeficientes de polen”, cuyo uso no es generalizado e implica amplio conocimiento
de la producción relativa de polen de cada especie, así como del comportamiento de las abejas y su capacidad para filtrarlos (Bryant y Jones, 2001). Para muchos de los tipos polínicos encontrados en las mieles no existen estos coeficientes de polen, y se consideró que para la correcta denominación botánica de las mieles se deben tomar en cuenta también parámetros fisicoquímicos y organolépticos (Von der Ohe et al., 2004). Por lo anterior, en el presente estudio no se aplicaron algunos criterios generalmente aceptados como excepciones en la denominación botánica de las mieles, al considerar que se requieren más análisis a las mieles de la región.
Por su frecuencia de ocurrencia, y por sus clases de frecuencia, Tamarix spp., Prosopis spp., y P. sericea se pueden considerar las principales fuentes de néctar y elementos representativos de las mieles del Valle de Mexicali.
Los elevados valores de polen de Poaceae, así como su alta frecuencia de ocurrencia, se relacionarían con una contaminación secundaria -producida dentro de la colmena, por una alta recolección de este recurso como fuente de polen-. Especies anemófilas o plantas sin nectarios, como las Poaceae llegan a ser fuente de polen en épocas de escasez, y forman parte integral del uso que las abejas hacen de los recursos vegetales y como elementos presentes y característicos de mieles producidas en regiones geográficas determinadas (Sajwani et al.,2007; Jones y Bryant, 2014; Ponnuchamy et al., 2014).
Este estudio reveló baja representación de polen de Gossypium hirsutum en las muestras de, miel del Valle de Mexicali, a pesar de la gran superficie sembrada con estecultivo en la zona de estudio (32 000 ha, SIAP, 2011). Esta planta se considera una especie que produce néctar en abundancia (Crane, 1990), cuyas flores son visitadas con frecuencia por A. mellifera (Pise y Viraktamath, 2015) y llegan a producirse mieles monoflorales (Persano-Oddo et al., 2004), aun cuando su productividad no depende de la polinización por abejas (Barfield et al., 2015). Esta baja presencia se podría explicar en parte debido a que el grano de polen de G. hirsutum es relativamente grande (Pise y Viraktamath, 2015) y en el transporte del néctar a la colmena las abejas filtran muchos de los granos de polen (Todd y Vansell, 1942). Adicionalmente, el uso frecuente de agroquímicos, como imidacloprid (Keefer y Gold, 2014), podría reducir la visita de las abejas a este cultivo, debido a efectos subletales (Schneider et al., 2012). Y finalmente, las variedades transgénicas de algodón que se utilizan localmente podrían no ser atractivas para las abejas. No obstante, en otras áreas geográficas no se ha reportado que existan diferencias entre el número de visitas de abejas a flores de algodón transgénico y no transgénico (Pierre y Hofs, 2010; Pise y Viraktamath, 2015).
Conclusiones
Los principales recursos néctar poliníferos usados por A. mellifera en el Valle de Mexicali corresponden a especies introducidas, tanto plantas naturalizadas como cultivos agrícolas, con presencia de Tamarix spp. en todas las mieles analizadas, así como algunos elementos representativos de la vegetación original, como Prosopis spp. y Pluchea sericea. Otras especies importantes en la producción de miel son Coriandrum sativum, Phoenix dactylifera, Washingtonia filifera y Sysimbrium irio.
El 65% de las mieles cosechadas en el Valle de Mexicali pueden describirse como monoflorales, y de éstas más de la mitad contienen como polen predominante el de Tamarix spp.; también se producen mieles monoflorales de Prosopis spp., Pluchea sericea, y en menor medida de Coriandrum sativum, Washingtonia filifera, Phonenx dactylifera, Myrtaceae y Sysimbrium irio. La mayor parte de las mieles monoflorales de Prosopis spp. se obtienen durante la primera cosecha (junio) y en la segunda cosecha (agosto) predominan las mieles de Tamarix spp. Los taxones Tamarix spp., Prosopis spp, y Pluceha sericea constituyen los elementos representativos de las mieles analizadas.

