











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Enterolobium cyclocarpum (Jacq.) Griseb (Parota, Guanacastle, Orejón) es una especie arbórea apreciada para artesanías, canoas, carpintería, chapa, decoración de interiores, duela, ebanistería, lambrín, muebles, utensilios de cocina, forraje (hojas y frutos), en medicina tradicional (corteza) y en la alimentación humana (semillas) (Román et al., 2011; Pineda-Herrera et al., 2012c).
Es frecuente en la vegetación perturbada de las zonas tropicales húmedas y subhúmedas de baja altitud de México y Centroamérica. En la República Mexicana se distribuye en la vertiente del Pacífico y la Cuenca del Balsas, en el Golfo de México, península de Yucatán y depresión central de Chiapas. Se le encuentra en asociaciones primarias de selvas medianas subcaducifolias y caducifolias, así como en zonas perturbadas de selvas altas perennifolias y medianas subperennifolias (Pennington y Sarukhán, 2005).
Siendo un elemento frecuente en varios tipos de selvas, su repoblación y desarrollo están fuertemente influenciados por el clima, aunque debe considerarse: la competencia por factores bióticos y abióticos, el estado ontogenético y aspectos funcionales que pueden direccionar su fenología, la actividad de su cambium vascular y su crecimiento (Worbes, 1995; Van der Sande et al., 2015). Todo ello es indispensable para entender las estrategias de colonización y supervivencia de los árboles en condiciones naturales (Callado, 2010).
La fenología de E. cyclocarpum ha sido compilada en las monografías de Vázquez-Yanes et al. (1999) y Orwa et al. (2009), en las investigaciones de Borchert (1994b), Borchert et al. (2004), Rojas-Jiménez et al. (2007), Moreno-Casasola et al. (2010) y Estrada-Castelo et al. (2012). Es necesario señalar que dichos estudios se han realizado en selvas y sabanas a altitudes menores a 150 metros, por lo que se confirma que en México son incipientes los estudios fenológicos sobre E. cyclocarpum en selvas de altitud mayor.
Los estudios sobre crecimiento de E. cyclocarpum se han realizado en Centroamérica (Reich y Borchert, 1984; Borchert, 1994b) en selvas secas de altitudes bajas y en una selva mediana subcaducifolia del Pacífico Mexicano (Makocki et al., 2012). Dichas investigaciones señalan diferencias entre sitios respecto a la influencia del clima en las fluctuaciones del diámetro y sólo este último trabajo cita que su crecimiento es anual y favorecido por la posición de su copa.
E. cyclocarpum es una especie representativa en la sucesión de selvas húmedas y secas en México, ello representa una oportunidad para estudiar estos temas en regiones donde no se han hecho investigaciones al respecto. Considerando lo anterior, se plantean las siguientes preguntas: 1) ¿En una selva seca, sus eventos fenológicos están influenciados únicamente por la dinámica de la precipitación? 2) ¿Árboles de diferentes dimensiones tienen tasas de crecimiento en diámetro distintas? 3) ¿Árboles con diferentes dimensiones y posiciones de copa crecen de forma distinta? 3) ¿El tamaño de los árboles influye en su fenología? 4) ¿En selvas medianas localizadas a mayor altitud se forma un solo anillo anual de crecimiento?
En Costa Grande, Guerrero, la región estatal de mayor extensión territorial (14 710.70 km2), es posible encontrar poblaciones de E. cyclocarpum en paisajes selváticos perturbados y/o dedicados a la actividad agropecuaria. No obstante que la región es una de las que mayor tradición maderable tiene en el estado y en el país, muchas especies de selvas no son aprovechadas adecuadamente a pesar de su potencial productivo y calidad maderable, debido en parte a la carencia de estudios sobre la edad y crecimiento de los árboles (Makocki et al., 2012; Pineda-Herrera et al., 2012a).
Los objetivos fueron: 1) evaluar la caída y el brote de hojas, la floración y la fructificación de E. cyclocarpum de marzo de 2010 a julio de 2012 en correlación con la precipitación y temperatura, 2) medir de marzo de 2010 a julio de 2012 el crecimiento en diámetro del fuste para establecer diferencias entre categorías diamétricas, simetrías e iluminaciones de copa, 3) establecer la relación entre diámetro e intensidad fenológica y 4) determinar la periodicidad de formación de sus anillos de crecimiento para el periodo marzo de 2010-julio de 2012.
Material y métodos
Área de studio
Se localiza en “Los Varillos” (440 m snm) en el ejido “La Laja y anexos” (17° 53’ 58’’ N, 101° 26’ 68” O) (INEGI, 2009), municipio de Zihuatanejo de Azueta, Costa Grande, Guerrero, provincia Sierra Madre del Sur y subprovincia Cordillera Costera del Sur. Geomorfológicamente Los Varillos es una meseta rodeada por laderas con pendientes pronunciadas y lomeríos moderados (INEGI, 2009). El clima es cálido subhúmedo con lluvias en verano (Aw): con una precipitación total anual de 1100 a 1500 milímetros y una temperatura media anual de 16 a 28°C (INEGI, 2009).
Se presentan afloramientos de rocas ígneas extrusivas, metamórficas y sedimentarias (INEGI, 2009). Los suelos son de origen volcánico de textura migajón arcillosa-arenosa (INEGI, 2009). Es una selva mediana subcaducifolia caracterizada por especies arbóreas como: Astronium graveolens Jacq., Brosimum alicastrum Swarts., Bursera simaruba (L.) Sarg., Ceiba aesculifolia (Kunth) Britton & Rose, Cordia elaeagnoides A. DC., Enterolobium cyclocarpum (Jacq.) Griseb, Ficus petiolaris Kunth, Hymenaea courbaril L., Licania arbórea Seem., Lysiloma acapulcensis ((Kunth) Benth., Swietenia humilis Zucc., Tabebuia rosea (Bertol.) DC.
Las alturas son en promedio de 20 a 25 m, con algunos ejemplares emergentes que alcanzan los 30 m (B. alicastrum) (Pineda-Herrera et al., 2012a).
Fenología
Se identificaron en la localidad, 15 árboles sanos con diámetros de 20 a 50 cm, de fuste recto y en edad reproductiva (Gómez, 2010). Bimestralmente desde marzo de 2010 a julio de 2012 se registró utilizando binoculares en cada uno de ellos: a) caída de hojas, b) flores, c) frutos y d) brote de hojas; todos expresados en una escala del cero al cuatro según el porcentaje de presencia de cada evento: Ausencia total del evento (0), 1-25% (1), 26-50% (2), 51-75% (3) y 76-100% (4) (Pineda-Herrera et al., 2012b). Los valores promedio obtenidos en campo se correlacionaron (Spearman) (Gotelli y Ellison, 2004) con la temperatura y precipitación promedios, obtenidos para los periodos de observación de la estación Vallecitos de Zaragoza del Servicio Meteorológico Nacional (SMN, 2012) (17° 55’ 20’’ N, 101° 19’ 02” O) que se localiza a 20 km de distancia de la localidad. El software estadístico utilizado fue STATA (STATA Corp., 2001).
Con el propósito de establecer la relación entre diámetro (DAP = 1.30 m del suelo) de los 15 árboles y la fenología, se categorizaron en seis intervalos: 15.0-25.0 cm (categoría 20); 25.1-35.0 cm (categoría 30), 35.1-45.0 (categoría 40), 45.1-55.0 cm (categoría 50). Se promedió el porcentaje por cada evento, por bimestre, multiplicando cada valor por 100 y dividiéndolo entre 4 y se graficó de acuerdo a su diámetro para el periodo de observación marzo 2010-julio 2012.
Con los valores fenológicos se realizó una regresión lineal en STATA (STATA Corp., 2001) con cada categoría diamétrica a través del siguiente modelo (ecuación 1):
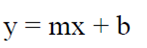 (1)
(1)
donde:
y |
porcentaje del evento fenológico de acuerdo a su categoría diamétrica |
m |
coeficiente de la regresión |
x |
diámetro (DAP) en cm |
b |
constante |
La bondad de ajuste del modelo, fue evaluada a través de los valores estadísticos de su coeficiente de determinación (R2) y el nivel de significancia (p ≤ 0.01).
Incremento en diámetro
Se midió el diámetro de 18 árboles a 1.30 m (DAP) encontrados por orden de aparición en la selva, distintos a los elegidos para el estudio de fenología. Bimestralmente desde marzo del 2010 hasta julio del 2012 se midieron empleando bandas dendrométricas (López-Ayala et al., 2006a; Makocki et al., 2012). Los árboles se clasificaron en seis categorías diamétricas (tres árboles por cada categoría): 2.5-7.49 cm (categoría 5), 7.5-12.49 cm (categoría 10), 12.5-17.49 cm (categoría 15), 17.7-22.49 cm (categoría 20), 22.5-27.49 cm (categoría 25) y 27.5-32.49 cm (categoría 30).
La definición de los límites inferior y superior de cada categoría, estuvo determinada por la disponibilidad de individuos. Estos límites se establecen con el propósito de obtener la dinámica de crecimiento por estructura de edades de la población a estudiar (Melo y Vargas, 2003).
Las copas de los árboles medidos se categorizaron modificando la propuesta de Manzano et al. (2010): Simetría o calidad de copa. 1: copa asimétrica, degradada, dañada, con una o pocas ramas 2: copa parcialmente simétrica o rala, con medio círculo o medio óvalo 3: copa simétrica, forman un círculo completo, perfecto. Iluminación de copa. 1: recepción de luz vertical o lateral poca, enteramente sombreada 2: recepción vertical o lateral parcial, sombreada parcialmente por otras copas 3: recepción vertical o lateral total, la parte superior de la copa totalmente expuesta a la luz y libre de competencia lateral.
Los incrementos por categoría y condición de copa se ajustaron al modelo no lineal Gompertz (ecuación 2), el cual fue elegido dadas las cualidades del ajuste como: suma de cuadrados del error (SCE), coeficiente de determinación (R2) y nivel de confianza estadística (p ≤ 0.01).
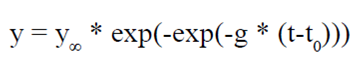 (2)
(2)
donde:
y |
variable de estudio (diámetro) |
y∞ |
parámetro de valor asintótico |
g |
parámetro de significado biológico limitado |
t0 |
parámetro que determina la tendencia de la curva en el tiempo |
t |
edad (días) |
La modelación de los incrementos se realizó en el paquete STATA en el módulo de análisis de regresión no lineal (STATA Corp. 2001). La comparación estadística entre las categorías diamétricas y las condiciones de copa en el paquete PAC (Programas para el análisis de crecimiento) (Salgado-Ugarte et al., 2005) mediante la prueba de razón de verosimilitud (Kimura, 1980) que calcula un estadístico que es comparado con la distribución χ2 con grados de libertad iguales al número de restricciones, de acuerdo a la ecuación 3.
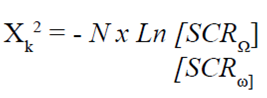 (3)
(3)
donde:
k |
son los grados de libertad. |
N |
es el número total de observaciones combinado de las dos curvas que se están comparando. |
SCRΩ |
es la suma total de cuadrados de los residuos obtenida de ajustar ambas curvas por separado. |
SCRω |
es la suma total de residuos cuadrados derivados de ajustar a las curvas con algunas de las restricciones supuestas (Ys iguales, gs iguales, ts iguales). |
La pareja de curvas con diferencias estadísticas significativas presentarán valores más altos de razón de verosimilitud (RV) (Salgado-Ugarte et al., 2005).
Periodicidad de anillos de crecimiento
Se eligió un árbol de categoría diamétrica 15 cm. Detienne (1989) recomienda que para una identificación confiable de anillos sean elegidos árboles jóvenes con la finalidad de disminuir la probabilidad de encontrar anillos falsos o discontinuos. Al árbol elegido se le realizaron incisiones de aproximadamente 3 mm de ancho, 5 mm de profundidad y 100 mm de longitud a una altura de 1.30 m (DAP) cada dos meses, de marzo de 2010 a abril de 2012. En esta última visita, el árbol se derribó para obtener la rodaja correspondiente, la cual fue cepillada y pulida con una lijadora orbital Trupper mod. ¼ Aen una secuencia de grados: 80, 120, 150, 220, 240, 320, 400 hasta distinguir los anillos y establecer la periodicidad en su formación (López-Ayala et al., 2006b; Galán-Larrea et al., 2011).
Para observar microscópicamente las estructuras celulares que delimitaron los anillos de crecimiento, la rodaja se sumergió en agua destilada durante 72 horas para ablandarla. De la rodaja se obtuvo un cubo de 1 x 1 cm, al cual se le realizaron cortes de aproximadamente 20 μm de grosor con un micrótomo Zeiss (Hyrax 530). Los cortes se tiñeron con fucsina básica al 0.05% durante 30 segundos y después en azul astra al 0.5% durante dos minutos. Se deshidrataron en una serie gradual de alcoholes del 50, 70, 85, 96 y 100%. Después de deshidratados se colocaron en xilol y se montaron con resina sintética (modificado de Sandoval, 2005).
Finalmente se observaron en un microscopio óptico marca Zeiss (Axiostar plus) a aumentos de 5x para la delimitación de anillos y a 20x para la identificación de estructuras celulares delimitantes. El tipo de porosidad y parénquima se clasificó de acuerdo con IAWA Committee (1989).
Resultados
Fenología
En el cuadro 1 se desglosa el desarrollo de los cuatro eventos fenológicos de E. cyclocarpum en el área de estudio. La caída de hojas, la floración y la fructificación se presentaron en dicho orden durante los últimos meses y durante el primer y segundo trimestres de los años evaluados. El brote de hojas se presentó con mayor intensidad durante el segundo semestre de los años de estudio.
Cuadro 1 Calendario fenológico de Enterolobium cyclocarpum.
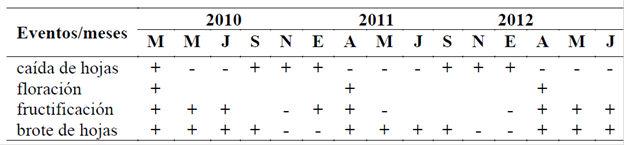
Intensidad del evento: + máxima - mínima
La caída de hojas fue un evento conspicuo, ya que un 95% de la copa de los individuos observados presentó caída de su follaje. La floración se caracterizó por las cabezuelas compactas de color blanco. Los frutos observados en el estudio son vainas evidentes y reconocibles por su tamaño y estructura. Las hojas bipinnadas nuevas se caracterizan por el verde lustroso que adquieren (fig. 1).

Fig. 1 Eventos fenológicos de Enterolobium cyclocarpum. A. árbol sin hojas. B floración. C. fructificación. D. árbol con hojas.
La figura 2 muestra que la caída de hojas tiene sus mayores picos en la época seca, con el aumento de la temperatura. La floración se presentó antes de las lluvias, con las mayores temperaturas del año principalmente en la primavera de 2011. La fructificación se presentó en época de secas en los años 2010, 2011 y 2012, y en los inicios de la época de lluvias (2011 y 2012). Los máximos valores del brote de hojas coinciden con la temporada de precipitación y aumentos de la temperatura a partir del mes de marzo en los tres años evaluados.

Fig. 2 Dendrofenograma de Enterolobium cyclocarpum. Eventos fenológicos: A. caída de hojas. B. floración. C. fructificación. D. brote de hojas.
El brote de hojas se correlacionó de forma significativa (P ≤ 0.01) con la temperatura. La fenología foliar y reproductiva, así como la caída de hojas no se asociaron con la precipitación ni con la temperatura (cuadro 2).
Cuadro 2 Coeficientes de correlación (Spearman) entre precipitación/temperatura
| Variable | Caída de hojas | Floración | Fructificación | Brote de hojas |
|---|---|---|---|---|
| precipitación | -0.3068 | -0.2775 | -0.3551 | 0.4113 |
| temperatura | -0.4811 | -0.2804 | 0.4228 | 0.6868* |
*P ≤ 0.01
La caída de hojas no se asocia a fustes más gruesos, mientras que la floración lo hace de forma significativa. La fructificación presentó los mayores valores fenológicos y una mediana asociación con diámetros mayores. El brote de hojas es la fenofase con mayor asociación significativa a árboles con mayores fustes (fig. 3).
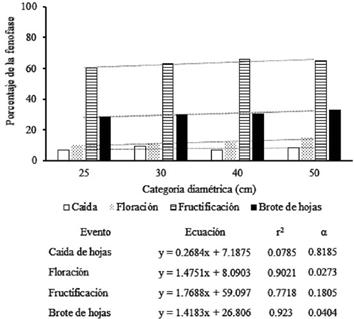
Incremento en diámetro
El ajuste general al modelo de crecimiento mostró altos valores estadísticos y significativos (P ≤ 0.01). Los árboles presentaron incrementos en las épocas de lluvias de 2010 y 2011, así como en el inicio del periodo en 2012 (fig. 4).
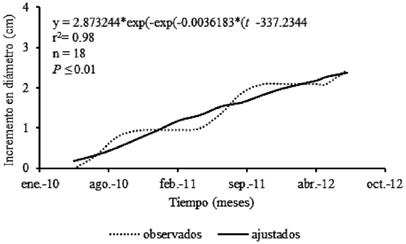
Fig. 4 Crecimiento de todos los árboles de Enterolobium cyclocarpum (•••• datos observados, ▬ ajuste).
La figura 5 muestra que el crecimiento de cada categoría diamétrica en orden descendente fue de: 30, 25, 20, 15, 10 y 5, los valores máximos para todo el periodo de medición superaron los 4 cm en la categoría 30 cm y fueron menores a 1 cm en el caso de la categoría 5 cm.
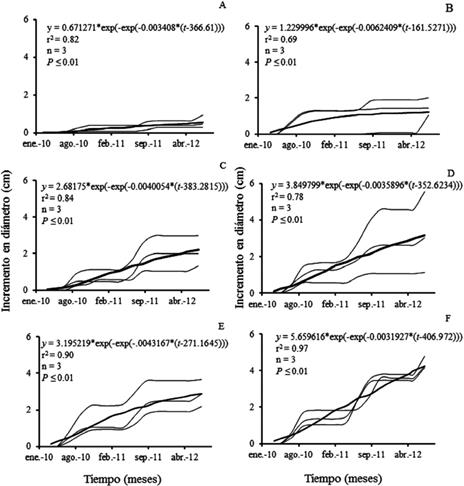
Fig. 5 Crecimiento en diámetro para las categorías diamétricas (A)5, (B)10, (C)15, (D)20, (E)25 y (F)30 cm de Enterolobium cyclocarpum (― datos observados, ▬ ajuste)
Los incrementos promedios anuales fueron mayores en las categorías 25 y 30 cm, seguidos por 20, 10, 15 y 5. Los valores mínimos son similares para 30 y 25, seguidos por 20, 15, 10 y 5 (cuadro 3).
Cuadro 3 Crecimiento en promedio anual de los árboles de Enterolobium cyclocarpum.
| Categoría diamétrica | Número de árboles | Crecimiento promedio anual (cm) | Desviación estándar | ||
|---|---|---|---|---|---|
| Mínimo | Medio | Máximo | |||
| 5 | 3 | 0.02 | 0.12 | 0.23 | 0.11 |
| 10 | 3 | 0.34 | 0.51 | 0.85 | 0.39 |
| 15 | 3 | 0.15 | 0.54 | 0.72 | 0.33 |
| 20 | 3 | 0.38 | 0.69 | 1.17 | 0.54 |
| 25 | 3 | 0.45 | 0.81 | 1.41 | 0.64 |
| 30 | 3 | 0.46 | 1.05 | 1.41 | 0.67 |
| Todas | 18 | 0.30 | 0.62 | 0.96 | 0.44 |
La prueba de verosimilitud mostró diferencias significativas para nueve interacciones a un nivel de significancia de p ≤ 0.001 y una (15 y 25 cm) a nivel p ≤ 0.05 (cuadro 4). Las interacciones entre categorías 10 y 15, 15 y 20, 20 y 25, 20 y 30, 25 y 30 fueron iguales (cuadro 4) y además tienen similitudes entre alguno de sus incrementos mínimo, promedio o máximo (cuadro 3).
Cuadro 4 Prueba de razón de verosimilitud para categorías diamétricas de Enterolobium cyclocarpum.
| Categorías | RV | gl | P |
|---|---|---|---|
| 5 y 10 | 25.079 | 3 | 0.0001 |
| 5 y 15 | 32.240 | 3 | 0.0001 |
| 5 y 20 | 38.233 | 3 | 0.0001 |
| 5 y 25 | 58.888 | 3 | 0.0001 |
| 5 y 30 | 63.911 | 3 | 0.0001 |
| 10 y 15 | 2.5075 | 3 | 0.474 |
| 10 y 20 | 12.787 | 3 | 0.005 |
| 10 y 25 | 19.506 | 3 | 0.0001 |
| 10 y 30 | 30.542 | 3 | 0.0001 |
| 15 y 20 | 5.5280 | 3 | 0.137 |
| 15 y 25 | 8.3543 | 3 | 0.039 |
| 15 y 30 | 18.372 | 3 | 0.0001 |
| 20 y 25 | 0.0083 | 3 | 1.000 |
| 20 y 30 | 2.8868 | 3 | 0.409 |
| 25 y 30 | 3.3351 | 3 | 25 y 30 |
RV: razón de verosimilitud, gl: grados de libertad, P: probabilidad (p < 0.001).
Las curvas para las copas parcial y totalmente simétricas tuvieron un ajuste similar, igual que en el incremento de su diámetro promedio anual, ambas condiciones superaron a aquellos árboles con copa asimétrica (fig. 6, cuadro 5). Los árboles con copas parcialmente iluminadas crecieron más en su curva que los totalmente iluminados y los poco iluminados, lo que también se encontró en sus incrementos (fig. 6, cuadro 5).
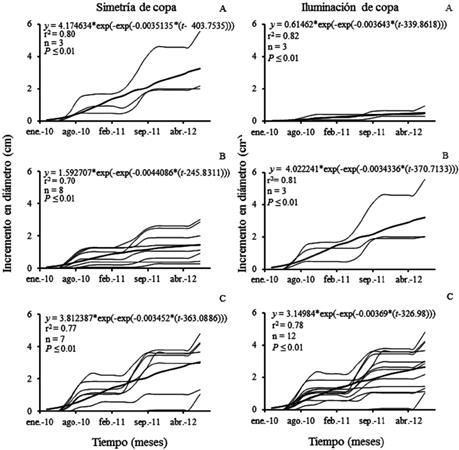
Fig. 6 Crecimiento en diámetro para simetrías: (A) asimétrica, (B) parcialmente simétrica, (C) simétrica e iluminaciones de copa (A) poco iluminada, (B) parcialmente iluminada, (C) totalmente iluminada de Enterolobium cyclocarpum (― datos observados, ▬ ajuste)
Cuadro 5 Crecimiento en diámetro de árboles para las condiciones de copas de Enterolobium cyclocarpum.
| Simetría | Número de árboles | Crecimiento promedio anual (cm) | Desviación Estándar | ||
|---|---|---|---|---|---|
| Mínimo | Medio | Máximo | |||
| 1 | 8 | 0.27 | 0.39 | 0.67 | 0.23 |
| 2 | 7 | 0.34 | 0.69 | 1.32 | 0.48 |
| 3 | 3 | 0.30 | 0.68 | 1.41 | 0.52 |
| Iluminación | |||||
| 1 | 3 | 0.02 | 0.11 | 0.22 | 0.08 |
| 2 | 3 | 0.41 | 0.71 | 1.40 | 0.51 |
| 3 | 12 | 0.34 | 0.62 | 1.16 | 0.42 |
El análisis de verosimilitud mostró que no hubo diferencia significativa entre copas parcialmente simétricas ni totalmente simétricas, como tampoco la hubo entre copas parcialmente iluminadas y totalmente iluminadas (cuadro 6).
Cuadro 6 Prueba de razón de verosimilitud para condiciones de copa de Enterolobium cyclocarpum.
| Condiciones de copa | RV | gl | P | |
|---|---|---|---|---|
| simetría | ||||
| 1 y 2 | 24.874 | 3 | 0.0001 | |
| 1 y 3 | 18.263 | 3 | 0.0001 | |
| 2 y 3 | 0.00001 | 3 | 1.0000 | |
| iluminación | ||||
| 1 y 2 | 40.753 | 3 | 0.0001 | |
| 1 y 3 | 44.291 | 3 | 0.0001 | |
| 2 y 3 | 1.2742 | 3 | 0.735 | |
RV: razón de verosimilitud, gl: grados de libertad, P: probabilidad (p < 0.001).
Periodicidad de anillos de crecimiento
Los anillos de crecimiento fueron visibles, no hubo anillos falsos o discontinuos (fig. 7A), su formación se aprecia entre la primera (marzo-2010) y la última (julio-2012) marca al cambium, con un anillo intermedio que corresponde al transcurso del año 2011 (fig. 7B). Estos anillos están delimitados por una o dos hileras de fibras y células parenquimáticas, las cuales presentan paredes gruesas y diámetros pequeños entre periodos de crecimiento y paredes delgadas y lúmenes amplios durante los periodos de crecimiento. La porosidad es difusa. El parénquima es paratraqueal vasicéntrico de hasta diez células de ancho. (figs. 7C y 7D).

Fig. 7 Periodicidad del crecimiento de Enterolobium cyclocarpum. A. heridas bimestrales en rodaja de 10 cm. B. heridas inicial y final. Anillo correspondiente al año 2011 (flechas).C. corte microscópico de la madera (5x). D. delimitación de anillos por fibras (flechas continuas) y parénquima (flechas discontinuas) (20x). Pv: parénquima vasicéntrico, V: vaso, R: radio (5x).
Discusión
El periodo durante el cual se presentó la caída de hojas en este trabajo (finales de septiembre-fines de enero) es más extenso que lo mencionado por Vázquez-Yanes et al. (1999) para Chamela, Jalisco (febrero-abril) y Guanacaste, Costa Rica (fines de diciembre-mediados de enero) (Borchert, 1994a; 1994b). Una mayor extensión de este evento se relaciona con menores temperaturas y escasez de humedad en el suelo, por lo que la estrategia de la especie es defoliarse durante el invierno (Rojas-Jiménez y Gutiérrez-Soto, 2011) para disminuir su transpiración, evita la pérdida de agua y mantener las reservas de carbohidratos para la consecuente producción de flores y frutos (Pallardy, 2008).
La floración y la fructificación se presentaron en los mismos meses que lo ya estudiado en Chamela, Jalisco y La Mancha, Veracruz (Vázquez-Yanes et al., 1999, Moreno-Casasola et al., 2010). El periodo de brote de hojas fue más extenso (seis a ocho meses) respecto a lo mencionado por Estrada-Castelo et al. (2012) en Cosalá, Sinaloa. De acuerdo con la clasificación de Borchert et al. (2004), Enterolobium cyclocarpum en este trabajo se comporta como una especie invernal por el tiempo en que florece (fines de invierno-principio de primavera), comportamiento que los autores relacionan con un incremento del fotoperiodo y cuya duración puede prolongarse hasta las primeras lluvias (Borchert y Rivera, 2001; Rivera et al., 2002).
La consecuente fructificación de especies invernales como Enterolobium cyclocarpum se caracteriza por un periodo corto de maduración de frutos (3-4 meses aproximadamente) que prepara a las estructuras para su dispersión y germinación para la próxima época de lluvias (Singh y Kushwaha, 2006). El retraso en algunos individuos en su floración y consecuente fructificación explican el desfase de este último evento, adjudicado a sitios con humedad distinta y texturas del suelo diversas (Pineda et al., 2012).
La correlación positiva entre el brote de hojas de Enterolobium cyclocarpum y la temperatura, se ha encontrado en árboles de Costa Rica (Rojas-Jiménez et al., 2007) y Sinaloa, México (Estrada-Castelo et al., 2012). Ambas investigaciones señalan, que esta especie es sensible al ascenso de la temperatura del aire hasta de unos cuantos grados, con lo que se estimula la actividad meristemática y el crecimiento foliar.
La relación significativa entre diámetros mayores, la floración y el brote de hojas se explica por una mayor actividad del cambium vascular debido a la estimulación de fitorreguladores producidos en los brotes (López-Ríos, 2005) y que coincide con los mayores incrementos en individuos de 25 y 30 cm medidos en esta investigación. La nula relación entre la caída de hojas y el diámetro es debida al patrón causal de disminución de la temperatura ya mencionado, el cual se reactiva con el aumento de la misma (Estrada-Castelo et al., 2012). La probable disminución del potencial en frutos en individuos maduros, implica una menor inversión de energía y recursos por árboles de mayor edad, debido a la cantidad de metabolitos requeridos para mantener dicha estructuras (López-Rios, 2005).
El incremento promedio anual medido en el presente trabajo es menor al citado por Makocki et al., (2012) (1.29 cm) en Nayarit, México (precipitación: 1410 mm), aunque es similar a los valores máximos de las categorías 25 y 30. El principal factor que define estas diferencias de crecimiento entre distintas selvas secas con similar precipitación, es el uso que cada especie hace del agua en los diferentes suelos, esto determina los patrones inter e intraanuales en cada caso (Baker et al., 2003; Roig et al., 2005).
Las diferencias encontradas entre categorías diamétricas muestran que a mayores diámetros, mayores incrementos, un hecho mencionado tanto por Baker et al. (2003) como por Adame et al. (2014) en selvas secundarias de Ghana y de Puerto Rico, respectivamente. En ambos estudios, este patrón se relaciona con una mayor copa, mayor captación de luz y producción de fotosintatos necesarios para el engrosamiento del fuste.
Con la respuesta a simetría e iluminación de copa, se comprueba que E. cyclocarpum en Los Varillos, es dependiente de la apertura de claros, en la medida en que requiere para su crecimiento de niveles lumínicos mayores y responde positivamente en crecimiento a la apertura del dosel, por lo que árboles mejor iluminados y simétricos tienen un mayor crecimiento (Ramírez-Angulo et al., 2009).
Las características de los anillos de crecimiento en Enterolobium cyclocarpum de Guerrero coinciden con lo observado en Nayarit, México (Makocki, et al., 2012) con respecto a la periodicidad de formación, delimitación del anillo y ausencia de anillos falsos o discontinuos. Difieren en el tipo de porosidad y cantidad de parénquima vasicéntrico: semianular vs difusa respectivamente, así como mayor cantidad de parénquima vasicéntrico en Enterolobium cyclocarpum de Costa Grande, Guerrero.
Tales semejanzas y diferencias confirman varios hechos: la estacionalidad de la precipitación se relaciona con la formación de anillos de crecimiento anual delimitados por bandas de fibra o parénquima como en otros trabajos en selvas secas (Pineda-Herrera et al., 2012c; Beltrán-Gutiérrez y Valencia-Ramos, 2013).
La porosidad difusa que presenta E. cyclocarpum en Los Varillos se ha descrito en especies donde los vasos inician su formación después que las yemas foliares y culminan cuando el follaje está completamente desarrollado; por tanto, sus dimensiones son similares tanto en la zona de crecimiento inicial como en la terminal (Aloni, 1991). Aun cuando este tipo de porosidad se ha clasificado como deficiente para transportar agua, la especie puede hacer uso de otras estrategias para compensar esta desventaja tales como abundantes reservas (carbohidratos) en células de parénquima que fungen como auxiliares osmóticos en épocas de secas, según se ha observado en E. contortisiliquum (Vell.) Morong (De Lima et al., 2009).
Conclusiones
La caída de hojas de E. cyclocarpum se presentó en meses sin precipitación y con menor temperatura y la formación de brotes se correlacionó con las oscilaciones de la temperatura del aíre en la época de lluvia. La etapa reproductiva se presentó en la época de secas y con las mayores temperaturas. El desarrollo de hojas y flores se ve favorecida en árboles de mayor diámetro. Crece en diámetro en periodos anuales favorecida por la precipitación y sus tasas de crecimiento son mayores en función del tamaño del árbol y mejores simetrías e iluminaciones de su copa.

