











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Los bosques pantanosos en Chile presentan distribución amplia y fragmentada (1 300 km aprox.), que abarca desde Coquimbo (IV región) en el norte, hasta Puerto Montt (X región) en el sur (González et al., 2003). Los árboles dominantes de estos ecosistemas pertenecen, preferentemente, a la familia de las mirtáceas donde se destacan los géneros Blepharocalyx, Luma, Myrceugenia y Tepualia. Hasta el momento se han identificado seis asociaciones florísticas a lo largo de Chile, siendo la conformada por Blepharocalyx cruckshanksii y Myrceugenia exsucca la principal en la región de La Araucanía (Ramírez et al., 1983). En condiciones originales estos bosques se caracterizan por ser siempre verdes, monoestratificados y exhibir un dosel arbóreo cerrado (Ramírez et al., 1995). La fisonomía y la composición florística de estos bosques contrastan con el paisaje y la vegetación dominante del sector en donde se desarrollan, ya que no concuerdan con el clima regional (Ramírez et al., 1983). En consecuencia, los bosques pantanosos corresponden a vegetación azonal, ya que su presencia depende en mayor medida de factores microclimáticos y edáficos (San Martín et al., 1990). Se encuentran asociados a suelos con mal drenaje y dependen del aporte de aguas subterráneas (Villa-Martínez y Villagrán, 1997), lo que les confiere un hidroperiodo que puede ser temporal o permanente (Correa-Araneda et al., 2011) y que es en gran medida responsable del crecimiento y composición de las especies de plantas (Gerritsen y & Greening, 1989; Brooks, 2000; Mitsch y Gosselink, 2007).
Los remanentes de bosques pantanosos que crecen en la depresión central de Chile presentan altos grados de deterioro, lo cual ocurre por acción del hombre al modificar el drenaje de sus suelos con el fin de habilitar terrenos para la agricultura y/o el establecimiento de plantaciones de especies exóticas (San Martín et al., 1988). Además, en muchos de ellos se extrae leña para uso doméstico, lo que puede modificar de manera considerable la estructura vegetacional del bosque (Benfield et al., 2005; Walters, 2005), y poner en peligro la permanencia e integridad del sistema boscoso. Esto puede ser peor si dicha perturbación va dirigida a individuos con un DAP < 10 cm, ya que se corre el riesgo de provocar la extinción local de dichos individuos, al limitar su regeneración natural (Obiri et al., 2002; Lawes et al., 2007).
El aspecto más estudiado de estos sistemas boscosos ha sido el florístico-vegetacional (Ramírez et al., 1983, 1995; San Martín et al., 1988, 1990, 1992, 2006; Hauenstein et al., 2014). En tanto que antecedentes sobre su composición estructural, en general, son muy escasos y, en el caso particular de los bosques pantanosos de La Araucanía, son inexistentes.
El escenario actual de los bosques pantanosos no augura un buen futuro, su escaso conocimiento y su nulo resguardo en sistemas de protección efectivos están limitando su conservación, por lo tanto, cualquier nueva información respecto de ellos es valiosa. De este modo, el objetivo del presente studio es caracterizar la composición estructural vertical y horizontal de remanentes de bosque pantanoso de la depresión central de la región de La Araucanía en Chile.
Material y métodos
Área de estudio
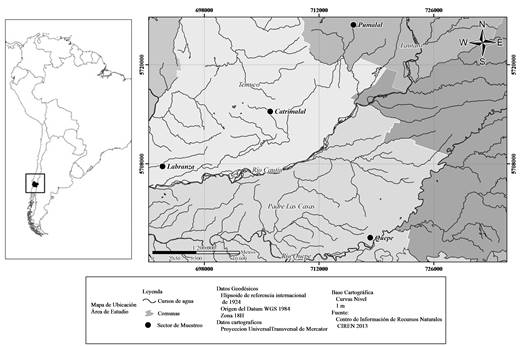
El área de estudio se emplaza en la depresión central de la región de La Araucanía, provincia de Cautín, Chile. Específicamente los remanentes evaluados prosperan en las localidades de Catrimalal, Labranza, Pumalal y Quepe (fig. 1). Para cada uno de ellos se indican las coordenadas, superficie, altitud e hidroperíodo (cuadro 1). Este último dato fue obtenido de Correa-Araneda et al. (2012) y complementado mediante observación tras varias prospecciones a las localidades. El clima del área de estudio pertenece al tipo mediterráneo perhúmedo, en el cual las influencias marítimas están parcialmente determinadas por el cordón longitudinal de la Cordillera de la Costa (Di Castri y Hajek, 1976). Los suelos, corresponden a “trumaos aluviales” y se caracterizan por ser planos a ligeramente ondulados, aluviales, estratificados, muy variables en profundidad (Peralta, 1976).
Cuadro 1 Coordenadas, altitud, superficie e hidroperíodo de las localidades estudiadas.
| Localidad | Coordenadas UTM | Altitud (m s.n.m.) | Superficie (hectáreas) | Hidroperíodo(meses) | |
|---|---|---|---|---|---|
| Este | Norte | ||||
| Catrimalal | 0706031 | 5714338 | 111 | 12.3 | 6 |
| Labranza | 0692903 | 5707674 | 59 | 11.6 | 6 |
| Pumalal | 0716194 | 5724835 | 158 | 13.8 | 6 |
| Quepe | 0718247 | 5699040 | 170 | 10.3 | 2 |
Muestreo y análisis de datos
La caracterización de la estructura de la vegetación se realizó mediante el levantamiento de parcelas de 300 m2 considerando una por localidad, con lo cual se superó ampliamente el área mínima establecida para este tipo de bosques por Ramírez et al. (1983).En cada parcela se registraron las siguientes variables: Identidad de la especie vegetal, diámetro a la altura del pecho (DAP > 5 cm), proyección de la copa en la dirección de los cuatro puntos cardinales, altura total y posición de los individuos dentro de la parcela (Steubing et al., 2002). Los datos fueron procesados en el software Stand Visualization System (SVS) versión 3.36, que permitió proyectar una imagen tridimensional de la estructura horizontal de los rodales. Además, se registró el componente florístico en el interior de cada parcela, lo cual se detalla en un cuadro de presencia de especies. Cuando se habla de estructura vegetacional es necesario distinguir tres componentes, la estructura cuantitativa, que expresa la abundancia de especies en una comunidad; la estructura vertical que indica el ordenamiento de la vegetación en capas o estratos; y la estructura horizontal, la cual hace referencia a la distribución espacial de los individuos en el plano horizontal o superficie del rodal (Donoso, 1993).
Para determinar posibles diferencias entre sitios y previa comprobación de normalidad con la prueba de Kolmogórov-Smirnov, se realizó un ANCOVA, utilizando el DAP como variable dependiente y la altura como variable independiente. Posteriormente, se hizo una prueba de Tukey, para determinar diferencias significativas entre las medias (Sokal y Rohlf, 1981). Además, se confeccionó un dendrograma de similitud con la abundancia de las especies de cada localidad, basado en el índice de Bray-Curtis (Bray y Curtis, 1957). Todos los análisis anteriores fueron hechos con el software XLSTAT 7.52.
Resultados
Catrimalal
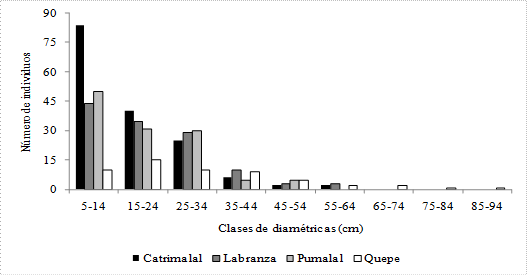
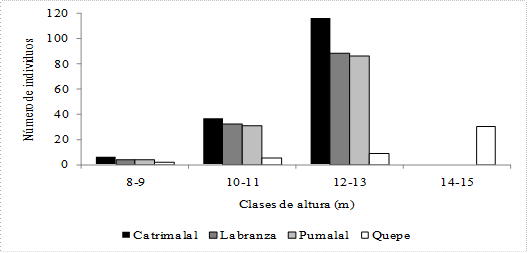
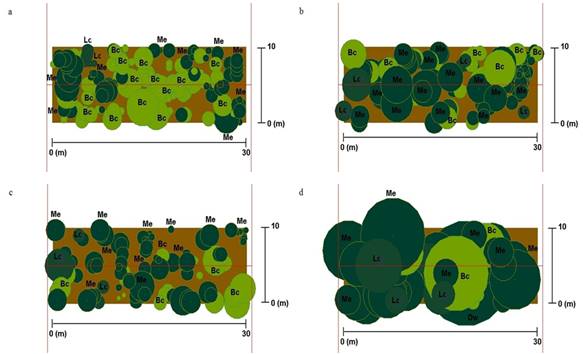
En esta localidad la densidad arbórea fue de 5 300 individuos/ha, el perfil vertical está conformado por tres especies, siendo la más abundante B. cruckshanksii con 2 733 individuos, M. exsucca con 2367, y Luma chequen con 200. La distribución de los árboles por clase diamétrica (fig. 2) se ajusta a una estructura de bosque multietáneo, el DAP promedio alcanzado por los árboles fue de 17 cm (5-62; ± 10,8), en tanto que la altura (fig. 3) fue de 11.5 m (9-12; ± 0.8). La cobertura de copa promedio (fig. 4) por especie fue de 0.91 m2 (± 0.04) para M. exsucca; 0.90 m2 (± 0.04) para B. cruckshanksii y 0.75 m2 (± 0.11) para L. chequen. La cobertura del dosel fue de 81% (fig. 5a).
Fuente: Autores, 2017
Fig. 2 Número de individuos por clase diamétrica para las localidades estudiadas.
Fuente: Autores, 2017
Fig. 3 Número de individuos por clase de altura para las localidades estudiadas.

Labranza
En esta localidad la densidad arbórea fue de 4 133 individuos/ha, el perfil vertical está compuesto por tres especies, con un dominio considerable de M. exsucca con 2 733 individuos, le siguen B. cruckshanksii con 967 y L. chequen con 433. La distribución de los árboles por clase diamétrica (fig. 2) se ajusta a una estructura de bosque multietáneo, el DAP promedio alcanzado por los árboles fue de 21.4 cm (6-58; ± 11.6), en tanto que la altura (fig. 3) alcanzó los 11.5 m (8-13; ± 0.89). La cobertura de copa promedio (fig. 4) por especie fue de 1.34 m2 (± 0.08) para B. cruckshanksii; 1.34 m2 (± 0.11) para L. chequen y 1.12 m2 (± 0.05) para M. exsucca. La cobertura del dosel fue de 89% (fig. 5b).
Pumalal
En esta localidad la densidad arbórea fue de 4 033 individuos/ha, el perfil vertical está conformado por tres especies, con un dominio considerable de M. exsucca con 3 033 individuos, le sigue B. cruckshanksii con 833 y L. chequen con 167. La distribución de los árboles por clase diamétrica (fig. 2) se ajusta a una estructura de bosque multietáneo, el DAP promedio alcanzado por los árboles fue de 19.2 cm (5-54; ± 11.6), en tanto que la altura (fig. 3) promedio fue de 11.5 m (9-12; ± 0.82). La cobertura de copa promedio (fig. 4) por especie fue de 1.3 m2 (± 0.3) para L. chequen; 1.03 m2 (± 0.1) para B. cruckshanksii y 0.9 m2 (± 0.04) para M. exsucca. La cobertura del dosel fue de 75% (fig. 5c).
Quepe
En esta localidad la densidad arbórea fue de 1 833 individuos/ha, el perfil vertical está compuesto por cuatro especies, siendo la más abundante M. exsucca con 1 033 individuos, le sigue L. chequen con 400, B. cruckshanksii con 300 y D. winteri con 100. La distribución de los árboles por clase diamétrica (fig. 2) se ajusta a una estructura de bosque multietáneo, el DAP promedio alcanzado por los árboles fue de 31.3 cm (8-87; ± 18.5), en tanto que la altura (fig. 3) fue de 14 m (8-15; ± 1.4). La cobertura de copa promedio (fig. 4) por especie fue de 4 m2 (± 1.6) para D. winteri; 2.41 m2 (± 0.16) para M. exsucca; 2.25 m2 (± 0.24) para L. chequen y 2.22 m2 (± 0.46) para B. cruckshanksii. La cobertura del dosel fue de 98% (fig. 5d).
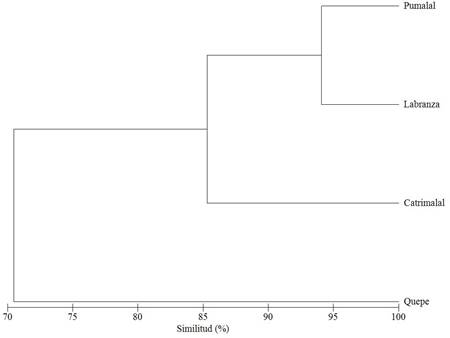
El ANCOVA indica la existencia de diferencias significativas entre las cuatro localidades estudiadas (F4 = 72,885; p < 0.0001), las que de acuerdo con la prueba de Tukey (cuadro 2) se dan entre Labranza y Catrimalal; y Quepe con todas las demás localidades. Esto es reforzado por el dendrograma de similitud (fig. 6) que muestra a Quepe distanciado del grupo conformado por Pumalal, Labranza y Catrimalal con más del 85% de semejanza entre sí.
Cuadro 2 Valores de la prueba de Tukey para las localidades estudiadas.
| Categorías | Dif | Diferencia estandarizada | Valor crítico | Pr. > Dif | Sig. |
|---|---|---|---|---|---|
| Quepe - Catrimalal | 14 064 | 8 731 | 2 579 | < 0.0001 | Sí |
| Quepe - Pumalal | 11 879 | 7 094 | 2 579 | < 0.0001 | Sí |
| Quepe - Labranza | 10 017 | 6 027 | 2 579 | < 0.0001 | Sí |
| Labranza - Catrimalal | 4 047 | 3 303 | 2 579 | 0.006 | Sí |
| Labranza - Pumalal | 1 862 | 1 424 | 2 579 | 0.485 | No |
| Pumalal - Catrimalal | 2 185 | 1 759 | 2 579 | 0.295 | No |
El componente florístico en el interior de las parcelas está constituido por 24 especies de plantas vasculares (apéndice 1), de las cuales sólo una es introducida (Rubus constrictus P.J. Müll. & Lefèvre) y fue registrada en Pumalal. La localidad que presenta la mayor riqueza florística es Quepe con 19 especies, le siguen Catrimalal y Pumalal con 10; y finalmente Labranza con nueve.
Discusión
Los bosques pantanosos de mirtáceas de la depresión central de la región de La Araucanía en el sur de Chile se presentan claramente como comunidades azonales, cuya conformación estructural está restringida al dominio de unas pocas especies adaptadas a condiciones extremas (inundaciones periódicas). Si bien la asociación boscosa dominante en esta latitud del país posee una distribución amplia, la misma se hace cada vez más fragmentada debido a la acción antrópica siempre presente.
La estructura horizontal de los bosques presenta una cobertura promedio del 86%, valor cercano al documentado por Hauenstein et al. (2005) para comunidades pantanosas costeras de la región de La Araucanía, pero que se aleja un poco de la cobertura absoluta del dosel arbóreo descrita en condiciones originales (Ramírez et al., 1995). En este sentido, Pumalal es la localidad con la menor cobertura de dosel (75%), situación que hace referencia a importantes perturbaciones en el sistema boscoso. Lo anterior concuerda muy bien con la presencia de la única especie introducida, en relación a esto, algunos autores han planteado que bosques con mayor apertura de dosel probablemente contengan especies exóticas (Meekins y McCarthy, 2001; Meiners et al., 2002).
La estructura vertical está compuesta por cuatro especies: M. exsucca, B. cruckshanksii, D. winteri y L. chequen, las cuales son comunes a todos los bosques pantanosos de Chile (Ramírez et al., 1995). Las tres primeras son consideradas muy abundantes, en tanto que L. chequen por ser menos habitual se le ha calificado de frecuente (Ramírez et al., 1995). Las clases diamétricas obtenidas para las cuatro localidades se presentan en forma de J invertida, donde los árboles están distribuidos en varias categorías, situación similar a lo reportado para bosques pantanosos de la región de Aysén (Promis et al., 2013). Los valores promedio de altura y DAP (11.8 m y 22.1 cm, respectivamente) registrados en las cuatro localidades estudiadas están dentro de los rangos informados para bosques pantanosos de Chile central (San Martín et al., 1988). Sin embargo, la altura promedio registrada en el presente estudio estaría bajo los 20 m documentados por Ramírez et al. (1995). Lo anterior puede tener su explicación en el impacto antrópico en las cuatro localidades estudiadas, lo que se refleja en la extracción de leña para uso doméstico (Ramírez et al., 1995). Se encontró que una mayor abundancia de individuos arbóreos (e.g. Catrimalal, Labranza y Pumalal) está en directa relación con diámetros más pequeños (Jaña-Padro et al., 2006), lo contrario ocurre en Quepe, donde una menor densidad de árboles (1 833 ind/ha) está relacionado con diámetros más grandes. La dominancia de mirtáceas en general y de M. exsucca en particular, sugiere en primer lugar, la resistencia a la inundación de los representantes de dicha familia (Latsague et al., 2010); y en segundo lugar plantea la idea de que M. exsucca es la especie mejor adaptada a soportar las condiciones extremas propias de estos ambientes (Ramírez et al., 1983), ya que presenta mayor resistencia constitucional y plasmática a la sequia que otros miembros de la familia (Weinberger et al., 1973).
De acuerdo con la prueba ANCOVA, la única localidad significativamente diferente a todas las demás fue Quepe. Ésta es dada principalmente por la conformación de especies en términos estructurales, donde D. winteri juega un rol fundamental ya que, primeramente, su presencia es exclusiva de esta localidad y, en segundo lugar, contribuye al registro de los mayores valores de DAP (87 cm), altura (15 m) y coberturas de copa (4 m2). Las demás localidades presentan valores de DAP y altura similares, siendo siempre más bajos que los registrados en Quepe. Dicha situación estaría condicionada por el régimen de inundación del bosque, ya que hidroperiodos más largos provocan no sólo menor riqueza de especies, sino también efectos fisiológicos como menor altura (Correa-Araneda et al., 2012).
En cuanto al componente florístico en el interior de las parcelas, el bajo número de especies registradas obedece al anegamiento estacional que experimentan las cuatro localidades, situación que los convierte en biotopos extremos donde la proliferación del sotobosque es muy poco probable (Ramírez et al., 1995). A pesar de lo anterior, la presencia de R. constrictus en Pumalal, sugiere por un lado el potencial invasor de dicha especie (Fuentes et al., 2014) y por otra parte, resalta la constante amenaza que representan las matrices aledañas (e.g. agrícola) como fuentes de ingreso de plantas exóticas (Charbonneau y Fahrig, 2004).
Conclusiones
Los resultados reportados son de especial interés, ya que generan información desconocida sobre la conformación estructural de los bosques pantanosos de la región de La Araucanía. Si bien este sistema boscoso es de singular importancia, también ha sido altamente perturbado por la acción del hombre. Lo anterior se refleja en los escasos niveles de cobertura del dosel arbóreo que fueron registrados y se confirma también con las bajas alturas encontradas en las localidades de Catrimalal, Labranza y Pumalal. El hidroperiodo parecer ser un factor clave en el desarrollo de este tipo de bosques, lo que queda claramente demostrado en las diferencias mostradas por Quepe, las cuales se deben principalmente a la presencia de Drimys winteri, especie arbórea que registra las mayores alturas, DAP y coberturas de copa.
Estos sistemas boscosos están bajo constante presión antrópica, por lo que tal y como ocurre en bosques pantanosos de otras partes del mundo (e.g. manglares), es muy probable que tanto la estructura como la composición florística estén influenciadas por las actividades que de manera directa y/o indirecta se realizan en ellos (Walters, 2005; Berger et al., 2006).

