










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPolibotánica
versión impresa ISSN 1405-2768
Polibotánica no.38 México ago. 2014
Patrones de nerviación foliar en Myrtaceae de la Estación Biológica La Selva, provincia de Heredia, Costa Rica
Leaf venation patterns in myrtaceae of la selva biological station, Heredia, Costa Rica
Gabriela Jolochin
Laboratorio de Botánica, Departamento de Biología Vegetal y Laboratorio de Dendrología, Departamento de Producción Forestal y Tecnología de la Madera. Facultad de Agronomía, Universidad de la República, Av. Eugenio Garzón 780, 12900, Montevideo, Uruguay. Correo electrónico: gjolochin@gmail.com.
Recibido: 14 agosto 2012.
Aceptado: 8 noviembre 2013.
Resumen
Se realizaron descripciones de la arquitectura foliar, con énfasis en patrones de nerviación, de cuatro géneros y 14 especies de Myrtaceae incluyendo dos especies citadas como exóticas, para la Estación Biológica La Selva, OET, Heredia, Costa Rica. El material fresco descrito fue coleccionado en la estación biológica y fueron visitados los herbarios INB y CR, además de consultar el herbario de la propia estación e imágenes del MO. El 85% de las especies presentan un patrón de nerviación secundaria camptódromo-broquidódromo y ca. 50% poseen nervaduras secundarias rectas, que sólo se curvan cercanas al margen simulando una nervadura intramarginal. Las nervaduras terciarias son detectables a simple vista sólo en siete de las especies, de las cuales cuatro poseen un patrón reticulado y la nerviación última marginal en arcos o no visible. La combinación de los datos de arquitectura foliar con patrones de nerviación permitió discriminar las especies y obtener una clave de determinación de fácil uso en el campo.
Palabras clave: Costa Rica, Estación Biológica La Selva, morfología foliar, Myrtaceae, patrones de nerviación.
Abstract
Descriptions base in leaf architecture, with emphasis in venation patterns, to four genera and 14 species found in the zone, including two species mentioned as exotic, for the Biological Station La Selva, OET, Heredia, Costa Rica. Fresh leaves were collected in the biological station and INB and CR herbaria were visited, besides to consult the herbarium of the own station and images of the MO. It was observed that 85% of the species have a secondary venation pattern camptodromus-brochidodromus and ca. 50% have straight secondary ribbings that only curve near the margin simulating intra-marginal ribbing. The third venation pattern was detectable only in seven species, four with a reticulate pattern and last marginal venation with arches or no visible. The combination of leaf architecture data with venation patterns allow discriminating species and generates a key of determination for easy field use.
Key words: Costa Rica, La Selva Biological Station, leaf morphology, Myrtaceae , venation patterns.
Introducción
La familia Myrtaceae posee alrededor de 144 géneros, con 3 000-3 800 especies, de amplia distribución en América, Asia tropical y Australia, donde son particularmente diversas, con sólo dos géneros de pocas especies en el Sur de Europa y en el Norte y Este de África (Zomlefer, 1994; Judd et al., 1999; Wilson et al., 2001; Watson & Dallwitz, 2007). Esta familia tradicionalmente comprende dos subfamilias, divididas principalmente con base en el tipo de fruto; cápsula o fruto seco indehiscente en Leptospermoideae y carnoso como baya o drupa Myrtoideae (Zomlefer, 1994). La primera de distribución principalmente en Australia y la segunda en América del Sur y Central (Johnson & Briggs, 1984). Myrtoideae presenta una tribu, Myrteae, representada en América así como en Australasia con su mayor diversidad en las regiones tropicales de América del Sur y Central. Costa Rica posee una mirtiflora con 17 géneros y 99 especies de los cuales 12 y 86 son nativos, respectivamente (Barrie et al., 2007). La Estación Biológica La Selva de la Organización para Estudios Tropicales (OET) está ubicada al noreste de Costa Rica (América Central) sobre la vertiente caribeña donde se encuentra la mayor diversidad de especies de Myrtaceae en los bosques tropicales húmedos (Barrie et al., 2007). A pesar de su importancia tanto económica como maderable, frutal, y ecológica (Zomlefer, 1994), los estudios taxonómicos de la familia que se han realizado aún no han resuelto muchas de las relaciones entre sus géneros, y sus especies son de difícil identificación y delimitación, por lo que varían considerablemente con las diferentes clasificaciones. La gran diversidad de Myrteae ha dado lugar a que se generaran subtribus basadas principalmente en caracteres relacionados a la morfología de semillas y embriones, lo que hace más complejas las delimitaciones en ausencia de frutos en las muestras coleccionadas (Mc Vaugh, 1968; Zomlefer, 1994).
Los patrones de nerviación foliar por lo general no son tomados en cuenta en la mayoría de los estudios taxonómicos, aunque se han realizado algunos trabajos en paleo-botánica (Anzótegui, 2000) y en los últimos años Romero & Dibbern (1982), Mantese & Montaldo (2002), Cardoso & Sajo (2004, 2006), Jolochin (2008) y González (2011), han realizado estudios morfoanatómicos para Myrtaceae, los que han demostrado la utilidad de los patrones de nerviación foliar en la determinación de material estéril.
Los órdenes de las nerviaciones se diferencian en clases de acuerdo al calibre de las venas, donde cada orden desarrolla cierto grado de uniformidad en su modelo de distribución en relación a las demás y a las características marginales de la hoja, aunque en la práctica la diferenciación de los órdenes es más compleja que observar el calibre, curso y modelo de nerviación. El reconocimiento de estos órdenes son parte fundamental para la descripción de la arquitectura foliar. Las nervaduras primarias son las de mayor calibre y aparecen solitarias, emergen del pecíolo de la hoja y su recorrido es derecho en Myrtaceae. Las nervaduras secundarias son de menor calibre, se originan de las primarias y sus ramificaciones son de igual calibre, medidas justo por encima de su punto de ramificación, siendo su recorrido derecho o curvado (uniformes: con arco suave o aumentando gradualmente su radio de curvatura; abrupto: aumento local brusco en el radio de curvatura), hasta ramificado (Hickey, 1974). El ángulo de divergencia, medido en la axila formada por la nervadura primaria y la secundaria, el cual generalmente varía entre moderado (45-65°) y ancho (65-80°) en Myrtaceae, aunque dentro de la misma hoja según se encuentren en las bases o hacia los ápices de las hojas pueden presentar variaciones (Cardoso & Sajo, 2006). Cuando las nervaduras secundarias forman arcos, éstos se pueden unir a las secundarias adyacentes en ángulo agudo, recto u obtuso, a veces formando arcos secundarios y hasta una nervadura intramarginal, probablemente como resultado de la fusión y enderezamiento de segmentos de arcos broquidódromos secundarios exmediales, aparentando ser una vena independiente paralela a los márgenes de la hoja. En el espacio entre las secundarias pueden aparecer las nervaduras intersecundarias, de calibre intermedio entre las secundarias y terciarias, originadas en la primaria y esparcidas entre las secundarias (generalmente de patrón de nerviación exmedial), de trayectoria paralela o casi. Pueden ser simples o compuestas cuando están formadas por segmentos de venas terciarias (de calibre menor a las secundarias) coalescentes en más del 50% de su longitud y formadas por fusión de las terciarias derivadas de los arcos, generalmente admediales (Hickey, 1974; Cardoso & Sajo, 2004; 2006).
Un gran porcentaje de las muestras coleccionadas en bosques tropicales húmedos sólo constan de material estéril debido a la dificultad de obtener material fértil de ramas de los árboles del dosel alto del bosque, existiendo una gran necesidad de descripciones de caracteres morfológicos vegetativos más detallados.
El objetivo de este trabajo es describir los patrones de nerviación de especies de Myrtaceae citadas para la Estación Biológica La Selva y generar una clave de determinación combinando dichos patrones con caracteres morfológicos foliares utilizados normalmente en las descripciones. Claves basadas en este tipo de descripciones permitirán identificar especies en ausencia de estructuras reproductivas siendo una buena herramienta tanto de campo para ser utilizada en la Estación Biológica de La Selva, como para determinaciones de materiales de herbario.
Material y métodos
La colección de materiales para la determinación de especies se realizó en la Estación Biológica La Selva de la Organización para Estudios Tropicales (OET), ubicada en la Provincia de Heredia (84° 00'12.9''W; 10° 25'52.6''N), en la vertiente caribeña, al norte de Costa Rica (fig. 1).

Se citan seis géneros y 18 especies, 13 nativas y cinco especies exóticas cultivadas en La Estación Biológica La Selva (La Selva Flórula Digital | http://sura.ots.ac.cr/local/florula4/; Barrie, 2005; Barrie et al., 2007): Eucalyptus globulus Labill., Eugenia apulcensis Steud., Eugenia basilaris Mc Vaugh, E. corusca Barrie, E. glandulosopunctata P. E. Sánchez & Poveda, E. hammelii Barrie, E. hartshornii Barrie, E. lithosperma Barrie, E. magniflora Barrie, E. sarapiquensis P. E. Sánchez, E. selvana Barrie, E. stipitata Mc Vaugh, E. uniflora L., Myrcia aliena Mc Vaugh, M. splendens (Sw.) DC., Myrciaria floribunda (West ex Willd.) O. Berg, Myrciaria vexator Mc Vaugh, Psidium friedrichsthalianum (O. Berg) Nied., P. guajava L. y Syzigium malaccense (L.) Merr. & L. M. Perry. Se excluyen de este trabajo E. globulus, E. apulcensis, M. aliena, M. vexator y S. malaccense, ya que no se accedió a material fresco ni a especímenes de herbario coleccionados en el área de estudio.
La revisión de los especímenes se realizó en los Herbarios de La Selva (OTS LS), del Instituto Nacional de Biodiversidad (INB) y del Museo Nacional (CR) y la consulta de imágenes disponibles en TROPICOS ® (http://www.tropicos.org/) del Missouri Botanical Garden (MO), para completar el estudio. Las determinaciones se cotejaron con la bibliografía sobre Myrtaceae en el Manual de la Flora de Costa Rica (2007) y Barrie (2005), ya que siete de las especies presentes en La Selva son nuevas combinaciones.
Las descripciones foliares con énfasis en los patrones de nerviación se realizaron incluyendo caracteres de fácil reconocimiento en muestras vegetativas que puedan observarse a campo, tomándose medidas y descripciones morfológicas de 3-4 hojas por espécimen. Se describen 14 especies las cuales fueron coleccionadas y revisadas en los herbarios ya mencionados, excluyendo a Eugenia hammelii la cual no fue incluida ya que sólo se accedió a un espécimen identificado como afín a la especie y no se obtuvo material fresco de la zona de estudio.
Los tipos de arquitectura foliar están basados en Hickey (1974) y Ash et al. (1999), con énfasis en los patrones de nerviación tanto en las descripciones como en la clave, observables a simple vista o con lupa 10x.
Resultados
Descripciones taxonómicas
1. Eugenia basilaris Mc Vaugh, Fieldiana, Bot., 29: 464. 1963.
Material examinado: COSTA RICA. Gerardo Herrera 1106 (CR); L.J. Poveda 1208 (CR); R. Espinosa 1738 (INB); Moraga & Calixto 733 (INB); Petrona Ríos 313 (INB); Moraga & Calixto 733 (MO); Orlando Vargas 277 (MO). NATIVA.
Hojas ovadas o elípticas, de 6-12 cm x 3-5 cm, cartáceas; patrón de nerviación secundaria camptódromo broquidódromo, vena media surcada adaxialmente, poco prominente en la cara abaxial; venas secundarias de 6-8 pares, algo prominentes en el envés, formando fuertes arcos con las secundarias adyacentes de 3-5 mm del margen; venas terciarias reticuladas; nerviación última marginal en arcos completos; pubescentes en el envés, con tricomas escamosos; puntos glandulosos esparcidos en la base; base redondeada hasta cordada; márgenes enteros, ápice agudo hasta un poco acuminado; peciolos cortos de ca. 3 mm, terete, hirsuto al igual que las ramillas jóvenes (fig. 2 H).

2. Eugenia corusca Barrie, Novon 15: 14. 2005.
Material examinado: COSTA RICA. Sin colector 171218 (CR); Gary S. Hartshorn 1315 (CR); Celso Alvarado 131 (INB); B. Boyle et al. 5280 (INB); Ricardo Rueda et al. 5228 (MO); Sin colector 11693 (OET LS). NATIVA.
Hojas elípticas obovadas, de 7-15 x 2.5-5 cm, cartáceas; nerviación camptódroma broquidódroma, vena media algo surcada en el haz, prominente en el envés; nervaduras secundarias 5-9 pares, curvadas desde la base, disminuyendo su calibre cerca del margen e interconectada con numerosos arcos de terciarias de la nerviación última marginal, arcos de 2-5 mm del margen; nerviación terciaria alterna percurrente; nerviación última marginal en arcos conspicuos; glabras adaxialmente, abaxialmente cubiertas por tricomas simples, diminutos, ócreos o hialinos los cuales producen un reflejo argenteo típico; puntos glandulosos numerosos; base cuneada hasta angostamente cuneada; márgenes decurrentes por el peciolo; ápice acuminado o caudado acuminado; peciolo de ca. 5 mm, algo surcado adaxialmente, generalmente glabros (fig. 2G).
3. Eugenia glandulosopunctata P.E. Sánchez et Poveda, Brenesia, 24: 413. 1985.
Material examinado: COSTA RICA. Reinaldo Aguilar 1359 (CR); L.D. Gómez et al. s/n (CR 107591); Karsten Thomsen 86 (CR); B. Boyle et al. 64 (INB); B. Boyle 190 (INB); Enia Navarro V. 716 (INB); Darío Pereyra 445 (MO); Elías Rojas 215 (MO). NATIVA.
Hojas de 5-8 x 2-3.5 cm, elípticas, cartáceas; nerviación camptódroma broquidódroma, vena media surcada adaxialmente, poco prominente abaxialmente; venas secundarias 12-18 prominentes en ambas caras, con venas intrasecundarias abundantes; arcos broquidódromos abtusos simulando una vena intramarginal de similar a 0.5-1 mm del margen; glabras; numerosas glándulas convexas conspicuas en ambas caras; base cuneada hasta aguda; margen decurrente por el pecíolo; ápice largamente acuminado, con el acumen de 10-20 mm; pecíolos 2-3 mm, surcado adaxialmente (fig. 2E)
4. Eugenia hartshornii Barrie, Novon 15: 24. 2005.
Material examinado: COSTA RICA. I. A. Chacón 152 (CR); Karsten Thomsen 1017 (CR); Nelson Zamora 2527 (INB); G. N. Frankie 1886 (MO). NATIVA.
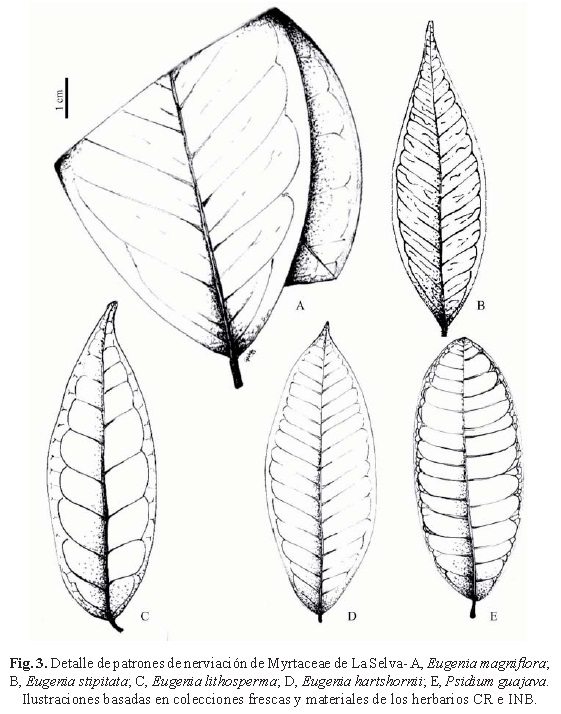
Hojas de 13-20 x 6-8 cm, elípticas u obovadas, cartáceas, discoloras; nerviación camptódroma broquidódroma, vena media prominente en el envés; venas laterales de 16-18 pares, con venas intersecundarias prominentes en la cara abaxial; cara adaxial glabra o pilosa con algunos pelos escamosos, persistentes y más densos en la vena media, la cara abaxial densa y uniformemente cubierta por pubescencia de pelos ócreos e infundibuliformes; glándulas numerosas, diminutas, convexas y salientes en el haz, en el envés mayores, típicamente oscuras; base cuneada; margen entero, algo revoluto; ápice acuminado; pecíolos 5-6 mm, surcados adaxialmente, pubescentes (fig. 3D).
5. Eugenia lithosperma Barrie, Novon, 15: 30. 2005.
Material examinado: COSTA RICA. L. J. Poveda 185 (CR); Ingrid Wattenberg 59 (CR); E. Bello 531 (INB); G. N. Frankie 78a (MO). NATIVA.
Hojas de 7-14 x 3-6 cm, ovadas a elípticas, cartáceas, concoloras; nerviación camptódroma broquidódroma, vena media prominente abaxialmente; venas laterales 8-14 pares, poco marcadas en la cara adaxial, prominentes en la cara abaxial y a menudo coloreadas; haz glabro o más comúnmente con pilosidad escasa, con tricomas escamosos y adpresos, con la vena media glabra o con pubescencia persistente, envés y vena media cubiertas por tricomas adpresos y persistentes, volviéndose más escasos en hojas adultas; glándulas numerosas, pequeñas, en ambas caras, usualmente oscuras abaxialmente; base cuneada; ápice acuminado hasta caudado-acuminado; pecíolos 8-12 mm, surcados adaxialmente, con tricomas escamosos adpresos (fig. 3C).
6. Eugenia magniflora Barrie, Novon, 15: 32. 2005
Material examinado: COSTA RICA. Michael Grayum et al. 5068 (CR). NATIVA.
Hojas de 20-35 x 9-16 cm, obovadas o elípticas, cartáceas, discoloras; nerviación camptódroma broquidódroma, vena media prominente en la cara abaxial; venas laterales 13-15 pares, planas adaxialmente, prominentes en la cara abaxial; ambas caras glabras o escasamente pubérulas, la vena media con pilosidad persistente en ambas caras; glándulas algo numerosas, pequeñas, cóncavas en la cara abaxial, algo oscuras; base cuneada; ápice abruptamente acuminado; pecíolos 10-15 mm, teretes o algo planos adaxialmente, pubescentes (fig. 3A).
7. Eugenia sarapiquensis P.E. Sánchez, Brenesia, 25: 317. 1986.
Material examinado: COSTA RICA. P. E. Sánchez V. & N. Zamora 1239 (CR); Evelio Alfaro 2120 (INB); Nelson Zamora & A. Zeledón 2365 (INB); M. Grayum 2830 (OET LS). NATIVA.
Hojas de 10-21 x 4-10 cm, elípticas a obovadas, cartáceas, concoloras; nerviación camptódroma broquidódroma, venas laterales 5-8 pares, planas en la cara adaxial, prominentes en la cara abaxial, con arcos fuertemente curvados ascendentes uniéndose a la secundaria adyacente con un ángulo agudo, basales de secundarias disueltos en arcos sucesivos fusionados al resto de secundarias; nerviación terciaria conspicua reticulada; haz algo híspido, con tricomas de 0.1 mm, inconspicuos en hojas adultas, pero persistentes en la vena media, envés algo híspido, más denso a lo largo de la vena media y las laterales; glándulas numerosas, pequeñas en ambas caras; base cuneada, decurrente por el pecíolo; ápice acuminado, con el extremo redondeado; pecíolos 8-10 mm, surcados adaxialmente, algo híspidos (fig. 2I).
8. Eugenia selvana Barrie, Novon, 15: 42. 2005.
Material examinado: COSTA RICA. L. J. Poveda 1153 (CR); Barry Hammel 9184 (MO); Barry Hammel 9184 (OET LS); Barry Hammel 11107 (OET LS). NATIVA.
Hojas de 4-9 x 1.5-3 cm, elípticas a ovadas, cartáceas, discoloras; nerviación camptódroma broquidódroma, vena media prominente en la cara abaxial, venas laterales seis o siete pares, con pocas venas intersecundarias, arcos secundarias curvados fuertemente en forma ascendente, uniéndose a las adyacentes con un ángulo agudo o casi, prominentes en la cara abaxial; haz con pilosidad hirsuta persistente a lo largo de la vena media, envés con tricomas escamosos a lo largo de la vena media y las laterales; base cuneada; márgenes enteros, ciliado, a veces solamente en la mitad inferior; glándulas inconspicuas en el haz, numerosas, pequeñas y oscuras en el envés; ápice acuminado hasta caudado acuminado; pecíolos 1-3 mm, teretes o cóncavos adaxialmente, pubescencia híspida persistente (fig. 2C).
9. Eugenia stipitata Mc Vaugh, Fieldiana, Botany 29(3): 219. 1956.
Material examinado: COSTA RICA. Álvaro Fernández 1413 (INB); Ronald Liesner & Germán Carnevali 23000 (MO). EXÓTICA.
Hojas de 7-12 x 3-6 cm, elípticas hasta oblongas, cartáceas, notablemente discoloras; nerviación camptódroma eucamptódroma, vena media prominente en la cara abaxial, venas secundarias ocho pares, irregularmente espaciadas, arcos de secundarias curvadas desde la base; nerviación terciaria alterna percurrente; pilosidad en el envés, el cual es notablemente blanquecino ceroso; glándulas en ambas caras poco conspicuas; base aguda; margen entero poco revoluto; ápice agudo; pecíolos 8-12 mm, teretes o algo surcados en la cara adaxial (fig. 3 B).
10. Eugenia uniflora L., Species Plantarum 1: 470-471. 1753. Stenocalyx uniflorus (L.) Kausel, Lilloa 32: 331. 1967. Tipo: ?. Hábitat in India, Native to southern South America, widely cultivated. Sin fecha, Micheli, Nov. Pl. Gen. 226, pl. 108 (1729) (Lectotipo: F). LT designado por McVaugh, Taxon 5: 140 (1956).
Material examinado: COSTA RICA. William A. Haber & Willow Zuchowski 10072 (CR); Robert K. Godfrey 66270 (MO). URUGUAY. M. Berro 2311 (MVFA); M. Bonifacino s/n (MVFA 25040). EXÓTICA.
Hojas de 3-6 x 1.5-3 cm, ovadas hasta elípticas, cartáceas, notablemente discoloras; nerviación camptódroma broquidódroma, vena media poco marcada en ambas caras; venas laterales 5-7 pares, con arcos prominentes uniéndose con las secundarias adyacentes con ángulos agudos muy conspicuos simulando una vena intramarginal a 3-5 mm del margen, el primer y hasta el segundo par de secundarias basales se disuelven en arcos sucesivos uniéndose a los arcos de las demás laterales; glándulas conspicuas y saliente en ambas caras, pequeñas y numerosas, formando un punteado cóncavo en hojas adultas; base redondeada hasta cordada en hojas juveniles; ápice agudo hasta acuminado, con el extremo redondeado; pecíolos 2-3 mm, dorsalmente acanalado, glabro (fig. 2B).
11. Myrcia splendens (Sw.) DC., Prodromus Systematis Naturalis Regni Vegetabilis. 3: 244. 1828. Myrtus splendens Sw., Prodr. 79. 1788; Myrcia costaricensis O. Berg; Myrcia gatunensis Standl.; Myrcia oerstediana O. Berg; Myrcia plicostata O. Berg.
Material examinado: COSTA RICA. Erick Bello C. 4242 (CR); Brad Boyle & S. Ezeta 2772 (CR); J.A. Gonzalez R., Eduardo Lépiz & Juan Francisco Morales 344 (CR); Gladys Rodríguez 271 (MO); B. Hammel 9159 (OET LS). NATIVA.
Hojas de 5-9 x 1.5-3.0 cm, largamente elípticas, cartáceas, discoloras, haz brillante; nerviación camptódroma broquidódroma, nervaduras secundarias 13-18 pares, rectas, sólo curvándose en la unión con la secundaria adyacente, simulando una nervadura intramarginal con escotaduras poco profundas, venas intrsecundarias muy abundantes y conspicuas; glabras o algo pilosas en ambas caras, algo más densa a lo largo de la vena media en la cara adaxial; glándulas numerosas, poco prominentes; base redondeada hasta obtusa; margen entero a veces algo sinuoso; ápice agudo hasta algo acuminado; pecíolos 8 mm, teretes, con pilosidad ócrea caediza (fig. 2F).
12. Myrciaria floribunda (West ex Willd.) O. Berg, Linnaea 27: 330. 1856. Eugenia floribunda H. West ex Willd., Sp. pl. 2: 960. 1799.
Material examinado: COSTA RICA. Quírico Jiménez M. 460 (CR); L. Acosta et al. 1114 (INB); Reinaldo Aguilar & R. Espinosa 3161 (INB); C. Moraga 887 (INB); Alex Rodríguez et al. 6932 (INB); Patrick Harmon 131 (MO); Gerardo Herrera 4401 (MO). NATIVA.
Hojas de 2-8 x 0.7-3 cm, angostamente elípticas hasta ovadas, subcoriáceas, haz lustroso, brillante, concolor o casi; nerviación camptódroma broquidódroma, vena media plana o algo surcada adaxialmente, convexa en la cara abaxial; venas laterales secundarias 15-30 pares, rectas, con venas intersecundarias visibles; arcos de secundarias, sólo curvándose en la unión con la secundaria adyacente, simulando una nervadura intramarginal con escotaduras poco profundas, a ca. 1 mm del margen; ambas caras glabras o con pilosidad muy escasa, tricomas persistentes a lo largo de la vena media, los que se encuentran también en el pecíolo; glándulas inconspicuas en ambas caras o con pocas glándulas convexas abaxialmente; base cuneada, a veces redondeada; márgenes decurrentes por el pecíolo; ápice caudado acuminado o largamente mucronado, no abrupto, con el extremo redondeado o cuspidado; pecíolos 3-6 mm, aplanados o adaxialmente planos, puberulentos, como las ramillas (fig. 2A)
13. Psidium friedrichsthalianum (O. Berg) Nied. Die Natürlichen Pflanzenfamilien, 3(7): 69. 1893. Calyptropsidium friedrichsthalianum O. Berg, Linnaea, 27: 350. 1856.
Material examinado: COSTA RICA. L. Acosta et al. 1803 (INB); R. Rueda & P. Paguaga 12998 (MO); Emilio Castro 45 (OET LS). EXÓTICA (Nativa de Costa Rica).
Hojas de 4-7 x 2-4 cm, ovadas hasta elípticas, coriáceas, brillantes, discoloras; nerviación camptódroma broquidódroma, nervaduras secundarias laterales 6-9 pares, rectas, uniéndose con las secundarias adyacentes en ángulos obtusos, generando escotaduras poco profundas; glabra; glándulas numerosas, conspicuas en ambas caras; base redondeada hasta obtusa; margen poco revoluto; ápice agudo nunca mucronado; pecíolos 6-10 mm, terete, glabro (fig. 2D).
14. Psidium guajava L., Species Plantarum 1: 470. 1753.
Material examinado: COSTA RICA. Juan Francisco Morales 65 (CR); Sandy Salas 45 (INB); James Folson 49735 (MO). NATIVA.
Hojas de 5-10 x 2-4 cm, elípticas hasta oblongas, cartáceas, algo discoloras; nerviación camptódroma eucamptódroma, nervaduras secundarias laterales 15 pares, rectas y marcadas en ambas caras, regularmente espaciadas, arcos de secundarias abruptamente curvados a 3 mm del margen; nerviación terciaria opuesta percurrente; glabras o con pubescencia hirsuta; glándulas numerosas, poco visibles a simple vista; base redondeada a veces cordada; margen entero a veces algo sinuoso; ápice agudo a veces obtuso; pecíolos 10-12 mm, teretes, glabros (fig. 3E).
Clave de determinación
Basados en los patrones de nerviación y otros caracteres foliares se obtuvo la siguiente clave de determinación de las especies presentes en la Estación Biológica La Selva (OET).

Discusión y conclusiones
Las Myrtaceae poseen un patrón de nerviación pinnada, por lo cual las variaciones en los patrones comienzan en las secundarias, las que son variables según la unión entre ellas, su calibre y el ángulo con respecto a la nervadura primaria. Según Klucking (1988) en Myrtaceae el patrón acródromo es el más frecuente, en el que existen uno o más pares de secundarias originadas basalmente que originan arcos marginales llegando al ápice foliar. Cardoso & Sajo (2004; 2006) hallaron que el patrón acródromo era el menos frecuente, siendo el camptódromo el que aparecía en la mayoría de las especies estudiadas. Coincidentemente, no se encontraron especies con patrón acródromo en el presente trabajo, siendo el patrón camptódromo el más frecuente, donde las venas secundarias no terminan en el margen, sino que se fusionan con las demás venas de igual o menor calibre. Dentro de este patrón podemos encontrar las del tipo broquidódromo (brochos = lazo corredizo) donde las nervaduras secundarias se anastomosan desde la base de las hojas formando una serie de arcos próximos al margen. Dependiendo del ángulo de cierre de los lazos, los arcos pueden ser más o menos prominentes, lo que sucede en todas las especies estudiadas excepto E. stipitata y P. guajava, ya que sus arcos pueden formar otros menores de segundo, tercero o cuarto orden, disminuyendo el calibre de las nerviaciones. A este tipo de patrón se le denomina eucamptódromo (Hickey, 1974) o débilmente broquidódromo (Ash et al., 1999). Muchas veces la misma hoja puede poseer una combinación de patrones definidos, por lo que se les denomina mixtos, describiéndose por partes. Los ángulos de inserción de las nerviaciones secundarias son muy variables, al igual que el número y su espaciamiento. Es notable en E. basilaris el bajo número de secundarias, con espaciamiento irregular y con un ángulo casi recto en la porción media de la lámina, a diferencia de E. lithosperma, la cual posee un espaciamiento regular y varias secundarias, con un ángulo de inserción de 30-40°. Los arcos producidos por las secundarias forman ángulos de ca. 90° como en E. corusca o tan obtusos como E. hatshormii, E. lithosperma, M. floribunda o P. friedrichthalianum, que simulan una nervadura intramarginal. Estos arcos pueden encontrarse cercanos o alejados hasta ca. 1 cm del margen como en E. corusca, E. basilaris, E. selvana y E. lithosperma, donde se observa la nerviación última marginal formando arcos de nerviaciones terciarias. Como ocurre en E. stipitata y P. guajava, estos arcos son parte de las secundarias que disminuyen su calibre y forman una sucesión de arcos fusionados con las secundarias supraadyacentes.
Las nervaduras terciarias puede ser del tipo; ramificadas, percurrentes o reticuladas, muchas veces estos patrones se encuentran más de uno en la misma hoja. Las nervaduras terciarias son; a) ramificadas, cuando éstas no se funden a las secundarias, aunque las ramificaciones derivadas de éstas si lo pueden hacer, ocurriendo en todas las especies menos en E. stipitata y P. guajava. Las ramificadas pueden ser admediales o exmediales; b) percurrentes, donde las nervaduras terciarias originadas de las nervaduras secundarias opuestas se funden formando otra nervadura paralela a la secundaria como en E. corusca (alterna percurrente); c) reticulado, cuando las nervaduras terciarias se funden unas con otras o con las nervaduras secundarias; cuando los ángulos de anastomosado son predominantemente rectos, se le llama reticulado ortogonal (Hickey, 1974) u opuesto percurrente (Ash et al., 1999) como sucede en P. guajava; y cuando los ángulos son variables se le llama reticulado propiamente dicho. En muchos casos estas fusiones de terciarias pueden confundirse con la aparición de venas intramarginales, aunque en general estas se originan de la nervadura primaria, esto sucede notablemente en E. basilaris, E. lithosperma, E. sarapiquensis, E. stipitata, M. splendens y P. guajava. La nerviación última marginal cuando es visible, aparece formando arcos en todas las especies, por lo que en este caso no es un carácter útil para discriminar estas especies.
En el cuadro 1 se presenta un resumen de los patrones de nerviación foliar de las especies encontradas en La Selva, dónde el 85% de las especies poseen un patrón de nerviación secundaria camptódromo-broquidódromo y ca. el 50% poseen nervaduras secundarias rectas que sólo se curvan a menos de 2 mm del margen, simulando la presencia de una vena intramarginal.
El uso de los patrones de nerviación combinados con caracteres morfológicos foliares permitieron discriminar las especies y realizar una clave de determinación que podrá ser utilizada con materiales sin estructuras reproductivas para la Estación Biológica La Selva.
Los resultados obtenidos tanto en este estudio como por Cardoso y Sajo (2004; 2006), Jolochin (2008) y Klucking (1988), coinciden en que los caracteres obtenidos de los detalles de los patrones de nerviación foliar pueden utilizarse como caracteres complementarios en la identificación de las especies de Myrtaceae, así como de otras familias, por lo que deberían tenerse en cuenta con mayor frecuencia a la hora de las descripciones de los taxa.
Agradecimientos
Agradezco a la Organización para Estudios Tropicales (OET) por brindarme la oportunidad de realizar esta investigación de corta duración en la Estación Biológica La Selva (Beca postcurso oet18-07 núm. 4156); a los Herbarios del Museo Nacional (CR) y del Instituto Nacional de Biodiversidad INBio (INB) por la consulta de materiales; a Paola Barriga y Mariana del Olmo por la ayuda en la colección de campo; a Fred Barrie, Mauricio Bonifacino y Orlando Vargas en la ayuda en la identificación de las muestras y a Robbin Moran por las sugerencias y aportes a este proyecto.
Literatura citada
Ash, A.; B. Ellis, L.J. Hickey, K. Johnson, P. Wilf y S. Wing, 1999. Manual of Leaf Architecture. Morphological description and categorization of dicotyledonous and netveined monocotyledonous angiosperms. Smithsonian Institution. 65 pp. [ Links ]
Barrie, F.R. 2005. "Thirty-five species of Eugenia (Myrtaceae) from Mesoamerica". Novon, 15(1): 4-49. [ Links ]
Barrie, F.R.; B. Holst, M.L. Kawasaki yL.R. Landrum, 2007. "Myrtaceae". En: Manual de Plantas de Costa Rica, VI: 728-784. [ Links ]
Cardoso Viana, C.M. y M. Sajo, 2004. "Vascularização foliar e identificação de espécies de Eugenia L. (Myrtaceae) da bacia hidrografica do Rio Tibagi, PR". Acta Bot. Bras., 17(1): 47-54. [ Links ]
----------, 2006. "Nervação foliar em espécies brasileiras de Myrtaceae Adans". Acta Bot. Bras., 20(3): 657-669. [ Links ]
González, C.C., 2011. "Arquitectura foliar de las especies de Myrtaceae nativas de la Argentina I: Grupos 'Myrcia', 'Myrceugenia' y 'Plinia'". Bol. Soc. Argent. Bot., 46(1-2): 41-63. [ Links ]
Hickey, L.J., 1974. "Clasificación de la Arquitectura de las hojas de Dicotiledóneas". Bol. Soc. Argent. Bot., 16(1-2): 1-26. [ Links ]
Jolochin, G., 2008. "Revisión de MYRTACEAE Adans. de la Flora uruguaya". Tesis de Ing. Agr. Facultad de Agronomía, Universidad de la República, Uruguay. 2 vol. 302 pp. [ Links ]
Johnson, L.A.S., y B.G. Briggs, 1984. "Myrtales and Myrtaceae - a phylogenetic analysis". Ann. Missouri Bot. Gard., 71: 700-756. [ Links ]
Judd, W.S.; Campbell, C.S., Kellogg, E.A. y P.F. Stevens, 1999. Plant Systematics. A Phylogenetic Approach. Massachusetts, U.S.A. Sinauer Associates, Inc. 464 pp. [ Links ]
Klucking, E.P., 1988. Leafvenation patterns. v 3. J. Cramer, Stuttgart. [ Links ]
Mantese, A.I., y N.H. Montaldo, 2002. "Contribución a la anatomía foliar de Blepharocalyx salicifolius (Myrtaceae)". Bol. Soc. Argent. Bot., 37(3-4): 167-170. [ Links ]
Mc Vaugh, R., 1968. "The genera of american Myrtaceae. An interim report". Taxon, 17: 354-41. [ Links ]
Romero, E.J., y M. Dibbern, 1982. "Arquitectura foliar de las especies de Myrtaceae de la Provincia de Buenos Aires". Bol. Soc. Argent. Bot., 20(3-4): 255-266. [ Links ]
Watson, L., y M.J. Dallwitz, 2007. "The Families of Flowering Plants" [en línea]. < http://delta-intkey.com> [marzo 2012] [ Links ].
Wilson, P.G.; O'Brien, M.M., Gadek, P.A. y C.J. Quinn, 2001. "Myrtaceae revisited: a reassessment of infrafamilial groups". Am. J. Bot., 88(11): 2013-2025. [ Links ]
Zomlefer, W., 1994. Guide to Flowering Plant Family. The University of North Carolina Press, Chapel Hill & London. 430 pp. [ Links ]

