










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkPolibotánica
versão impressa ISSN 1405-2768
Polibotánica no.37 México Fev. 2014
Influencia de la maduración del fruto de Arbutus xalapensis kunth sobre la germinación de semillas y embriones cigóticos
Influence of fruit ripening of Arbutus xalapensis kunth on the germination of seeds and zigotyc embryos
Violeta Tovar-Rocha, Ma. del Carmen Rocha-Granados y Patricia Delgado-Valerio
Facultad de Agrobiología "Presidente Juárez", Universidad Michoacana de San Nicolás de Hidalgo. Av. Lázaro Cárdenas s/n, esq. Berlín. Col. Viveros. CP 60190. Uruapan, Mich. Correo electrónico: crochagra05@gmail.com.
Aceptado: 5 julio 2013.
Recibido: 20 febrero 2013.
Resumen
En este trabajo se determinó el efecto del estado de maduración del fruto en la germinación in vitro de semillas y embriones cigóticos de Arbutus xalapensis Kunt. Se colectaron frutos de 10 árboles en cada uno de los dos sitios de estudio y se clasificaron según su tamaño y peso dentro de tres grupos caracterizados por el color del fruto: 1 a 6 frutos verde oscuro (FVO), 7 y 8 frutos verde-amarillo (FVA), y 9 y 10 frutos naranja-rojizo (FNR). De cada estado se seleccionaron 50 semillas y se colocaron en medio MS para evaluar su germinación. El experimento se repitió dos veces y los resultados obtenidos fueron sometidos a un análisis de comparación de medias de Tukey el cual indicó que los tres grupos de frutos presentan características de peso fresco y diámetro diferentes (P ≤ 0.05), con un promedio de 6.3 g y de 0.21 mm para FVO, 9.5 g y 0.46 mm para FVA y 10.8 g y 0.70 mm para FNR. El análisis de microscopía estereoscópica demostró que todos los estados de maduración presentan semillas y embriones; los estados 1 al 4 presentan embriones no desarrollados, y en los estados 5 al 10 los frutos contienen un mayor número de semillas con embriones desarrollados y con una mayor germinación (7.6 a 8.8 semillas/unidad experimental). Se propone el uso de semillas provenientes de frutos de los estados de maduración 5 al 10 para la germinación adecuada de semillas de madroño, mientras que para los embriones cigóticos es necesario realizar estudios de prueba de medios de cultivo para su germinación.
Palabras clave: clasificación de frutos, embriones, germinación, madroño, microscopia, semillas.
Abstract
The aim of this study was to determine the effect of fruit ripeness of in vitro germination of seeds and zygotic embryos of Arbutus xalapensis Kunth. Fruits were collected from 10 trees in each of the two study sites and were classified according to their size and weight in 10 states into three groups characterized by the color of the fruit: 1 to 6 dark green fruits (DGF), 7 and 8 yellow-green fruits (YGF) and 9 and 10 orange-red fruits (ORF). 50 seeds of each state were selected and placed on MS medium to evaluate their germination. The experiment was repeated two times and the results were submitted to Tukey's analysis of mean comparison which indicated that the three groups have different characteristics related to fresh weight and diameter (P ≤ 0.05), with an average of 6.3 g and 0.21 mm for DGF, 9.5 g and 0.46 mm for YGF and 10.8 g and 0.70 mm for ORF. The stereomicroscopic analysis showed that all states have seed maturation and embryos; states one to four embryos have developed, and in the states 5 to 10, fruits contain a greater number of seeds with embryos developed and higher germination (7.6 to 8.8). Therefore, is proposed the use of ripening fruit from stages 5 to 10 for right germination of seeds of strawberry tree, while for zygotic embryos is necessary to carry out studies of culture media for its germination.
Key words: classification of fruit, embryos, germination, strawberry tree, microscopy, seeds.
Introducción
Michoacán es una región maderera muy reconocida a nivel nacional por la elaboración de artesanías, la cual es una tradición que se ha practicado desde tiempos remotos y que actualmente es fuente de ingresos para una gran parte de su población (Rzedowski, 1985). Para dicha práctica se emplean especies como Alnus arguta (Schltdl.) Spach (aile), Tilia mexicana Schlechtendal (cirimo) y Platymiscium lasiocarpum Sandwith (granadillo) que se aprecian y usan para la confección de guitarras, y Arbutus xalapensis Kunth (madroño) la cual presenta excelentes cualidades para la fabricación de artículos torneados y/o artesanales (Guridi, 1981; Rzedowski, 1985, 2006; Pérez, 1993; Comisión Forestal del Estado de Michoacán, 2009). Arbutus xalapensis pertenece a la familia Ericaceae, es un árbol de 10 a 20 metros de altura, perennifolio, cuyo tronco y ramas poseen corteza amarillento-rojiza, delgada, lisa y frecuentemente exfoliante, sus frutos son en forma de baya globosa con el pericarpio granular, 5-locular y tienen varias semillas (Hernández et al., 1991). Se distribuye en los estados de Chihuahua, Nuevo León, Durango, Sinaloa, Jalisco, Oaxaca, Michoacán, San Luis Potosí, Guanajuato, Hidalgo, Puebla, Tlaxcala y Veracruz. Forma parte del matorral xerófilo, de los bosques de pino-encino o de encino sólo (González y González, 1992). Sin embargo, actualmente sus poblaciones han disminuido presentando una distribución restringida y fragmentada, lo cual ha sido originado por diferentes factores como son la sobreexplotación de su madera, el sobre-pastoreo y la presencia de plagas forestales, y como consecuencia, se tiene una disminución en la producción de frutos y semillas, lo cual implica una baja regeneración y con ello la pérdida del material genético nativo de esta especie (INAFED, 2009). Por tanto, es de una importancia inmediata realizar estudios que permitan proponer metodologías de propagación vegetal eficientes para salvaguardar este recurso.
A pesar de que el madroño es una especie de importancia ecológica y económica, hasta el momento ha sido poco estudiada. Por ejemplo, se ha visto que en especies cercanamente relacionadas a madroño como Arbutus unedo L., se presentan problemas de germinación debido a que la pulpa del fruto contiene inhibidores (proteasas) que hacen que los frutos que caen al suelo permanezcan en él por un largo periodo hasta que se desintegran, y a menos de que las semillas sean separadas de la pulpa del fruto no germinan (Narbona et al., 2003). Las plántulas que logran germinar no toleran el estrés por sequía durante el verano en sitios expuestos, por lo que su establecimiento se restringe a zonas con una densa vegetación (Mesleard y Lepart, 1991). Para A. xalapensis Kunth no existen estudios que corroboren este comportamiento, siendo más estudiada a nivel de la calidad de la madera y sobre el efecto de los factores ambientales (temperatura y estrés hídrico) en su adaptabilidad y evolución (Aguilar y Barajas, 2005). Sólo existen dos estudios muy generales sobre el efecto que tiene la calidad de semilla o el grado de maduración del fruto sobre su germinación; uno de ellos es el de Nokes (1986), en donde señala que esta especie se multiplica mediante semilla fresca obtenida de frutos bien maduros con lo cual se asegura un alto porcentaje de germinación. Mientras que el segundo estudio es sobre reproducción asexual de A. xalapensis Kunth, la cual se logra utilizando estacas juveniles colocándolas en una cámara humedad a una temperatura de 24 a 28°C (Tipton, 1988). Un estudio reciente en donde se hace una clasificación de frutos es el realizado por Kaur y colaboradores (2009), quienes analizaron frutos de Chlorophytum borivilianum (planta herbácea originaria de la India) agrupándolos según su color y tamaño. Debido a la importancia que guarda el estado de desarrollo de los embriones cigóticos en estudios morfogenéticos in vitro de plantas leñosas, el objetivo de este trabajo fue determinar el estado óptimo de maduración del fruto de Arbutus xalapensis para la germinación in vitro de semilla y embriones cigóticos.
Material y métodos
Sitios de estudio
Primero, se hizo una revisión bibliográfica para conocer las zonas de distribución de la especie. Se exploraron cuatro sitios en los municipios de Uruapan, Pátzcuaro, Quiroga y Zacapu, de los que se seleccionaron dos sitios representativos de la especie con un mayor número de árboles maduros, reproductivos y con presencia de frutos. El primer sitio está ubicado cerca de la localidad de San Miguel Charahuén, municipio de Pátzcuaro (N 19°00'19.5" y W 101°51'05.2", altitud de 1 990 m.s.n.m.), y el segundo sitio en San Jerónimo, municipio de Quiroga (N 19°41'46" y W 101°35'37.3", altitud de 2 199 m.s.n.m.) (fig. 1). Los sitios pertenecen al bosque templado frío donde se encuentran principalmente Pinus, Abies, Quercus, Cupressus, Alnus y Arbutus (Rzedowski, 2006).
Material vegetativo
La colecta se realizó durante los meses de mayo a junio (2010 y 2011), periodo en el que el madroño está en su fase de fructificación. Se colectaron racimos de frutos en distintos estados de maduración de 10 árboles reproductivos para cada uno de los dos sitios. Los racimos fueron colocados en bolsas de papel de estraza, etiquetándolas con el sitio y fecha de colecta, los cuales fueron almacenados en el laboratorio de Fisiología Vegetal de la Facultad de Agrobiología "Presidente Juárez" de la Universidad Michoacana de San Nicolás de Hidalgo para su posterior procesamiento y análisis.
Desinfección de la semilla
Para la desinfección de las semillas se hicieron lavados de los frutos con jabón líquido y agua corriente, enseguida se colocaron en una solución con jabón en polvo más 1 g L-1 de Agrimicin® por espacio de 30 min en agitación, se enjuagaron con agua corriente y se procedió a su desinfección en una campana de flujo laminar. Los frutos fueron colocados en etanol al 70% v/v por 45 segundos y posteriormente en una solución desinfectante de hipoclorito de sodio (Cloralex®) al 40% por 30 min adicionado con Tween 20 al 0.1%. Finalmente, se enjuagaron de tres a cinco veces con agua destilada estéril, condiciones en las cuales se pudo obtener un 95% de semillas estériles, encontrándose que a menor concentración del desinfectante o menor tiempo de desinfección de los frutos provenientes del campo, las semillas presentaron contaminación por hongos o bacterias en los cultivos (datos no mostrados). Debido a que las semillas de madroño presentan una testa o cubierta dura, una vez desinfectados los frutos, fue necesario dejarlos en agua destilada estéril durante 24 h en refrigeración a 8°C, lo cual permitió una mejor extracción de los embriones cigóticos a partir de las semillas, para ser sembrados en el medio MS (Murashige y Skoog, 1962).
Clasificación de los frutos
Como no existía una escala de clasificación de frutos para madroño, primeramente se realizó un análisis del peso (g), tamaño (mm) y color de 60 frutos de cada árbol, y de esta manera nos permitió obtener un sistema de clasificación en 10 estados de maduración de frutos. Posteriormente, los frutos de cada estado fueron seccionados de manera longitudinal para exponer las semillas y embriones y poder observarlos en un microscopio compuesto (Leica DM 1000) a aumentos de 20X y 40X, esto con la finalidad de corroborar el grado de desarrollo de las semillas y de sus respectivos embriones para cada uno de los 10 estados establecidos anteriormente.
Germinación
Los experimentos de germinación se hicieron con 50 semillas de los estados de maduración del 5 al 10, correspondiendo a los frutos de los tres grupos de maduración: frutos inmaduros estados 5 y 6 (FVO), frutos intermedios estados 7 y 8 (FVA) y frutos maduros estados 9 y 10 (FNR), mientras que para las pruebas de germinación de embriones cigóticos se utilizaron 50 embriones provenientes de los frutos intermedios (estados 7 y 8). En campana de flujo laminar, se procedió a sembrar en caja Petri (60 x 15 mm) 10 semillas y 10 embriones cigóticos por unidad experimental con cinco repeticiones en dos experimentos independientes, colocándolos en el medio MS a la mitad de su concentración de sales minerales, sacarosa y vitaminas MS, y gelificado con 8 g L-1 de agar plant TC (PhytoTechnology). Una vez sembrados se colocaron en un cuarto de crecimiento a una temperatura de 24°C ± 1°C y un fotoperiodo de 16 h luz y 8 h oscuridad, evaluando el número de semillas y embriones cigóticos germinados después de 30 días.
Análisis estadístico
Los valores obtenidos de tamaño y peso para cada uno de los diez estados de maduración de los frutos fueron promediados para obtener sus medias y desviaciones estándar. Para el resto de los experimentos se utilizó un diseño experimental completamente al azar, con tres tratamientos y cinco repeticiones, para efecto de la maduración de frutos sobre la germinación de semillas, y dos tratamientos con cinco repeticiones para la comparación de la germinación de semillas y embriones en donde la unidad experimental en cada caso consistió en una caja Petri con 10 semillas y 10 embriones cigóticos, respectivamente. Los resultados obtenidos se sometieron a un análisis de varianza y prueba de comparación de medias (Tukey, P ≤ 0.05) utilizando el paquete estadístico Java Memory Profiler (JMP, versión 7, año 2008 ).
Resultados y discusión
Clasificación del fruto
Se encontró que los estados 1-6 muestran un crecimiento más lento con un intervalo de 0.00 a 0.37 mm con una diferencia media aproximada de 0.40 g y de 0.05 mm entre estados. Una vez que comienza el proceso de maduración y éstos pasan de un color verde oscuro a un verde-amarillo y naranja-rojizo se observa un aumento en peso y tamaño de los frutos teniendo una diferencia en peso y tamaño de hasta 1 g y 0.10 mm, respectivamente (fig. 2, cuadros 1 y 2). Con base en estos resultados se propone la agrupación de los frutos en tres grupos o escalas de maduración: a) frutos inmaduros (FVO), del estado 1-6; b) frutos intermedios (FVA), estados 7 y 8; y c) frutos maduros (FNR), estados 9 y 10; los cuales fueron diferentes estadísticamente (P ≤ 0.05). Esto indica que el peso de los frutos es importante para determinar el grado de maduración de los frutos, mientras que visualmente, el color es un factor fundamental para dicho parámetro. El cambio de color es la característica más notoria en muchas frutas durante su maduración, y por ello se utiliza como criterio para definir la madurez de una fruta (Wills et al., 1998; Reid, 2002; Yommi y Godoy, 2002). La transformación más importante es la degradación del color verde, la cual está asociada con la síntesis o desenmascaramiento de pigmentos cuyos colores oscilan entre el amarillo (carotenoides) y el rojo-morado (antocianinas) (Kays, 2004).

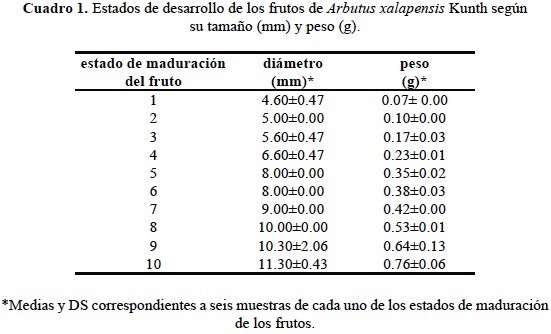
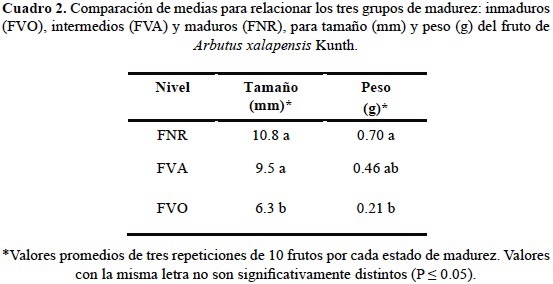
En cuanto al análisis de las variables peso y tamaño de los tres grupos de frutos clasificados según su grado de maduración: frutos inmaduros (FVO), 2) frutos intermedios (FVA), y 3) frutos maduros (FNR), se observaron diferencias significativas, siendo los frutos inmaduros completamente diferentes a los frutos intermedios y maduros, no habiendo diferencias significativas entre estos dos últimos grupos (cuadro 2). Un estudio similar a la clasificación aquí propuesta es el realizado por Pinzón et al. (2007) quienes determinaron, según el color de la cáscara, seis estados de madurez de la gulupa (Passiflora edulis Sims.) desde totalmente verde (estado 0) hasta sobremaduro (color púrpura intenso, estado 6) para facilitar el reconocimiento del momento óptimo de cosecha.
Análisis de microscopía
Este análisis demostró que todos los estados de madurez de los frutos presentan semilla y embriones cigóticos desde el más inmaduro (fruto núm. 1) hasta el más maduro (fruto núm. 10) (figs. 3b, c y d). También se pudo determinar la medida que éstos presentan observándose que las semillas suelen tener una longitud de 2.8 mm, mientras que los embriones tan sólo son de 1.7 mm (figs. 3c y d). Asimismo, se detectó que el grado de desarrollo de éstas varía de un estado a otro observándose menor desarrollo de semillas en los estados del 1 al 6, y un mayor desarrollo de ésta a partir del estado 7 (fig. 3d). La extracción de la semilla sólo fue posible a partir del estado 5, lo cual dio la pauta para elegir los frutos de los estadíos 5 al 10 para realizar los estudios de germinación. Las observaciones microscópicas de las semillas, y sus embriones, mostraron claramente que el desarrollo de la semilla va acompañado de la maduración de los frutos, suceso que se da en la mayoría de las plantas angiospermas (Willan, 1987). Estos resultados concuerdan con los obtenidos por Buzeta (1997), quien determinó que existe una relación entre el contenido de semillas en el fruto y el tamaño de éste.

Germinación de semilla y embriones cigóticos
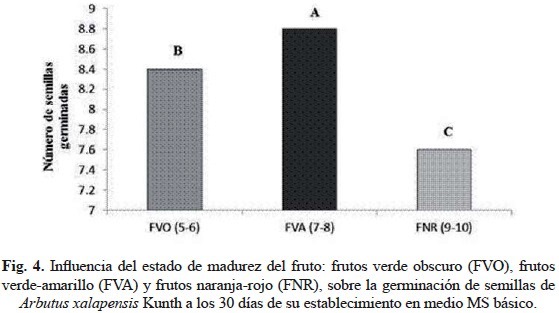
Los resultados obtenidos del análisis estadístico referente a la germinación de semillas mostró diferencias significativas, observándose una mayor germinación en las semillas provenientes de frutos intermedios (8.8 semillas/unidad experimental) en comparación con las semillas de fruto inmaduros (8.4 semillas/unidad experimental) y maduros (7.6 semillas/unidad experimental), siendo las semillas de los frutos maduros las que mostraron la menor respuesta germinativa (fig. 4), lo que indica que el estado de madurez del fruto tiene influencia sobre la misma. Los resultados obtenidos en este experimento muestran que la germinación de las semillas in vitro en esta especie (A. xalapensis) es afectada por el grado de madurez del fruto, en donde a mayor madurez menor poder de germinación, condición que puede deberse a los mecanismos intrínsecos de las semillas en donde antes de entrar en un periodo de latencia hay una pérdida de humedad e inhibición de su metabolismo, así como producción de inhibidores químicos (a-amilasas) que les permite permanecer en climas con periodos de sequía y frío prolongados (Rodríguez y Vázquez, 1992). Algunos estudios realizados con semillas de varias especies de Arbutus han demostrado que algunas pierden su poder germinativo en cortos periodos (A. texan y A. unedo), mientras que otras requieren temperaturas frías por encima del punto de congelación para poder germinar (A. menziessi Pursh) (McDonald, 1978; Nokes, 1986). Narbona et al. (2003) encontraron que la germinación de Arbutus unedo L., una especie de madroño que crece en el mediterráneo, es afectada por la pulpa que contienen sus frutos concluyendo que ésta debe de presentar algún tipo de inhibidores químicos sin tener la posibilidad de germinar mientras permanezcan adheridas a la pulpa. Para Arbutus xalapensis H.B.K., se conoce poco sobre su reproducción y se considera que presenta una baja tasa de reproducción (Stager et al., 1973). Aunque la semillas se producen en abundancia (Vines, 1960), se ha observado en condiciones de laboratorio que su germinación puede variar entre el 20 al 90%, dependiendo del grado de humedad (Wiedenfeld, 1975). Comportamiento que es semejante a las semillas recalcitrantes pues pierden rápidamente su capacidad de germinar al quedar expuestas a condiciones de baja humedad (Kermode y Finch-Savage, 2002). Sin duda alguna, la eficiencia de germinación y desarrollo de las semillas está muy correlacionado con el grado de maduración de los frutos, pues se ha observado que semillas de berenjena provenientes de frutos inmaduros son deficientes en germinación, mientras que aquellas que provienen de frutos maduros, además de tener mayor tamaño, aumentan su porcentaje de germinación (Passam et al., 2010), fenómeno que es observado también en jitomate, pues Heidary y Dastjerdi (2012) reportan que la semilla proveniente de frutos de color rojo presentan mayor eficiencia de germinación que los provenientes de frutos verdes, resultados que difieren de lo obtenido en este estudio, pues a mayor coloración y maduración de los frutos, menor eficiencia sobre la germinación de las semillas de madroño.
De manera opuesta a lo observado con la semilla, se encontró que la germinación de los embriones cigóticos, extraídos de las semillas y sembrados en un medio MS, disminuye de manera considerable (0.4 embriones/unidad experimental) (fig. 5), por lo que es muy probable que el medio de cultivo influya más en su germinación que el estado de desarrollo del embrión. El madroño es una especie leñosa para las que se ha observado que el medio de cultivo es fundamental en las repuestas morfogenéticas in vitro de los embriones. Por ejemplo, los embriones cigóticos de varias especies de pinos requieren de medios suplementados con maltosa, nitrato de plata, nitrato de magnesio y casa aminoácidos para su germinación, formación de callo o bien para la formación de embriones somáticos (Ramarosandratana et al., 2001a; Pullman et al., 2003; Miguel et al., 2004; Niskane et al., 2004; Lelu-Walter y Bernier-Cardou, 2006). En casos específicos como el reportado por Pulman et al. (2003) en Pinus tadea L, para que los embriones cigóticos puedan establecerse se requiere de un medio suplementado con mio inositol, ácido abscísico y nitrato de plata, donde además consideran que el estado de maduración de éstos es fundamental para la iniciación de los cultivos, ya que los embriones inmaduros producen mayor cantidad de callos embriogénicos, condición que también se ha observado por Lelu-Walter y Bernier-Cardou, (2006), en Pinus pinaster Ait. Una especie no leñosa para la que se ha observado la importancia del estado de desarrollo de los embriones es Glycine max (L.) Merrill. Para esta especie se encontró que la germinación es mayor en los embriones cigóticos provenientes de semillas más maduras (90%) que en los provenientes de semillas inmaduras (43%), condición que se mantiene siempre y cuando los embriones sean cultivados en un medio B5 (Gamborg) conteniendo 1% de sacarosa (Tekrony et al., 1996).

Conclusiones
Los resultados generados en este estudio permitieron establecer una escala de clasificación de frutos basada en su tamaño y color: frutos verde obscuro (FVO) o inmaduro (estado 1 al 6), frutos verde amarillo (FVA) o intermedios (7 y 8), y frutos naranja rojo (FNR) o maduros (9 y 10).
Las observaciones microscópicas corroboraron que en los 10 estados de los frutos se presentan semillas y embriones, sin embargo los estados del 5 al 10 contienen un mejor desarrollo de éstos.
El estado de madurez del fruto influye sobre la germinación de las semillas, observándose que a mayor madurez de frutos menor cantidad de semillas germinadas, mientras que los resultados de eficiencia de germinación de embriones cigóticos mostraron una drástica deficiencia en su germinación. Esto indica que probablemente lo que más influye en la germinación de los embriones sea el medio de cultivo. Por tanto, se sugiere realizar pruebas de germinación de embriones en diferentes medios de cultivo, así como estudios de morfogénesis in vitro con semillas provenientes de frutos de los estados 5 al 10.
Agradecimientos
Este trabajo fue financiado por el Consejo Nacional de Ciencia y Tecnología (CONACYT) con una beca de maestría núm. 239177 de Violeta Tobar Rocha, por el Programa de Mejoramiento del Profesor (PROMEP-SEP) y por la Coordinación de la Investigación Científica (CIC) de la Universidad Michoacana de San Nicolás de Hidalgo a través de los proyectos PTC-292, y CIC-2011 de Ma. del Carmen Rocha Granados.
Literatura citada
Aguilar, R.S., y J. Barajas M., 2005. "Anatomía de la madera de especies arbóreas de un bosque mesófilo de montaña: un enfoque ecológico-Evolutivo". Boletín de la Sociedad botánica de México, diciembre, núm. 077. Sociedad botánica de México, A.C., Distrito Federal, México. pp. 51-58. [ Links ]
Buzeta, A., 1997. Chile: Berries para el año 2000. Departamento Agroindustrial, Fundación Chile. Santiago, Chile. 135 pp. [ Links ]
Comisión Forestal del Estado de Michoacán, enero de 2009. Boletín informativo núm. 2. [ Links ]
Guridi, L.C., 1981. "La Madera en las artesanías del estado de Michoacán". Boletín divulgativo núm. 50 Subsecretaría Forestal y de la Fauna, Instituto Nacional de Investigaciones Forestales. SARH. Diciembre 1980. [ Links ]
Hardesty, W.D., y D.C. Whitenberg, 1976. Texas madrone. Texas Parks and Wildlife. 34: 24-26. [ Links ]
Heidary, M., y A.M.A. "Dastjerdi. Effect of fruit maturity on seed germination of tomato (Lycopersicon licopersicon)". www.science.alzahra.ac.ir/Portals/0/Majlat (accesada en abril de 2012). [ Links ]
Hernández, S.L., C. Romo, y F. González M., 1991. "Plantas útiles de Tamaulipas", Anales, Serie Botánica. Instituto de Biología, UNAM, 2(1): 1-38. [ Links ]
INAFED (Instituto Nacional para el Federalismo y el Desarrollo Municipal), 2009. Enciclopedia de los Estados de México: Michocán, Medio Físico. Gobierno del Estado de Michoacán. http://www.e-local.gob.mx/work/templates/enciclo/michoacan (accesada en marzo de 2010). [ Links ]
Kaur, R.; S. Arora, y A.K. Thukral, 2009. "Enhancing seed germination of Chlorophytum borivilianum Sant. et Fernand. with PGRs, steroidal hormones and zinc". Research Journal of Seed Science, 2: 32-39. [ Links ]
Kays, S., 2004. Postharvest biology. Exon Press, Athens, Georgia. 568 pp. [ Links ]
Kermode, H.R., y W.E. Finch-Savage, 2002. "Desiccation sensitivity in orthodox and recalcitrant seeds in relation to development". In: Black, M. y H.W. Prichard, editors. Desiccation and survival in plants. Drying without dying. CABI Publishing. pp. 149-184. [ Links ]
Lelu, M.A.; M. Bernier., y K. Klimaszewska, 2006. "Simplified and improved somatic embryogenesis for clonal propagation of Pinus pinaster (Ait.)". Plant Cell Reports, 25:767-776. [ Links ]
McDonald, P.M., 1978. Silviculture-ecology of three native California hardwoods on high sites in north-central California. [PhD Dissertation]. Oregon State University. Department of Forest Science. 309 pp. [ Links ]
Mesleard, F., y J. Lepart, 1991. "Germination in seedling dynamics of Arbutus unedo and Erica arborea on Corsica". Journal of Vegetation Science, 2: 155-164. [ Links ]
Miguel, C.; S. Goncalves., S. Tereso., L. Marum., y M.M. Oliveira, 2004. "Somatic embryogenesis from 20 open-pollinated seed families of Portuguese plus tree of maritime pine". Plant Cell, Tissue and Organ Culture, 76: 121-130. [ Links ]
Murashige, T., y F. Skoog, 1962. "A revised medium for rapid growth and bioassays with tobacco tissue cultures". Plant Physiology, 15: 473-479. [ Links ]
Narbona, E.; M. Arista, y P. L. Ortiz, 2003. "Germinación de las semillas de madroño (Arbutus unedo L. Ericaceae)". Acta Botánica Malacitana, 28: 73-78. [ Links ]
Niskane, A-M.; J. Lu., S. Seitz., K. Keinonen, K. von Weissenberg, y A. Pappinen, 2004. "Effect of parent genotype on somatic embryogenesis in Scots pine (Pinus sylvestris)". Tree Physiology, 24: 1259-1265. [ Links ]
Nokes, J., 1986. How to grow native plants of Texas and the southwest. Texas Monthly Press, Austin, Texas. USA. [ Links ]
Passam, M.C.; D. Makrogianni., F. Gregriou., y I.C. Karapanos, 2010. "The size and germination of eggplant seed is relation to fruit maturity at harvest, after-ripening and ethylene application". Analele Universitatii-Fascicula Biologie. Tom XVII, Issue 2. pp: 225-229. [ Links ]
Pérez, O.C.P., 1993. "Anatomía de la madera de ocho especies con importancia en las artesanías del Estado de Michoacán". Acta Botánica Mexicana núm. 23. Instituto de Ecología, A.C. Pátzcuaro, México. pp. 103-136. [ Links ]
Pinzón, I.M.P.; G. Fischer, y G. Corredor, 2007. "Determinación de los estados de madurez del fruto de la gulupa (Passiflora edulis Sims.)". Agronomía Colombiana, 25(1): 83-95. [ Links ]
Pullman, G.S.; S. Johnson., G. Peter., J. Cairney, y N. Xu, 2003a. "Improving loblolly pine somatic maturation: comparison of somatic and zygotic embryo morphology, germination, and gene expression". Plant Cell Reports, 21: 747-758. [ Links ]
Pullman, G.S.; K. Namjoshi, y Y. Zhang, 2003b. "Somatic embryogenesis in loblolly pine (Pinus taeda L.): improving culture initiation with abscisic acid and silver nitrate". Plant Cell Reports, 22: 85-95. [ Links ]
Ramarosandratana, A.; L. Harvengt, A. Bouver, R. Calvayrac, y M. Paques, 2001a. "Effects of carbohydrate source, polyethylene glicol and gellan gum concentration on embryonal-suspensor mass (ESM) proliferation and maturation of maritime pine somatic embryos". In vitro Cellular and Developmental Biology-Plant, 37: 29-34. [ Links ]
Reid, M.S., 2002. "Maturation and maturity indices". En: Kader, A. A. (Ed.) Post-harvest technology of horticultural crops. 3th ed. University of California, Agricultural and Natural. pp. 55-62. [ Links ]
Rodríguez, C., y C. Vázquez Y., 1992. "La conservación de las plantas en peligro de extinción a través del almacenamiento a largo plazo de semillas". Interciencia, 17: 293-297. [ Links ]
Rzedowski, J., y G. Calderón R., 1985. Flora fanerogámica del valle de México. Instituto Nacional de Ciencias Biológicas. Instituto de Ecología. México, DF. [ Links ]
Rzedowski, J., 2006. Bosque de coníferas. En: Vegetación de México. Primera edición digital. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. México. [ Links ]
Steger, R.; E. Beck., y F. Reldon, 1973. "Range plants as ornamentals". Journal of Range Management, 26: 72-74. [ Links ]
Tekrony, D.M.; L.J. Grabau., M. Delary, and M. Kane, 1996. "Early planting of early maturing soyabean:effects on seed germination and phomopsis infection". Agronomy Journal, 88(3): 428-433. [ Links ]
Thagana, W.P.; C.M. Ndirangu., E.O. Omolo., T.C. Riungu., M.G. Kinyua., N.W. Ndembei, and W.K. Koskey, 2005. "In vitro embryo culture and immature seed germination techniques for rapid generation advance in soyabean [Glycine max (L.) Merrill]". Proceedings of the African Crop Science Conference, Kampala, Uganda, 5-9th December. pp. 269-272. [ Links ]
Tipton, J.L., 1988. "Asexual propagation of juvenile Arbutus xalapensis in a high humidity chamber". Plant Propagator, 27: 11-12. [ Links ]
Vines, R.A., 1960. Trees, shrubs, and woody vines of the Southwest. Austin, TX: University of Texas Press. 1104 pp. [ Links ]
Wiedenfeld, C.C., 1975. "The Texas madrone". Texas Horticulturist, 2(1): 14-16. [ Links ]
Willan, R.L., 1987. A guide to forest seed handling. Chapter 2: "Seed and fruit development germination, dormancy". Food and Agriculture Organization of the United Nations. Rome. M-13 ISBN 92-5-102291. [ Links ]
Wills, R.; B. McGlasson, D. Graham, y D. Joyce, 1998. Postharvest: an introduction to the physiology and handling of fruit, vegetables and ornamentals. CAB International, Nueva York. 262 pp. [ Links ]
Yommi, A., y C. Godoy, 2002. Arándanos: Fisiología y Tecnologías de Postcosecha. http://www.elsitioagricola.com/gacetillas/balcarce/bl20021029/arandanos.asp (accesada en agosto de 2012). [ Links ]

