










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPolibotánica
versión impresa ISSN 1405-2768
Polibotánica no.36 México ago. 2013
Germinación y morfología de alpiste (Phalaris canariensis L.) Poaceae, bajo distintas condiciones de calidad de luz
Germination and morphology of canary grass (Phalaris canariensis L.) Poaceae, under different conditions of light quality
M. S. Ferrarotto y H. Da Silva
Universidad Central de Venezuela. Facultad de Agronomía. Maracay, Edo. Aragua. Apartado Postal 4579. Venezuela. Correo electrónico: ferrarottom@agr.ucv.ve.
Recibido: 10 abril 2012.
Aceptado: 12 diciembre 2012.
Resumen
El alpiste (Phalaris canariensis L.) es utilizado como material vegetal y planta indicadora en estudios de detección y caracterización de virus, alelopatía, control biológico y bioquímica de sustancias bioactivas funcionales. No obstante, no se encontraron antecedentes sobre la morfología de plántulas y respuesta germinativa in vitro. Esta investigación se plantea como objetivo, estudiar aspectos morfológicos de plántulas de alpiste y la respuesta germinativa in vitro bajo diferentes calidades de luz, las cuales podrían ser de utilidad en el estudio e interpretación de resultados en la investigación aplicada en plantas. Se expusieron las semillas a tratamientos en cápsulas de Petri: T1, luz natural; T2, luz roja; T3, luz roja-rojo lejano; T4, luz roja-rojo lejano-roja; T5, oscuridad; T6, filtro transparente; T7, filtro azul; T8, filtro verde y T9, filtro rojo. Las semillas resultaron fotoblásticas negativas, con 48% de germinación en oscuridad y 16% bajo filtro rojo, el crecimiento de la radícula fue inhibido en todos los tratamientos con luz, siendo más corta bajo filtro verde (0.8 cm). Por el contrario, combinaciones de luz roja, rojo lejano y de oscuridad estimularon su alargamiento hasta 7.8 cm. La luz natural inhibió el crecimiento del coleoptilo en la misma magnitud que el filtro verde (0.8 y 0.7 cm, respectivamente). Bajo luz roja y combinaciones con rojo lejano las plántulas se etiolaron mostrando un coleoptilo entre 9.4-10.3 cm y la primera hoja entre 13.2-14 cm. El contenido de clorofila fue mayor en luz que en oscuridad (6.42 x10-3 y 3.49x10-3 mg mL-1, respectivamente).
Palabras clave: alpiste, germinación, luz, morfología, Phalaris canariensis.
Abstract
The canary grass (Phalaris canariensis L.) is used as vegetal material and indicative plant in studies for virus detection and characterization, allelopathic effects, biological control and biochemistry of bioactive functional substances. Nevertheless, no previous reports upon germination response and morphological aspects of seedlings growing in vitro were found. The objectives of this study were to study morphological aspects of seedlings of canary grass and the germination response in vitro, under different light qualities, which would be useful in the interpretation of further results in plant research. Seeds were exposed to treatments in Petri dishes: T1, natural light; T2, red light; T3, red light-far red light; T4, red light-far red light-red light; T5, darkness; T6, transparent filter; T7, blue filter; T8, green filter and T9, red filter. Results show that seeds are photoblastic negative with 48% germination in the dark in 48h and 16% under red filter. The natural light and all the filters suppress the radicle growth, shorter ones were observed under the green filter (0.8 cm). Red light alone and its combination with far red, as well as darkness stimulated radicle elongation up to 7.8cm. Natural light suppressed coleoptile growth in the same magnitude than with green filter (0.8 and 0.7 cm respectively). Under red light and its combination with far red, seedlings were etiolated showing a coleoptile between 9.410.3 cm and the first leaf between 13.2-14 cm. Chlorophyll content was higher in light than in darkness (6.42 x 10-3 y 3.49 x 10-3 mg mL-1, respectively).
Key words: canary grass, germination, light, morphology, Phalaris canariensis.
INTRODUCCIÓN
El alpiste (Phalaris canariensis L.) pertenece a la familia de las Poáceas y es originaria del Mediterráneo. A diferencia de P. paradoxa y P. minor, consideradas como especies invasoras, P. canariensis es la única especie del género Phalaris cultivada para la producción de grano (SENASA, 2011). Representa una especie promisoria para su uso en investigación biológica en estudios sobre alelopatía y control biológico (Truol etal., 2001), detección de virus (Enamorado et al., 2004) y bioquímica de sustancias bioactivas funcionales en plantas (Paredes et al., 2009). En este sentido, el alpiste, se ha utilizado en la caracterización del virus del mal de Río Cuarto (MRCV) que afecta al maíz (Zea mays L.) (Enamorado et al., 2004), y en bioensayos para determinar el efecto alelopático de hongos filamentosos sobre la germinación de semillas (Truol et al., 2001), así como en el estudio de la presencia de la melatonina en plantas (Paredes et al., 2009).
Sin embargo, aun cuando en la literatura hay autores que afirman que es una planta promisoria para investigación aplicada, tal como Cataús (1997) quien consideró que el alpiste podría resultar una excelente planta para estudios genéticos debido a su bajo número cromosómico 2n=12, no se encontraron antecedentes relacionados con aspectos morfológicos de las plántulas y la germinación in vitro de esta especie bajo distintas condiciones de luz. En este sentido, la presente investigación se planteó como objetivo, describir la germinación, morfología de las plántulas de alpiste (Phalaris canariensis), establecer la relación coleoptilo: porción expuesta de la primera hoja, presencia de raíces adventicias, contenido de clorofila en plántulas y respuesta a la calidad de luz.
MATERIAL Y MÉTODOS
La investigación se realizó en el Laboratorio Auxiliar de la cátedra de Fisiología Vegetal de la Facultad de Agronomía de la Universidad Central de Venezuela. Se utilizó un total de 1 350 semillas de alpiste (Phalaris canariensis L.) compradas en un establecimiento comercial que fue seleccionado debido a que presentó el mayor porcentaje de germinación que otros lotes de semillas evaluados previamente.
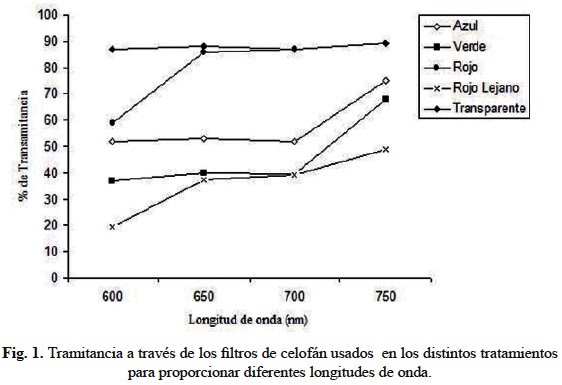
Las semillas se distribuyeron al azar en cápsulas de Petri, se colocaron 25 semillas por cápsula para conformar nueve tratamientos con seis repeticiones cada uno bajo un diseño experimental completamente aleatorio. En cada cápsula se colocó papel absorbente en el fondo y se esterilizó en estufa a 60°C durante 24 horas. Una vez culminada la esterilización se colocaron 25 semillas por cápsula agregando 10mL de agua destilada en cada una, allí permanecieron durante dos horas previo a la aplicación de los tratamientos a una temperatura ambiente de 27°C. Una vez culminado este tiempo para imbibición, se procedió a aplicar los tratamientos que se indican a continuación, en los cuales la irradianza provino de una lámpara fluorescente de bombillo circular TLE 32W/54 Philips, colocada a 50 cm sobre las cápsulas de Petri. También se usó celofán transparente, azul, verde y rojo. Para el caso del rojo lejano, éste fue obtenido mediante cuatro bombillos de 100 W iluminando una capa de papel celofán azul colocado entre dos capas de celofán rojo (Ross, 1978). Para determinar las características ópticas del celofán, que en lo sucesivo se denominará filtro, se examinaron muestras de cada color con un Spectronic 20, determinando el barrido de tramitancia a 600, 650, 700 y 750 nm, definida como absorbancia = -log T o fracción de luz que pasa a través de la muestra. Se incluyó una muestra de celofán transparente.
De este modo, se aplicaron nueve tratamientos, que se describen en el cuadro 1. A los siete días se determinó el porcentaje de germinación por cápsula y se midió la longitud de la radícula, coleoptilo y hoja. Se estableció la relación coleoptilo: porción expuesta de la primera hoja, dividiendo la longitud del coleoptilo entre la longitud de la porción visible de la primera hoja y se observaron aspectos morfológicos resaltantes en las plántulas tales como presencia de raíces adventicias.
Para la determinación de clorofila 100 semillas de alpiste se distribuyeron en cuatro contenedores de vidrio sobre papel de filtro humedecido con agua destilada. Seguidamente, se mantuvieron en condiciones de umbráculo durante siete días. Transcurrido este tiempo se procedió a separar los contenedores para establecer dos tratamientos: T1, condiciones de luz natural, y T2, condiciones de oscuridad, con dos contenedores con 25 plántulas en cada uno. Al cabo de una semana bajo estas condiciones, se procedió a evaluar la morfología de las plántulas y determinar el contenido de clorofila. Para ello, se tomaron tres muestras de hojas de alpiste al azar, conformadas por cinco hojas cada una por cada tratamiento, para un total de seis muestras. Se utilizó el método colorimétrico utilizando un espectrofotómetro marca Spectronic 20 calibrado con un blanco de 6 mL de acetona al 80%. Se determinó la tramitancia (%T) a 645 y 663 nm y finalmente la concentración de clorofila (mg mL-1) calculada mediante la relación: concentración de clorofila = 0.0202 * A(645 nm) + 0.00802* A (663 nm).
Una vez obtenidos los resultados, se realizó el análisis de varianza (ANOVA), utilizando el paquete estadístico Statistix Versión 7.0 (Statistix, 1985), se determinó la media y la desviación estándar para cada tratamiento, para un nivel de significación de 5%.
RESULTADOS
En la figura 1 se muestra el porcentaje de tramitancia determinado a 600, 650, 700 y 750 nm. Se registró 90% de tramitancia para los filtros rojo y transparente a 650, 700 y 750 nm, 40-70% para los filtros azul y verde. El menor porcentaje de tramitancia correspondió a la irradiación con luz en el rojo lejano (20-40%).
Respuesta de la germinación de semillas in vitro
La semillas de alpiste (P canariensis L.) "in vitro" mostraron una respuesta fotoblástica negativa, es decir que no requieren luz para germinar (Salisbury y Ross, 1992). La figura 2, muestra el porcentaje de germinación en los tratamientos aplicados. El mayor porcentaje se obtuvo en el tratamiento T5 (condiciones de oscuridad), seguido por T1 y T2 (luz natural y luz roja), existiendo diferencias estadísticamente significativas entre ellos, por lo que se puede afirmar que las semillas exhiben una respuesta fotoblástica negativa. El menor porcentaje de germinación se obtuvo en semillas bajo T3 (luz roja-roja lejana), T6 (filtro transparente-luz), T8 (filtro verde-luz) y T9 (filtro rojo-luz). Así mismo, los tratamientos con luz roja-rojo lejano-luz roja (T4) y filtro azul (T7) se comportaron iguales, con porcentaje de germinación de 24 y 27%, respectivamente, siendo éstos superiores a los grupos de tratamientos anteriores. A este respecto, se ha señalado que la luz roja promueve la germinación en semillas de Coriandrum sativum L. (Peroza et al., 2007) coincidiendo esta respuesta con la mostrada para alpiste y, por el contrario, inhibida por radiaciones en la faja del infrarrojo (Peroza et al., 2007), tal como se evidenció en T3 cuando las semillas fueron irradiadas con luz roja y posteriormente rojo lejano y en T9 con filtro rojo, que revierte el fitocromo a su forma inactiva, por lo que la cantidad de semillas que germinaron fue menor en estos tratamientos, pero sin diferencias estadísticamente significativas entre ellos (fig. 2). Así mismo, los resultados muestran que bajo filtro transparente (T6), al igual que bajo filtro de color verde (T8) se generó un efecto inhibitorio de la germinación, el primero no señalado en la literatura con anterioridad, y el segundo ampliamente señalado en la literatura (Folta y Maruhnich, 2007). Estadísticamente, de acuerdo al porcentaje de germinación de semillas en todos los tratamientos, se conformaron tres grupos con respuesta similar: el primero con T5, el segundo grupo con T1, T2, el tercer grupo conformado por T3, T4, T6, T7, T8 y T9 (fig. 2).

La figura 3 muestra que los tratamientos en los cuales la radícula alcanzó mayor longitud fueron T2 (7.6 cm), T3 (5.7 cm), T4 (6.8 cm) y T5 (7.8 cm). La radícula presentó entre 1.8 y 3.07cm en los tratamientos T1 (3.07 cm), T6 (3.5 cm), T7 (2 cm) y T9 (1.8 cm), que no mostraron diferencias significativas entre ellos. El tratamiento con filtro verde (T8) difirió estadísticamente del resto de los tratamientos, ya que en éste las semillas exhibieron una radícula de 0.8 cm de longitud.

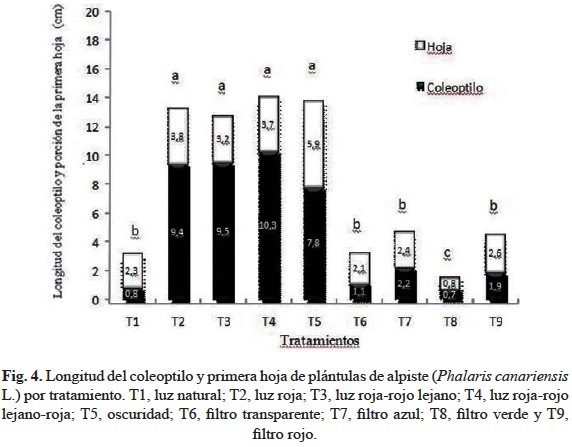
En la figura 4 se realiza una comparación de la longitud promedio del coleoptilo y primera hoja de las plántulas en los distintos tratamientos. En esta figura, la porción mostrada como hoja es la que sobresalió por dentro del coleoptilo, por lo que la longitud total de la primera hoja corresponde a la sumatoria de la longitud del coleoptilo más la porción señalada en la figura 4 como hoja. La proporción longitud del coleoptilo: porción expuesta de la primera hoja varió entre tratamientos, siendo de 3:1 en los tratamientos T2, T3, T4, T5; de 2:1 en T1, T6, T9 y de 1:1 en T7 y T8.
Aspectos morfológicos de las plántulas en cada tratamiento:
Cuando las plántulas fueron expuestas a luz natural (T1) la longitud de la primera hoja fue mayor (3 cm) observándose ésta de color verde intenso y la radícula de 3 cm en relación con el coleoptilo que presentó 0.8 cm de largo. En oscuridad (T5) la longitud de la primera hoja fue de 13.7 cm observándose ésta clorótica, la radícula alcanzó 7.8 cm y el coleoptilo fue inhibido (0.8 cm) (figs. 3 y 4).
Bajo luz roja (T2) se observó claramente el patrón de etiolación y coleoptilo de color blanquecino-rojizo con una longitud de 9.4cm, y la primera hoja clorótica de 13.2 cm (fig. 4). Así mismo en T2, la radícula fue de 7 cm de largo (fig. 3) y presentó desarrollo de raíces adventicias muy pequeñas y delgadas, ubicadas a pocos centímetros por encima de la base, indicando que bajo luz roja (T2) el crecimiento de la radícula fue estimulado.
En el tratamiento bajo luz roja-luz roja lejana (T3), las plántulas mostraron etiolación con valores de longitud de coleoptilo y primera hoja similares a los obtenidos bajo luz roja (en T2) 9.5 y 12.7 cm respectivamente (fig. 4). Adicionalmente, en T3 se observó una coloración blanquecina-rojiza del coleoptilo y clorosis de la primera hoja alargada y ausencia de raíces adventicias emergiendo de la radícula.
En el tratamiento bajo luz roja-rojo lejano-luz roja (T4), las plantas presentaron mayor etiolación que el resto de los tratamientos, siendo la longitud de la primera hoja de 14 cm de largo y su aspecto clorótico. El coleoptilo con coloración rojiza-blanquecina se alargó hasta 10.3 cm (fig. 4). En T4 la radícula de 7 cm de largo (fig. 3) fue muy delgada y presentó raíces adventicias en la base, denotando que este tratamiento favoreció la rizogénesis.
En oscuridad (T5), las plántulas mostraron coleoptilos etiolados de 7.8cm de largo y la primera hoja clorótica, de 13.7 cm (fig. 4). Bajo estas condiciones las plántulas presentaron una radícula de 7.8 cm (fig. 3) casi transparente con presencia de raíces adventicias en su base. Los resultados muestran que en T5 tanto el alargamiento de la radícula como su diferenciación fueron estimulados.
Cuando se colocó filtro transparente (T6) no se observaron plántulas etioladas, las hojas fueron de color verde intenso con 3.2 cm de largo y la relación coleptoilo: la porción expuesta de la primera hoja fue de 2:1 (fig. 4). En T6 las plántulas presentaron raíces adventicias en la parte media de la radícula que midió 3.5 cm (fig. 3).
Bajo filtro azul (T7) no se observaron plántulas etioladas. Las hojas fueron de color verde intenso con una longitud de 4.6 cm y la relación coleptilo: la porción expuesta de la primera hoja fue de 1:1 (fig. 4). En T7 se observó la formación de raíces adventicias en parte media y base de la radícula.
Cuando se utilizó filtro verde (T8) el crecimiento de las plántulas fue el más reducido en comparación con el resto de los tratamientos, el efecto inhibitorio se observó a nivel del coleoptilo, la primera hoja y la radícula. El coleoptilo fue de 0.7 cm y la primera hoja de 1.5 cm (fig. 4). Así mismo, se evidenció que el tratamiento con filtro verde (T8) ejerció el mayor efecto inhibitorio sobre el crecimiento de la radícula que en este caso fue de 0.8 cm de largo.
Bajo filtro rojo (T9) no se observaron plántulas etioladas, el coleoptilo midió 1.9 cm y la primera hoja 4.5 cm de largo (fig. 4) con proporción 2:1, respectivamente. En el tratamiento T9 se notó el efecto inhibitorio del filtro rojo sobre del crecimiento de la radícula, en comparación con el transparente con 1.8 cm de largo (fig. 3).
El contenido de clorofila fue mayor en plántulas creciendo en luz que en oscuridad (6.42 x 10-3 y 3.49 x 10-3 mg mL-1, respectivamente), tal como era de esperarse.
DISCUSIÓN
El efecto inhibitorio del filtro del rojo lejano sobre la germinación no dependió de la cantidad de luz aplicada sino de su calidad, debido a que el efecto es observable aun cuando la tramitancia bajo este filtro fue inferior a la de los demás filtros utilizado, así lo destaca Chory (1997) en sus estudios de respuestas moduladas por la luz en las plantas.
Por otra parte, la fuente de luz roja, combinaciones de luz roja con rojo lejano y la oscuridad, parecieron no tener efecto estimulante sobre la emergencia de la radícula, lo cual se evidencia en los bajos porcentajes de germinación. Sin embargo, después de su emergencia, la radícula respondió positivamente a irradiaciones con luz roja y combinaciones de luz roja con rojo lejano, las cuales estimulan su alargamiento. A este respecto Folta y Maruhnich (2007) refieren que las plántulas que se desarrollan bajo la influencia de luz roja, poseen mayor cantidad de tejido vegetativo que las que crecen bajo luz natural.
Por el contrario, el crecimiento de la radícula respondió negativamente a la presencia de luz natural y filtro verde. A este respecto, Folta y Maruhnich (2007) reportaron que la luz natural tiene en su espectro componentes en el rango del rojo y del azul, las cuales son eficientes en promover el desarrollo y la fotosíntesis, no obstante, al suministrar luz natural, no se está exento de suministrar su componente en el verde. En este sentido, los citados investigadores señalan que el sistema de sensores para luz verde producen ajustes en el desarrollo y crecimiento en concordancia con los sensores para luz roja y azul.
Así, el coleoptilo de alpiste fue fuertemente inhibido bajo luz natural y filtro verde, seguido por el azul, que son los colores que corresponden a los colores de la clorofila, en los cuales el máximo de absorción del pigmento es menor. Respuestas similares han sido reportadas por expertos en los estudios de fotomorfogénesis (Folta, 2004). La relación entre ellos: longitud del coleotilo: porción expuesta de la primera hoja, varió significativamente dependiendo de la calidad de luz, mostrando la mayor inhibición bajo irradiación con el verde.
En esta investigación, condiciones de oscuridad generaron igual respuesta en plántulas de alpiste que bajo combinaciones de luz roja que se observaron como alargamiento del coleoptilo debido a etiolación, por lo que podría inferirse que estos tratamientos inactivaron el fitocromo.
El coleoptilo y la primera hoja formada mostraron diferencias en sus características morfológicas en los tratamientos. En un trabajo realizado por Parks et al. (2001) se afirma que el hipocotilo en crecimiento, es un órgano dinámico que ajusta su tasa de crecimiento a la condición ambiental que prevalece. En el caso de la presente investigación y en virtud de las respuestas obtenidas en alpiste, podríamos afirmar que el coleoptilo y la primera hoja son estructuras en crecimiento dinámico en las cuales se observaron respuestas contrastantes que permitieron agrupar a los tratamientos como base de la presente discusión.
CONCLUSIONES
El alpiste (Phalaris canariensis) posee características que permiten considerarla como una especie promisoria de gran utilidad en la investigación biológica aplicada, debido principalmente a que germina en corto tiempo, no requiere luz para germinar, la obtención de una gran cantidad de plántulas in vitro es fácil y rápida. Además, esta especie exhibe una respuesta diferencial a la calidad de luz que se puede interpretar con facilidad al registrar la relación longitud del coleotilo; porción expuesta de la primera hoja y al ser una planta de fácil muestreo y manejo en el laboratorio puede proveer material vegetal con características deseables para estudios biológicos.
LITERATURA CITADA
Cataús, G.L., 1997. "Las gramíneas (Poaceae) de Cuba. I". Fontiqueria, 46: 15-17. [ Links ]
Chory, J., 1997. "Light modulation of vegetative development". The Plant Cell., 9: 1225-1234. [ Links ]
Enamorado, M., A. Tapia, S. Gouiric, G. E. Feresin, 2004. "Actividad biológica de extractos obtenidos a partir de cultivo de hongos filamentosos aislados en la provincia de San Juan, Argentina". Reunión del XXXII Congreso Argentino de Horticultura, 28(3): 2004-2055. [ Links ]
Folta, K.M., 2004. "Green light stimulates early stem elongation, antagonizing light-mediated growth inhibition". Plant Physiol., 135: 1407-1416. [ Links ]
Folta, K.M., S.A. Maruhnich, 2007. "Green light: a signal to slow down or stop". J. Exp. Bot., 58(12): 3099-3111. [ Links ]
Paredes, S.D., A. Korkmaz, L.C. Manchester, D.Y, Tan., R.J. Reiter, 2009. "Phytome-latonin: a review". Journal of Experimental Botany, 60(1): 57-69. [ Links ]
Parks, B. M., K.M. Folta, E.P. Spalding. 2001. "Photocontrol of stem growth". Current Opinion in Plant Biology, 4(36): 436-440. [ Links ]
Peroza, J., O. Guillén, R. Santos, Z. Viloria, 2007. Incidencia de diferentes intensidades de radiación solar en la germinación del cilantro (Coriandrum sativum L.). XVII Con. Ven. Bot., FE-09. pp: 604-606. [ Links ]
Ross, 1978. Plant Physiology Laboratory Manual. University of California, Berkely. USA. 212 pp. [ Links ]
Salisbury, I.B.C., y W. Ross, 1992. Plant Physiology. Wadswoth, Publishing, California. USA. 682 pp. [ Links ]
SENASA, 2011. "Situación de la Producción Orgánica en la Argentina durante el año 2011". Servicio Nacional de Sanidad y Calidad Agroalimentaria y Agropecuaria. Informe estadístico SENASA 2011. Argentina. 37 pp. [ Links ]
Truol, G., T. Usugi, J. Hirao, J. Arneodo, P. Pecci, y I. Laguna, 2001. "Transmisión experimental de virus del mal de Río Cuarto por Delphacodes kuscheli". Rev. Fitopatol. Bras., 26(1): 39-44. [ Links ]

