










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPolibotánica
versión impresa ISSN 1405-2768
Polibotánica no.35 México feb. 2013
Producción de hojarasca y retorno potencial de nutrientes en tres sitios del estado de Nuevo León, México
Litterfall and potential nutrient return in three sites of the state of Nuevo Leon, Mexico
Juan Manuel López-Hernández1, Humberto González-Rodríguez1, Roque Gonzalo Ramírez-Lozano2, Israel Cantú-Silva1, Marco Vinicio Gómez-Meza3, Marisela Pando-Moreno1 y Andrés Eduardo Estrada-Castillón1
1 Facultad de Ciencias Forestales, Carr. Nac. Núm. 85, km 145 Linares, Nuevo León 67700, México. Correo electrónico: humberto.gonzalezrd@uanl.edu.mx.
2 Facultad de Ciencias Biológicas, San Nicolás de los Garza, Nuevo León, México.
3 Facultad de Economía, Monterrey, Nuevo León, México. Universidad Autonoma de Nuevo León.
Recibido: 1 mayo 2012.
Aceptado: 19 diciembre 2012.
Resumen
Los objetivos de este estudio fueron estimar y cuantificar la dinámica mensual (enero a diciembre 2009) de la deposición de la hojarasca y macro (Ca, K, Mg, N y P) y microminerales (Cu, Fe, Mn, y Zn) de las hojas de la hojarasca en los municipios Los Ramones, China y Linares, del estado de Nuevo León, México. La vegetación está caracterizada como matorral espinoso tamaulipeco. La deposición anual total de hojarasca fue de 321.5 (Los Ramones), 431.6 (China) y 462.9 (Linares) g/m2/año. Las hojas representaron el mayor componente (154.4 a 304.5 g/m2/ año) del total de hojarasca depositada. La deposición de ramas varió de 48.25 a 75.16 g/m2/año y la de estructuras reproductivas de 58.4 a 98.9 g/m2/año. La de otros componentes (material vegetal no identificado, cuerpos y heces de insecto) varió de 19.9 a 29.2 g/m2/año. La deposición anual de Ca en los tres sitios fluctuó de 3.4 a 8.1; K de 1.4 a 3.2; Mg de 0.76 a 1.6; N de 3.0 a 5.5 y P de 0.05 a 0.11 g/m2/año. El Cu fluctuó de 0.84 a 2.8; Fe de 14.7 a 21.7; Mn de 5.5 a 11.8 y Zn de 3.3 a 5.2 mg/m2/año. En invierno se depositó la mayor cantidad de minerales. Además, en Linares se depositó la mayor cantidad de minerales y en Los Ramones la menor. Hubo diferencias espaciales y temporales en cantidad de hojarasca colectada y retorno de minerales, mas no así en la calidad foliar entre los sitios de estudio.
Palabras clave: producción de hojarasca, minerales en las hojas de la hojarasca, matorral espinoso tamaulipeco, noreste de México.
Abstract
The aims of the study were to estimate and quantify the monthly dynamics (January to December 2009) of litterfall deposition and macro (Ca, K, Mg, N y P) and microminerals (Cu, Fe, Mn, y Zn) of litterfall leaves in the counties Los Ramones, China y Linares, of Nuevo León State, México. The plant community is named as Tamaulipan Thornscrub. Annual deposition was 321.5 (Los Ramones), 431.6 (China) and 462.9 (Linares) g/m2/year. Leaves represented the highest component (154.4 to 304.5 g/m2/year) of total litterfall production. Branches varied from 48.25 to 75.16 g/m2/year. Reproductive structures from 58.4 to 98.9 g/m2/year and other components (unidentified material, bodies and insect feces) ranked from 19.9 to 29.2 g/m2/year. Annual deposition of Ca in the three sites varied from 3.4 to 8.1; K from 1.4 to 3.2; Mg from 0.76 to 1.6; N from 3.0 to 5.5 and P from 0.05 to 0.11 g/m2/year. Cu ranked from 0.84 to 2.8; Fe from 14.7 to 21.7; Mn from 5.5 to 11.8 and Zn from 3.3 to 5.2 mg/m2/year. In winter mineral deposition was higher. Moreover, the highest mineral deposition occurred in Linares and Los Ramones was the lowest. There were spatio-temporal variations in quantity of literfall collected and return of minerals, but not so in leaf quality between study sites.
Key words: litterfall, minerals, Tamaulipan Thornscrub, Northeastern Mexico.
INTRODUCCIÓN
La producción de hojarasca y su descomposición son procesos fundamentales en el ciclo de nutrientes, ya que representa la principal transferencia de materia orgánica y nutrientes desde la parte aérea a la superficie del suelo (Isaac y Nair, 2006). Esta vía, además de la precipitación directa y flujos corticales (Cantú y González, 2001), es la principal fuente de fertilización natural. Más de la mitad de la absorción anual de nutrimentos en los bosques es debido a la reincorporación de hojarasca al suelo y el subsecuente reciclaje de estos nutrientes, y representa la principal fuente de minerales disponibles (Del Valle-Arango, 2003).
La hojarasca es una medida de la producción primaria neta del ecosistema y está fuertemente correlacionada con el incremento de la biomasa, la densidad de árboles y la apertura del dosel; sin embargo, es afectada por variables ambientales como precipitación, temperatura, elevación, fertilidad de los suelos y la evapotranspiración potencial (Oelbermann y Gordon, 2000). Los aportes de hojarasca varían ampliamente entre ecosistemas forestales en términos de calidad y cantidad. La calidad de la materia orgánica del suelo es de gran importancia para la mayoría de los procesos funcionales que se registran en el suelo de los ecosistemas forestales (Santa Regina et al., 2005). Los matorrales subtropicales de las planicies semiáridas de la región noreste de México, constituye la vegetación natural que abarca aproximadamente el 80% de la superficie de los estados de Nuevo León y Tamaulipas, en las cuales predominan las asociaciones caracterizadas por estratos arbustivos y arbóreos altos o medianos (3-6 m), zacatales, nopales, siendo las más comunes las especies espinosas con hojas compuestas. Este tipo de vegetación, denominada matorral espinoso tamaulipeco (MET), constituida en su gran mayoría por especies caducifolias que pierden sus hojas en periodos de sequía y algunas perennifolias (Northup et al., 1996), se distinguen por tener un amplio rango taxonómico diferente, dinámica de crecimiento, diversidad en desarrollo fenológico y longevidad foliar (González et al., 2011).
El uso eficiente de los minerales por las plantas [(g hojas m2/año)/(g mineral en hojas m2/año)] provenientes de la hojarasca está relacionado al uso del mineral para producir nueva biomasa, como una estrategia competitiva para las plantas que crecen y se desarrollan en suelos de baja fertilidad, a la capacidad funcional del ecosistema para producir hojarasca (producción primaria neta), y a las características estructurales del ecosistema tales como edad, cobertura, diversidad y hábitos de crecimiento (Vitousek, 1982).
En particular, se puede considerar que la biomasa arbustiva constituye una importante fuente forrajera, energética, de estantería, construcciones rurales, alimentarias y medicinales (Reid et al., 1990) entre otros beneficios. Debido a su valor, en esta comunidad vegetal se han realizados diversos estudios científicos sobre diversidad (Rodríguez, 1994; Sariñana, 1999), sistemas de muestreo (Villalón et al., 1991) y en reciclaje de nutrientes (Ramírez-Lozano et al., 2010; Rodríguez et al., 2011). A pesar de la diversidad de estudios ecológicos y biológicos realizados en el MET del noreste de México, y particularmente en el estado de Nuevo León, no se ha documentado en si la deposición y ciclaje de minerales vía hojarasca en este tipo de ecosistemas. Por tanto, los objetivos de este estudio fueron evaluar el ciclo anual de producción de hojarasca y flujo de nutrientes a través del componente foliar de la hojarasca en tres sitios del noreste de México.
MATERIAL Y MÉTODOS
Área de estudio
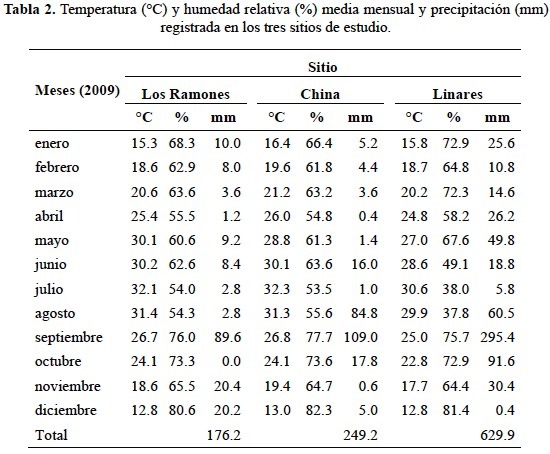
El estudio se llevó a cabo en tres sitios sin disturbios antropogénicos del MET en el estado de Nuevo León, noreste de México. El sitio 1 (Los Ramones: Rancho El Abuelo) se encuentra ubicado en el municipio de Los Ramones (25° 40 N; 99° 27 O, 200 m.s.n.m.). El sitio 2 (China; Rancho Zaragoza) se encuentra en el municipio de China (25° 31 N; 99° 16 O, 200 m.s.n.m.). El sitio 3 (Linares) se sitúa en la Estación Experimental de la Facultad de Ciencias Forestales de la Universidad Autónoma de Nuevo León (24° 47 N; 99° 32 O, 350 m.s.n.m.), 8 km al sur del municipio de Linares. Las distancias entre sitios son: de Los Ramones a China es de aproximadamente 54 km, de China a Linares es de 131 km y de Los Ramones a Linares es de 185 km. El clima de la región es subtropical y semiárido con verano cálido, con lluvias entre los meses de abril a noviembre y con la presencia de un periodo de sequía intraestival, presentándose temperaturas de hasta 45°C durante el verano, con temperatura promedio entre 14.7° y 22.3°C. La precipitación promedio anual es de 805 mm con una distribución bimodal (González et al., 2004). Los suelos dominantes de estos tres sitios son vertisoles profundos (0.80 a 1.50 m aproximadamente), de color gris oscuro, limo-arcillosos, con Montmorillonita, que se contraen y se expanden notablemente en respuesta a los cambios en el contenido de humedad del suelo.
Vegetación del área de estudio
El principal tipo de vegetación que se encuentra en los sitios de estudio (Los Ramones, China y Linares), es conocida como matorral espinoso tamaulipeco (MET), caracterizándose por ser una formación arbustiva y subarbóreas, con elementos florísticos dominantes de 4 a 6 m de alto, perennes subespinosos, con hojas pequeñas y caducifolias. Citándose entre las especies más representativas de este tipo de vegetación a: mezquite, ébano, chaparro prieto, chaparro amargoso, granjeno, palo verde, panalero, anacahuita, cenizo, guayacán, tasajillo, nopal, colima, coma, barreta, zapotillo, destacando entre todos éstos la palma china (Yucca filifera) alcanzando ésta una altura de más de 10 m, todas éstas presentes en los sitios de estudios. En la tabla 1 se muestran algunas especies identificadas por sitio de estudio de acuerdo a su fenología, número de individuos, altura media y cobertura. Las especies caducifolias pierden sus hojas en otoño o a inicios del invierno, mientras que la caída de hojas de las perennifolias se origina de manera constante a lo largo de todo el año, aunque la mayor abscisión de hojas ocurre en la época estival (verano-otoño) (Moro, 1992).
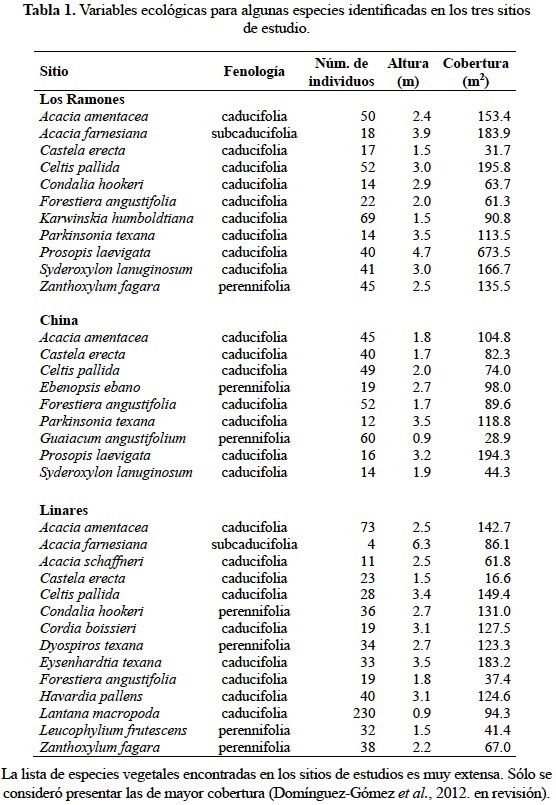
Colecta de hojarasca
A principio del año 2009, en cada sitio de estudio, se establecieron diez canastas o colectores de hojarasca (1.0 m2), consistentes de un marco de madera biselado cuyo fondo se cubrió con malla plástica fina (1.0 mm2) con la finalidad de que no se acumule el agua en los periodos de lluvia. La ubicación de las canastas se realizó al azar en un área de aproximadamente 2 500 m2. Cada canasta fue colocada aproximadamente a 0.50 m sobre el nivel del suelo para interceptar la hojarasca. La producción de hojarasca de cada canasta se colectó y colocó en bolsas de papel previamente etiquetadas con la leyenda sitio, fecha y número de repetición. Con el propósito de prevenir una subestimación y minimizar la pérdida de hojarasca a través del tiempo, la recolecta de hojarasca de cada canasta se realizó a intervalos de 15 días durante los meses enero a diciembre de 2009, no obstante, las dos recolectas de cada mes se mezclaron.
Análisis de laboratorio
El contenido de hojarasca se clasificó manualmente en hojas, estructuras reproductivas (flores, frutos y semillas), ramas, y otros componentes (tejidos no identificados, estructuras finas, corteza, cuerpos de insectos, heces, entre otros). No se cuantificó la pérdida de peso de la muestra de la hojarasca que pudo haber ocurrido entre fechas de muestreo o la cantidad de hojarasca depositada dentro o fuera de las canastas por la acción del viento. Una vez separado cada componente de la hojarasca, las muestras se secaron en estufa a 65°C por espacio de 72 h hasta peso constante. Sumando los muestreos realizados por mes, repetición y sitio de estudio, se determinó la deposición anual total de hojarasca y sus componentes. Una vez cuantificada la hojarasca, las hojas fueron molidas utilizando un molino Thomas Willey (Thomas Scientific Apparatus, Modelo 3383) usando una malla del núm. 60 (1 mm × 1 mm). El material molido fue recolectado en bolsas tipo ziploc previamente etiquetadas por fecha, repetición y sitio de muestreo para su posterior análisis químico. Se consideraron sólo las hojas por ser el componente principal de la hojarasca y estar presente a lo largo de todo el periodo de estudio.
Análisis químico
De cada canasta (repetición) proveniente de cada sitio y mes de colecta se usó 1.0 g para determinar la concentración de minerales. Primeramente, fueron incineradas en una mufla a 550°C durante 5 h. Las cenizas obtenidas fueron digeridas en una solución de HCl y HNO3 utilizando la técnica de digestión húmeda (Cherney, 2000). Las concentraciones de Ca (óxido nitroso/llama de acetileno), K, Mg, Cu, Mn, Fe y Zn (aire/llama de acetileno) se determinaron por espectrofotometría de absorción atómica mediante el uso de un espectrofotómetro marca Varian, modelo SpectrAA-200, mientras que el P se cuantificó utilizando un espectrofotómetro Perkin-Elmer (modelo Lamda 1A) a 880 nm (AOAC, 1997). El contenido total de nitrógeno se realizó por el método micro-Kjeldahl (AOAC, 1997). La deposición de minerales se cuantificó multiplicando la producción de hojas de la hojarasca por la concentración de cada mineral. Los valores mensuales acumulados en cada sitio se utilizan como una estimación de la deposición anual de minerales. La deposición anual de Ca, K, Mg, N y P fueron utilizados para determinar, en cada sitio, la eficiencia en el uso de macrominerales (EUN) de la hojarasca vía hojas que se define como la relación de la masa anual de hojas depositadas a la deposición anual de macrominerales (Vitousek, 1982).
Variables ambientales
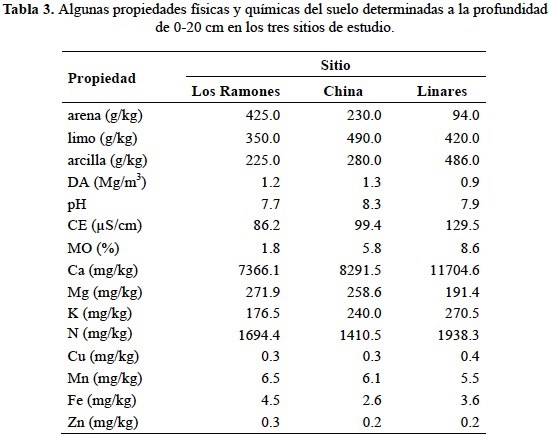
Variables ambientales tales como temperatura ambiente (°C) y humedad relativa (%) fueron obtenidos (base horaria) en cada sitio de estudio usando sensores automatizados tipo HOBO (Familia H8). La cantidad diaria de precipitación pluvial (mm) que se registró durante el periodo experimental fue cuantificada en cada sitio de estudio usando un pluviómetro automatizado marca HOBO (tabla 2). Adicionalmente, se tomaron muestras de suelo en cada sitio de estudio a una profundidad de 0-20 cm para caracterizar algunas propiedades físicas tales como densidad (mg/m3), g/kg de arena, de arcilla y de limo. Entre las propiedades químicas caracterizadas en cada sitio fueron el pH, CE (μS/cm), contenido de materia orgánica (%), y concentración de macro y micronutrientes. Las determinaciones químicas y físicas del suelo se realizó al inicio del año y para ello se consideraron cuatro repeticiones en cada sitio de estudio (tabla 3).
Análisis estadísticos
Los datos de producción de la hojarasca, así como los de concentración de nutrientes de las hojas fueron sometidos a un análisis de varianza utilizando un diseño completamente al azar (Steel y Torrie, 1980). Para probar los supuestos de normalidad y homogeneidad de varianzas de cada componente de la hojarasca y concentración de nutrientes, los datos se sometieron a las pruebas estadísticas de Kolmogorov-Smirnov, Shapiro Wilk y Levene (Steel y Torrie, 1980). Los resultados demostraron que la mayoría de los datos no se distribuyeron normalmente y, en la mayoría de los meses de muestreo, el análisis de varianza no mostró los supuestos de igualdad de varianzas, por lo que se utilizó la prueba no paramétrica de Kruskal-Wallis (Ott, 1993) para detectar diferencias significativas entre los sitios en cada mes de muestreo. Todos los análisis estadísticos se realizaron usando el paquete computacional Statistical Package for the Social Sciences (SPSS) versión 17.0 para Windows, SPSS Inc, Chicago, IL, USA.
RESULTADOS
Producción de hojarasca
La producciones mensuales de hojarasca (Fig. 1a), hojas (Fig. 1b), estructuras reproductivas (Fig. 1c), ramas (Fig. 1d) y otros componentes (Fig. 1e) mostraron variaciones significativas entre los sitios de estudio sólo en ciertos meses como lo demuestran los análisis estadísticos indicados en las figuras respectivas. La producción total de hojarasca fue mayor en Linares variando de 18.7 g/m2 (noviembre) a 68.3 g/m2 (abril), seguido de Los Ramones fluctuando de 6,8 (diciembre) a 62.8 g/m2 (octubre) y China con un rango de 16 (diciembre) a 53 (junio). La deposición de hojas también fue mayor en Linares con un rango de 13 g/m2 (noviembre) a 48 g/m2 (abril), seguido de China que fue de 12.1 (diciembre) a 38.3 (febrero) y Linares que fluctuó de 3.5 (diciembre) a 27.7 (enero). La producción de estructuras reproductivas fue mayor en China con un rango de 0.12 g/ m2 (diciembre) a 31 g/m2 (junio), seguido de Los Ramones que varió de 0.27 (diciembre) a 30 (octubre) y en Linares que fue de 0.8 (diciembre) a 19 (octubre). La producción de ramas fue mayor en Linares y fluctuó de 2.2 g/m2 (diciembre) a 11.2 g/m2 (febrero), seguido de China con 3 (junio) y 10 (enero) y Los Ramones que fue de 2.3 (diciembre) a 8 (abril). La producción del componente otros en China y Linares se registraron las deposiciones mínimas en noviembre (0.2 g/m2) y las máximas en septiembre (11.5 g/m2); en Los Ramones varió de 0.2 (noviembre) a 9.5 (octubre). En lo que respecta a las producciones anuales de hojarasca, Linares presentó la mayor producción total anual (462.9 g/m2/año) de hojarasca, seguido por China (431.6) y Los Ramones (321.5) (tabla 4).

Concentración de minerales en las hojas
Las concentraciones de macro (Figs. 2a,b,c,d,e) y microminerales (Figs. 3a,b,c,d) de las hojas fueron significativamente diferentes entre sitios. El K fue el que más varió significativamente entre meses, en ocho de los doce estudiados, mientras que el N sólo en tres meses se detectaron diferencias significativas. De los microminerales, el Cu fue el que obtuvo mayores diferencias significativas en nueve fechas, en contraste el Zn varió significativamente en dos fechas. En general, durante los meses de verano y otoño los macrominerales en China, y los microminerales en Los Ramones, tuvieron concentraciones mayores comparados con otros meses y sitios.
Deposición de minerales
La producción potencial de macrominerales (Figs. 4a,b,c,d,e) fueron significativamente diferentes entre los sitios de estudio en la mayoría de los doce meses analizados. En general, durante el invierno (enero, febrero y marzo), donde se observó la mayor caída de hojas (Fig. 1b), la producción de todos los minerales fue mayor que en otros meses, con una tendencia a ser mayor en Linares. La producción anual de Ca para Los Ramones, China y Linares fue de 3.4, 4.7 y 8.1 g/m2, respectivamente. El aporte de K fue de 1.3, 3.2 y 3.1 g/m2, respectivamente. El de Mg fue de 0.8, 1.4 y 15 g/m2, respectivamente; mientras que el P tuvo una contribución potencial de 0.05, 0.1 y 1 g/m2, respectivamente. El aporte de N fue de 3.0, 4.9 y 55 g/m2, respectivamente (tabla 4). Independientemente del sitio, la contribución potencial anual de nutrientes a través de las hojas mostró el siguiente orden: Ca>N>K>Mg>P (tabla 4). El total de la producción anual de macros (Ca+K+Mg+N+P) para Los Ramones, China y Linares fue de 8.6, 14.4 y 18.4 g/m2, respectivamente.
El uso eficiente [(g hojas m2/año)/(g mineral en hojas m2/año)] de macrominerales en forma individual no mostró una tendencia clara entre los sitios (tabla 4). Para K y P el patrón fue el siguiente: Los Ramones>Linares>China, para Mg y N fue: Linares>Los Ramones>China, y para Ca fue: China>Los Ramones>Linares. Lo anterior pudiera explicarse a las diferencias entre especies y abundancia de plantas encontradas en cada sitio de estudio (González et al., 2010).
La producción potencial de microminerales (Figs. 5a,b,c,d) siguió la misma tendencia que los macrominerales; más de la mitad de los meses analizados presentaron diferencia significativa entre los sitios de estudio. En general, en enero, febrero y marzo, cuando se registró la mayor cantidad de hojas (Fig. 1b), la producción de todos los microminerales fue mayor que en otros meses, siendo mayores en Linares. El aporte potencial anual de Cu fue de 0,8, 1,2 y 2,8 mg/m2 para los sitios de Los Ramones, China y Linares, respectivamente; para Fe fue de 14.7, 15.4 y 21.7; para Mn fue de 5.5, 6.9 y 11.8 y para Zn fue de 3.3, 4.8 y 5.2. Independientemente del sitio, la producción potencial anual de micronutrientes fueron en el siguiente orden: Fe>Mn>Zn>Cu (tabla 4). El total de la producción anual (Cu+Mn+Fe+Zn) fue de 24.3, 28.4 y 41.4 mg/m2, en Los Ramones, China y Linares, respectivamente.
DISCUSIÓN
Aun cuando, la producción de hojarasca se produjo durante todo el periodo experimental, no se encontraron diferencias sobresalientes en las producción de los componentes de la hojarasca, pero sí se encontraron diferencias en la cantidad de entrada, la variabilidad y el orden de magnitud (Figs. 1a,b,c,d,e). Independientemente de los componentes de la hojarasca total o individual que se registraron en cada sitio, se encuentran dentro del rango de las producciones observadas previamente en estudios llevados a cabo en el MET (Rodríguez, 2010; González et al., 2011) y en el matorral desértico micrófilo donde se observaron producciones de 141.76 a 390.47 g/m2/año (Marroquín, 2011). Estudios realizados en ecosistemas forestales tropicales (Veneklaas, 1991; Sundarapandian y Swamy, 1999; Epstein et al., 2002; Del Valle-Arango, 2003) reportan producciones de hojarasca que varían de 240 a 760 g/m2/año. Estudios realizados en diferentes ecosistemas forestales de México (bosque de encino, bosque mesófilo de montaña y bosque perennifolio), la variación en la producción anual de hojarasca fluctuó de 430 a 1090 g/m2/año. (Lawrence, 2005; Rocha-Loredo y Ramírez, 2009).
En general, dentro de una misma región forestal y debido a que las hojas son retenidas durante menos tiempo en el árbol, los bosques caducifolios presentan una mayor producción de hojarasca que los de coníferas, debido a que la productividad es más alta. En la zona boreal los bosques de coníferas y caducifolios producen 32 y 264 g/m2/año, respectivamente. En la zona templada la producción es superior a la registrada en la zona boreal, ya que los bosques caducifolios aportan 540 g/m2/año y los de coníferas 438 g/m2/año. En el bosque de tipo mediterráneo se han registrado valores de producción de hojarasca de 384 g/m2/año (Lousier y Parkinson, 1976).
En este estudio, las hojas representaron el principal componente de la hojarasca. Asimismo, Santa Regina y Tarazona (2001a) reportaron que las acículas representaron el 50% del material de la hojarasca en plantaciones de Pinus sylvestris, el 55% en el caso de Quercus suber (Caritat et al., 2006) el 62% en Fagus sylvatica (Santa Regina y Tarazona, 2001b) y 80% en Quercus pyrenaica (Santa Regina, 2000). La cantidad de hojarasca que retorna al suelo y la concentración de nutrientes acumulados en ella determina la cantidad de cada uno de los elementos minerales que retornarán al suelo. Las hojas presentan una alta demanda de nutrientes con respecto al resto de los órganos del árbol; entre el 30 y el 70% del total de los nutrientes almacenados anualmente se encuentran en las hojas (Piatek y Allen, 2000).
La variación mensual en la producción de hojarasca y sus componentes ha sido reportado con anterioridad por Prause et al. (2003) quienes reportaron variaciones mensuales que van de 0.42 a 28.2 g/m2/año y su aporte al suelo está asociado al periodo del ciclo vegetativo con la sucesión de las etapas fenológicas características de cada especie vegetal. Asimismo, se ha observado que la producción de hojarasca y producción de minerales en los ecosistemas forestales está determinada por la edad de las plantas (Lawrence, 2005), la precipitación, la composición de especies, la disponibilidad de nutrientes en el suelo y por la etapa de sucesión (Yankelevich et al., 2006).
En este estudio, el patrón de la producción de los diferentes componentes de la hojarasca varió dentro y entre los sitios a lo largo del año. No obstante, la producción de estructuras reproductivas mostró dos incrementos importantes en su producción; en junio y octubre (Fig. 1c). El hecho de que la producción de los diferentes componentes de la hojarasca no sigan un patrón similar al del otros componentes, se pudiera deber a que la composición botánica de las comunidades vegetales es distinta entre los sitios (tabla 4).
En general, la producción de hojarasca es mayor en zonas húmedas y cálidas con suelos fértiles, mientras que disminuye en zonas secas y frías y con una baja disponibilidad de nutrientes (Kimmins, 1997). Sin embargo, la producción de hojarasca es muy dependiente de la variabilidad de las condiciones climáticas entre diferentes años, por lo que es difícil establecer un patrón general a escalas más pequeñas o en zonas con gran variabilidad climática, como en aquellas zonas climáticas del tipo de clima mediterráneo (Martínez-Alonso et al., 2007). La producción de hojarasca y las concentraciones de nutrientes que retorna al suelo por las hojas, son alteradas por las variaciones en la precipitación y temperatura. Por un lado, las precipitaciones pueden causar la caída prematura de las hojas debido al viento que acompaña a las tormentas; además, lavan nutrientes de las hojas verdes (Wood et al., 2005). En cambio, la sequía puede aumentar las concentraciones en nutrientes de la hojarasca, debido a una caída prematura de las hojas maduras en respuesta al estrés hídrico, lo que impediría una reabsorción de nutrientes; esto se debe sobre todo a que los nutrientes se pueden transportar hacia las raíces a través de películas de agua dentro del suelo y, por tanto, debe haber una película continua conectando los nutrientes con las raíces. La falta de agua en el suelo también disminuye la disponibilidad de los nutrientes reduciendo la actividad microbiana, la cual es responsable de la liberación de nitrógeno, fósforo y azufre de la materia orgánica del suelo (Imbert et al., 2004). En este sentido, Martínez-Alonso et al. (2007) señalan que el aumento de temperaturas y los periodos secos condicionan la dinámica y productividad de los ecosistemas. Esto se debe a que el contenido de humedad de suelo es muy pobre y reduce el crecimiento de las plantas, limitando la producción de alimento para la fauna que ahí habita, así como la lenta descomposición de la materia orgánica provocando con lo anterior la baja productividad de nutrientes.
La producción de minerales vía foliar de la hojarasca en el MET sólo se tiene lo registrado por Rodríguez (2010), donde reporta deposiciones que van de 0.23 (P) a 14.16 g/m2/año (Ca) y microminerales de 0.00244 (Cu) a 0.123 mg/m2/año (Fe). Valores muy similares a lo encontrado por González et al. (2011), quien reporta para el sitio de Linares producciones de macrominerales que van de 0.4(P) a 18.23 g/m2/año (Ca) y microminerales de 0.0272 (Cu) a 0.63 mg/m2/año (Fe), en el presente estudio se tienen producciones muy bajas para los macrominerales que van de 0.11 (P) a 8.05 g/m2/año (Ca) y producciones muy altas para microminerales de 2.79 (Cu) a 21.68 mg/m2/año (Fe), a pesar de que se trata del mismo sitio se ven muy marcadas las diferencias que pueden estar relacionadas, a las producciones de hojarasca, precipitación y temperatura y a la diversidad de plantas tanto caducifolias como perennifolias que pudieron estar presentes o ausentes en los diferentes años de estudio.
Estudios relacionados a este trabajo en diferentes tipos de ecosistemas, muestran valores similares [0.127 (P) a 5.26 g/m2/año (Ca)] como los reportados por Marroquín (2011) en dos comunidades de matorral desértico micrófilo del noreste de México, mientras que Yang et al. (2004) en un bosque natural subtropical en China reporta valores de 0.13 (P) a 4.6 g/m2/año (Ca). Sin embargo, cantidades más elevadas [0.25 (P) a 21.68 g/ m2/año (Ca)] fueron reportadas por Sánchez et al. (2008) en bosques forestales tropicales en Ecuador. Por su parte, Del Valle-Arango (2003) en un bosque pantanoso encontró valores de 0.5 (P) a 13.04 g/m2/año (Ca). A pesar de la diferencia en cuanto a estructura, densidad y cobertura, entre estos tipos de ecosistemas, la variación que hay en cuanto a la producción de minerales entre algunos ecosistemas es muy similar, no así en cuanto a cantidad y calidad de la hojarasca. Esto lo demuestra González et al. (2011), en un estudio realizado en diferentes tipos de ecosistemas, donde encontró que las mayores producciones de hojarasca se registraron en el ecotono de un bosque de pino-encino y el matorral espinoso tamaulipeco y la menor producción se registró en el bosque de pino-encino. En tanto, para las producciones de minerales, las mayores se registraron en el matorral espinoso tamaulipeco, mientras que el ecotono presentó las menores producciones.
La producción potencial de P fue muy baja. Gairola et al. (2009) en India y Andivia et al. (2009) en España también reportaron bajas producciones de P, 0.7 y 0.034 g/m2/año, respectivamente. En general, se observa que las cantidades de N y P que retornan al suelo a través de la hojarasca son mayores para especies caducifolias que para perennifolias. Imbert et al. (2004) documentan aportes anuales de N y P comprendidos entre 1.0 a 8.8 g/m2/año y 0.03 a 0.79 g/m2/año, respectivamente, para especies caducifolias, mientras que los aportes máximos de N y P en especies perennifolias son inferiores a 4.7 y 0.44 g/m2/año, respectivamente.
En el presente estudio, aun cuando la deposición de P fue baja, su uso eficiente fue relativamente mayor que Ca, K, Mg y N. Lo anterior también ha sido reportado en estudios realizados en diferentes ecosistemas forestales (Del Valle-Arango, 2003; Swany et al., 2004; Safou-Matondo et al., 2005;). Se ha argumentado que el P pudiera ser retranslocado a otras estructuras de las plantas antes de la senescencia de las hojas y esta reabsorción pudiera ser usada para la producción de nuevas estructuras vegetales o procesos fisiológicos. Frecuentemente se ha sugerido que las especies de hábitats pobres en nutrientes presentan una mayor eficiencia en la retranslocación (Vitousek, 1982). Otros autores han encontrado que en ambientes ricos en nutrientes la eficiencia en la reabsorción es muy baja. Así, Aerts (1996) apenas encontró diferencias significativas en la reabsorción de nutrientes entre especies caducifolias (en suelos más fértiles) y especies perennifolias (características de suelos más pobres en nutrientes), solamente observó que la eficiencia en la reabsorción era ligeramente mayor en las primeras (54 vs. 47% para el nitrógeno). Otros autores no encontraron relación entre la disponibilidad de nutrientes del suelo y la eficiencia en la retranslocación (Wright y Westoby, 2003; Rentería et al., 2005). En cambio, en otros estudios se ha encontrado que las plantas que crecen en hábitats fértiles son más eficientes en la reabsorción (Nambiar y Fife, 1987). Del Arco et al. (1991) establecen que el grado de reabsorción depende del periodo de abscisión de la hoja, de modo que las especies con una caída de la hoja más lenta en el tiempo presentan una menor eficiencia en la reabsorción de nutrientes debido probablemente a lo impredecible de la época de abscisión foliar. La abscisión gradual de hojas parece ser una adaptación al estrés hídrico en regiones con climas áridos o semiáridos. El estado nutricional de la planta y la humedad del suelo están entre los posibles factores que controlan la reabsorción de nutrientes (Del Arco et al., 1991; Rentería et al., 2005). Algunos autores postulan que la eficiencia en la reabsorción está influenciada por el incremento de la radiación debido a que el proceso de retranslocación requiere una cierta cantidad de energía que tiene que ser proporcionada por la fotosíntesis (Boerner, 1986). Adicionalmente, la radiación afecta al contenido de nitrógeno por unidad de área, el cual puede estar correlacionado con la eficiencia en la reabsorción (Nordell y Karlsson, 1995). Sin embargo, Yasumura et al. (2005) no detectaron relación alguna entre la radiación y la eficiencia de la reabsorción en tres especies caducifolias. Si este razonamiento es válido, la reabsorción puede o no mantener la producción de nuevas hojas u otra clase de estructuras y disminuir la demanda de minerales del suelo en especies caducifolias (Palma et al., 2000).
CONCLUSIONES
Los resultados emanados de la presente investigación demuestran que la producción de los diversos constituyentes de la hojarasca, cumplen con una función en un escenario fluctuante y complejo que cada vez requiere más atención para entender el funcionamiento de los ecosistemas y ciclaje de nutrientes, ya que por medio de la hojarasca se regenera el suelo, se evita la erosión, se mejoran las propiedades físicas y químicas del suelo, y se mantiene la fertilidad del mismo, además que ayuda a sostener la vida de la fauna heterótrofa.
Los resultados muestran que existe variación espacial y temporal en la producción de cada componente de la hojarasca, siendo el sitio de Linares el de mayor producción y Los Ramones el de menor. Las máximas producciones foliares se presentaron en la época de invierno (enero-abril). La producción total de ramas y de hojas, fue significativamente diferente en los tres sitios de estudio, mientras que en estructuras reproductivas y otros no se encontraron diferencias significativas. El sitio de Linares fue el más productivo y con mayor aporte potencial de componentes orgánicos depositados al suelo que los sitios de Los Ramones y China.
Asimismo, manteniendo la misma tendencia entre los sitios, se observaron diferencias en la productividad de nutrientes en cada sitio, mas no así en la calidad de las hojas, medida a través del aporte de minerales donde se encontraron porcentajes similares en Mg, P y N. Por tanto, lo que permite la diferencia en la aportación de minerales es la productividad de cada sitio de estudio; es decir, a mayor productividad mayor aportación de nutrientes. A pesar de pertenecer al mismo tipo de comunidad vegetal, los sitios de estudio difieren en el retorno potencial de nutrimentos y esta diferencia se relaciona a las condiciones edáficas, a la estructura florística, a la calidad de hojarasca, a los cambios estacionales, a las variaciones de temperatura y precipitación, los cuales conjuntamente influyeron en la eficiencia del uso de nutrientes.
En este estudio, tanto las características ecológicas, como las variables climáticas que se presentaron en cada sitio fueron determinantes en las producciones de hojarasca y minerales. En Linares fue donde se encontró la mayor cobertura, así como dominancia y frecuencia, en comparación con Los Ramones y China.
En cuanto al uso eficiente el P fue relativamente mayor que Ca, K, Mg y N. En este trabajo no se estudio la eficiencia de la retranslocación del P en la planta. Según estudios la retranlocación en especies caducifolias es un poco mayor que en especies perennifolias, por lo que se supone que en estos sitios puede darse la reabsorción de P para la producción de nuevas hojas u otras clases de estructuras, ya que la mayorías de las especies presentes en los sitios de estudios son caducifolias.
Dado que los estudios realizados en esta línea de investigación son mínimos, los resultados generados en el presente estudio indican la importancia de continuar investigando con mayor profundidad la relación suelo-planta o retorno de nutrimentos en comunidades vegetales de regiones semiáridas, sobre todo, si consideramos que este tipo de comunidades representan un 20% del territorio nacional; lo que implica más el entendimiento de la dinámica de la deposición de hojarasca y nutrimentos que contribuyan al mejoramiento y conservación de las propiedades físicas y químicas del suelo y a mantener los procesos ecológicos en este tipo de comunidades.
AGRADECIMIENTOS
Al Consejo Nacional de Ciencia y Tecnología (CONACyT) por proporcionar la beca de doctorado al primer autor. Se agradece a Tilo Gustavo Domínguez, Manuel Hernández Charles y Elsa Dolores González por su participación en los muestreos en campo y actividades de laboratorio. Además, se agradece también a los propietarios de los predios Rancho Zaragoza y Rancho El Abuelo por brindar todas las facilidades para llevar a cabo esta investigación. Adicionalmente, se agradece a tres revisores anónimos por enriquecer y hacer observaciones críticas al presente estudio. Al Proyecto PAICYT CT289-10 financiado por la Universidad Autónoma de Nuevo León.
LITERATURA CITADA
Aerts, R., 1996. "Nutrient resorption from senescing leaves of perennials: are there general patterns?" Journal of Ecology, 84: 597-608. [ Links ]
Andivia, E., Fernández, M., Vázquez, P.J., González, P.A. y Tapias, R., 2009. "Nutrient return from leaves and litterfall in a mediterranean cork oak (Quercus suber L.) forest in southwestern Spain". European Journal of Forest Research, 129: 5-12. [ Links ]
AOAC, 1997. Official methods of analysis, 16th ed., Association of Official Analytical Chemists, Washington, DC. [ Links ]
Boerner, R.E.J., 1986. "Seasonal nutrient dynamics, nutrient resorption and mycorrhizal infection intensity of two perennial forest herbs". American Journal of Botany, 73: 1249-1257. [ Links ]
Cantú, S.I., y González, R.H., 2001. "Interception loss, throughfall and stem flow chemistry in pine and oak forests in northeastern Mexico". Tree Physiology, 21: 1009-1013. [ Links ]
Caritat, A., García, B.E., Lapeña, R. y Vilar, L., 2006. "Litter production in a Quercus suber forest of Montseny (NE Spain) and its relationship to meteorological conditions". Annals Forest .Sciences, 63: 791-800. [ Links ]
Cherney, D.J.R., 2000. "Characterization of forages by chemical analysis". In: Givens DI, Owen E, Axford RFE, Omed HM (eds), Forage evaluation in ruminant nutrition. CAB International, Wallingford, pp. 281-300. [ Links ]
Del Arco, J.M., Escudero, A. y Garrido, M.V., 1991. "Effects of site characteristics on nitrogen retranslocation from senescent leaves". Ecology, 72: 701-708. [ Links ]
Del Valle-Arango, J.I., 2003. "Cantidad, calidad y nutrientes reciclados por la hojarasca fina de bosques pantanosos del Pacífico Sur Colombiano". Interciencia, 28: 443-449. [ Links ]
Domínguez, G.T.G., 2009. "Deposición de hojarasca y retorno potencial de nutrimentos en diferentes comunidades de vegetación". Tesis de maestría en ciencias forestales. Facultad de Ciencias Forestales, Universidad Autónoma de Nuevo León. Linares, Nuevo León, México. 132 pp. [ Links ]
Epstein, H.E., Burke, I.C. y Lauenroth, W.K., 2002. "Regional patterns of decomposition and primary production rates in the U.S. Great Plains". Ecology, 83: 320-327. [ Links ]
Gairola, S., Rawal, R. y Dhar, U., 2009. "Patterns of litterfall and return of nutrients across anthropogenic disturbance gradients in three subalpine forests of west Himalaya, India". Journal of Forest Research, 14: 73-80. [ Links ]
González, R.H., Cantú, S.I., Gómez, M.M.V y Ramírez, L.R.G., 2004. "Plant water relations of thornscrub shrub species, northeastern Mexico". Journal of Arid Environments, 58: 483-503. [ Links ]
González, R.H., Domínguez, G.T.G., Cantú, S.I., Gómez, M.M.V., Ramírez, L.R.G., Pando, M.M. y Fernández, C.J., 2011. "Litterfall deposition and leaf litter nutrient return in different locations at Northeastern México". Plant Ecology, 212: 1747-1757. [ Links ]
González, R.H., Ramírez, L.R.G, Cantú, S.I., Gómez, M.M.V. y Uvalle, S.J.I., 2010. "Composición y estructura de la vegetación en tres municipios del estado de Nuevo León". Polibotánica, 29: 91-106. [ Links ]
Imbert, J.B., Blanco, J.A. y Castillo, F.J., 2004. "Gestión forestal y ciclos de nutrientes en el marco del cambio global". F. Valladares (ed.). Ecología del bosque mediterráneo en un mundo cambiante. pp. 479-506. Ministerio de Medio Ambiente. Madrid, España. [ Links ]
Isaac, S.R. y Nair, M.A., 2006. "Litter dynamics of six multipurpose trees in a homegarden in Southern Kerala, India". Journal of Agroforestry System, 67: 203-213. [ Links ]
Kimmins, J.P., 1997. Forest ecology. A foundation for sustainable management. Prentice-Hall, Nueva Jersey. USA. [ Links ]
Lawrence, D., 2005. "Regional-scale variation in litter production and seasonality in tropical dry forests of Southern Mexico". Biotropica, 37: 561-570. [ Links ]
León, D.J., González, M.I. y Gallardo, J.F., 2011. "Ciclos biogeoquímicos en bosques naturales y plantaciones de coníferas en ecosistemas de alta montaña de Colombia". Journal of Tropical Biology, 59: 1883-1894. [ Links ]
Lousier, J.D. y Parkinson, D., 1976. "Litter decomposition in a cool temperate deciduous forest". Canadian Journal of Botany, 54: 419-436. [ Links ]
Marroquín, C.J.J., 2011. "Deposición de hojarasca y retorno de nutrientes en el matorral desértico micrófilo, Noreste de México". Tesis de de licenciatura. Facultad de Ciencias Forestales, Universidad Autónoma de Nuevo León. Linares, Nuevo León, México. 77 pp. [ Links ]
Martínez-Alonso, C., Valladares, F., Camarero, J.J., López Arias, M., Serrano, M. y Rodríguez, J.A., 2007. "The uncoupling of secondary growth, cone and litter production by intradecadal climatic variability in a mediterranean scots pine forest". Forest Ecology and Management, 253: 19-29. [ Links ]
Moro, M.J., 1992. "Desfronde, descomposición y fijación de nitrógeno en una microcuenca con repoblación joven de coníferas y matorral de Cistus laurifolius y Adenocarpus decorticans en la Sierra de los Filabres (Almería)". Tesis doctoral. Facultad de Ciencias. Universidad de Alicante, 463 pp. [ Links ]
Nambiar, E.K.S. y Fife, D.N., 1987. "Growth and nutrient retranslocation in needles of radiata pine in relation to nitrogen supply". Annals of Botany, 60: 147-156. [ Links ]
Nordell, K.O. y Karlsson, P.S., 1995. "Resorption of nitrogen and dry matter prior to leaf abscission: variation among individuals, sites and years in the mountain birch". Functional Ecology, 09: 326-333. [ Links ]
Northup, B.K., Zitzer, S.F., Archer, S.R. y Boutton, T.W., 1996. "A technique to allocate biomass of woody plants. La Copita Research Area: 1996 Consolidated Progress Report". Texas Agricultural Experiment Station, Corpus Christi, Texas A&M University System, College Station, Texas, USA, pp. 47-50. [ Links ]
Oelbermann, M. y Gordon, A.M., 2000. "Quantity and quality of autumnal litterfall into a rehabilitated agricultural stream". Journal of Environmental Quality, 29: 603-610. [ Links ]
Ott, L., 1993. An introduction to statistical methods and data analysis. 775 p. 2nd ed., Duxbury Press. Boston, Massachusetts. [ Links ]
Palma, R.M., Defrieri, R.L., Tortarolo, M.F., Prause, J. y Gallardo, J.F., 2000. "Seasonal changes of bioelements in the litter and their potential return to green leaves in four species of the Argentine subtropical forest". Annals of Botany, 85: 181-186. [ Links ]
Piatek, K.B., Alen, H.L., 2000. "Site preparation effects on foliar N and P use, retranslocation, and transfer to litter in 15-years old Pinus taeda". Forest Ecology and Management, 129: 143-152. [ Links ]
Prause, G.J., Arce, C. y Angeloni, P.N., 2003. "Variación mensual en el aporte de hojas de cuatro especies forestales nativas del Parque Chaqueño Húmedo (Argentina)". Revista de Ciencias Forestales-Quebracho, 10: 39-45. [ Links ]
Ramírez-Lozano, R.G., González, R.H., Gómez, M.M.V., Cantú, S.I. y Uvalle, S.J.I. 2010. "Spatio-temporal variations of macro and trace mineral contents in six native plants consumed by ruminants at northeastern Mexico". Tropical and Subtropical Agroecosystems, 12: 267-281. [ Links ]
Reid, N., Marroquín, J. y Beyer, M.P., 1990. "Utilization of shrubs and trees for browse, fuelwood and timber in the tamaulipan thornscrub, northeastern Mexico". Forest Ecology and Management, 36: 61-79. [ Links ]
Rentería, L.Y., Jaramillo, V.J., Martínez-Yrízar, A. y Pérez-Jiménez, A. 2005. "Nitrogen and phosphorus resorption in trees of a Mexican tropical dry forest". Trees, 19: 431-441. [ Links ]
Rocha-Loredo, A.G y Ramírez, M.N., 2009. "Producción y descomposición de hojarasca en diferentes condiciones sucesionales del bosque de pino-encino en Chiapas, México". Boletín de la Sociedad Botánica de México, 84: 1-12. [ Links ]
Rodríguez, H.E.J., 2010. "Producción de hojarasca y retorno potencial de nutrimentos, vía hojas en diferentes tipos de vegetación, noreste de México". Tesis de de licenciatura. Facultad de Ciencias Forestales, Universidad Autónoma de Nuevo León. Linares, Nuevo León, México. 60 pp. [ Links ]
Rodríguez, R.G.A., 1994. "Análisis de la fitodiversidad (sinusias: arbóreas y arbustivas) de dos comunidades del matorral espinoso tamaulipeco de Linares Nuevo León, México". Tesis profesional. F.C.B. Universidad Autónoma de Nuevo León. San Nicolás de los Garza, Nuevo León. [ Links ]
Sariñana, F.R., 1999. "Análisis de la fitodiversidad del estrato fisonómicamente dominante en 15 tipos de vegetación en Nuevo León, México". Tesis de maestría en ciencias forestales. Facultad de Ciencias Forestales, Universidad Autónoma de Nuevo León. Linares, Nuevo León, México. [ Links ]
Safou-Matondo, R., Deleporte, P., Laclau, J.P. y Bouillet, J.P., 2005. "Hybrid and clonal variability of nutrient content and nutrient use efficiency in Eucalyptus stands in Congo". Forest Ecology and Management, 210: 193-204. [ Links ]
Sánchez, F.C., Lama, D. y Suatunce, C.P., 2008. "Hojas caídas y aporte de nutrientes de diez especies forestales tropicales". Ciencia y Tecnología, 1: 73-78. [ Links ]
Santa Regina, I., 2000. "Biomass estimation and nutrient pools in four Quercus pyrenaica in Sierra de Gata Mountains, Salamanca, Spain". Forest Ecology and Management, 132: 127-141. [ Links ]
Santa Regina, I. y Tarazona, T., 2001a. "Nutrient pools to the soil through organic matter and throughfall under a Scots pine plantation in the Sierra de la Demanda, Spain". European Journal of Soil Biology, 37: 125-133. [ Links ]
Santa Regina, I. y Tarazona, T., 2001b. Organic matter and nitrogen dynamics in a mature forest of common beech in the Sierra de la Demanda, Spain. Annals of Forest Sciences, 58: 301-314. [ Links ]
Santa-Regina, J., Salazar, S., Leonardi, S. y Rapp, M., 2005. "Nutrient pools to the soil through organic matter in several Castanea sativa Mill. coppices of Mountainous mediterranean climate areas". Acta Horticulturae, 693: 341-348. [ Links ]
Steel, R.G.D. y Torrie, J.H., 1980. Principles and procedures of statistics. A biometrical approach, 2nd ed. New York, NY: McGraw-Hill Book Company. 633 pp. [ Links ]
Sundarapandian, S.M. y Swamy, P.S., 1999. "Litter production and leaf-litter decomposition of selected tree species in tropical forests at Kodayar in the Western Ghats, India". Forest Ecology and Management, 123: 231-244. [ Links ]
Swamy, S.L., Kushwaha, S.K. y Puri, S., 2004. "Tree growth, biomass, allometry and nutrient distribution in Gmelina arborea stands grown in red lateritic soils of central India". Biomass and Bioenergy, 26: 305-317. [ Links ]
Veneklaas, E.J., 1991. "Litterfall and nutrient fluxes in two montane tropical rain forests, Colombia". Journal of Tropical Ecology, 7: 319-336. [ Links ]
Villalón, M.H, Carrillo, P. A. y Soto, R.J.M., 1991. "Comparación de dos longitudes de líneas de muestreos para estimar coberturas en el matorral tamaulipeco mediante el método de puntos modificados". Facultad de Ciencias Forestales, Reporte Científico, 23: 1-19. [ Links ]
Vitousek, P.M., 1982. "Nutrient cycling and nutrient use efficiency". American Naturalist, 119: 553-572. [ Links ]
Wackerly, D.D., Mendenhall, W. y Scheaffer, R.L., 2002. Estadística Matemática con Aplicaciones. 6a. ed., Editorial Thomson. International. México. 872 pp. [ Links ]
Wood, T.E., Lawrence, D. y Clark, D.A., 2005. "Variation in leaf litter nutrients of Costa Rica rain forest is related to precipitation". Biogeochemistry, 73: 417-437. [ Links ]
Wright, I.J. y Westoby, M., 2003. "Nutrient concentration, resorption and lifespan: leaf traits of Australian sclerophyll species". Functional Ecology, 17: 10-19. [ Links ]
Yasumura, Y., Onoda, Y., Hikosaka, K. y Hirose, T., 2005. "Nitrogen resorption from leaves under different growth irradiance in three deciduous woody species". Plant Ecology, 178: 29-37. [ Links ]
Yang, Y.S., Guo, J.F., Chen, G.S., Xie, J.S., Cai, L.P. y Lin, P., 2004. "Litterfall, nutrient return, and leaf-litter decompositions in four plantations compared with a natural forest in subtropical China". Annals of Forest Science, 61: 465-476. [ Links ]
Yankelevich, S.N., Fragoso, C., Newton, A.C., Russell, G. y Heal, O.W., 2006. "Spatial patchiness of litter, nutrients and macroinvertebrates during secondary succession in a tropical montane cloud forest in Mexico". Plant and Soil, 286: 123-139. [ Links ]

