










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPolibotánica
versión impresa ISSN 1405-2768
Polibotánica no.33 México ene. 2012
Morfología y anatomía de radículas múltiples en plántulas de maíz derivadas de cariopsis con poliembrionía
Multiple morphology and anatomy radicles in derived maize seedlings from caryopses with polyembryony
José Espinoza-Velázquez1, Jesús Valdés-Reyna2 y José Manuel Alcalá-Rodríguez2
1 Instituto Mexicano del Maíz "Dr. Mario E. Castro Gil" de la Universidad Autónoma Agraria Antonio Narro (UAAAN). Buenavista, Saltillo, Coahuila, México. CP 25315. Correo electrónico: jespvel@uaaan.mx.
2 Universidad Autónoma Agraria Antonio Narro.
Recibido: 12 enero 2011.
Aceptado: 17 octubre 2011.
Resumen
Se presentan resultados relativos a la morfología y anatomía de radículas de maíz, así como de la frecuencia de plántulas y radículas múltiples por semilla germinada. Se utilizaron tres poblaciones y las seis cruzas posibles entre ellas. De las poblaciones, una corresponde a maíz común (no-poliembriónico) y dos de alta frecuencia poliembriónica (PE). Se indujeron a germinación 24 semillas de cada uno de los nueve genotipos. La morfología y anatomía de radículas se estudió en plántulas de tres días de edad, pero la identificación fenotípica y cuantificación de poliembrionía y radículas múltiples se llevó a cabo hasta los 10 días en plántulas contemporáneas a las primeras. Se evaluaron seis variables anatómicas de la radícula, valoradas en cortes histológicos longitudinales y transversales; también se determinaron las proporciones de radículas múltiples y poliembrionía. Los resultados indicaron que los fenómenos PE y radículas múltiples se presentaron solamente en las progenies de las dos poblaciones poliembriónicas y los híbridos entre ellas; las plántulas que exhiben PE pueden o no presentar simultáneamente el fenómeno de radículas múltiples; éstas fueron observadas en número y conformación variables i.e. en casos de dos y hasta cuatro raíces, separadas o con cierto grado de fusión a nivel histológico, incluyendo el cilindro vascular. El promedio de frecuencia de PE y de radículas múltiples fue de 62 y 14% respectivamente. Las características morfológicas y anatómicas de radícula con mayores atributos se observaron en algunas combinaciones híbridas entre genotipos PE x No-PE, lo cual pudiera significar una manifestación de heterosis, aprovechable agronómicamente en la utilización de ellos con otros grupos germoplásmicos.
Palabras clave: Zea mays L., poliembrionía, radículas múltiples por semilla, anatomía de radícula.
Abstract
Results related to maize seedlings anatomy and morphology as well as multiple seedlings and radicles per germinated seed are presented. Three populations and their six possible hybrids were used. Two populations are highly polyembryonic (PE) meanwhile the third represents a normal maize type (Non-Polyembryonic). 24 seeds from each genotype were induced to germination. The seedling's anatomy and morphology was studied in three days old seedlings, but the phenotypes and quantification of polyembryony and multiple radicles were done in contemporary seedlings of 10 days of age. Six anatomical variables, valued through longitudinal and transversal histological cuts, were evaluated; also, the multiple radicles and polyembryony proportions were determined. Results indicated that the PE and multiple radicles phenomena were only present in progenies of polyembryonic populations and their respective hybrids. There were cases where polyembryony and multiple radicles were simultaneous in the same seedling; these radicles varied in number and conformation, i.e. cases of two to four seedlings which can be fully separated or sharing some anatomical parts, including the vascular cylinder. The average frequencies for PE and multiple radicles were 62 and 14% respectively. The better values for anatomical and morphological characteristics were shown by some hybrids among the PE x No-PE genotypes; this could mean a positive heterosis which could be agronomically useful in handling them with other germplasm sources.
Key words: Zea mays L., polyembryony, multiple radicles per seed, morphology, radicle anatomy.
INTRODUCCIÓN
La raíz es un órgano fundamental cuya función principal es la de anclar a la planta y la de absorber agua y minerales. Por lo tanto, un sistema radical sano es determinante en el desarrollo de todos sus órganos aéreos (Gould y Shaw, 1983; Tuberosa y Salvi, 2007). Esta razón ha sido suficiente para estudiar las características de la raíz como apoyo en la mejora genética de cultivos (Mc Phee, 2005) y en la investigación para el análisis genético de la formación y desarrollo de la raíz (Hochholdinger, 2004; 2009).
La semilla del maíz común germina normalmente en una plántula, la cual presenta una radícula y una plúmula; sin embargo, se han detectado variantes naturales que alteran este patrón, provocando que sus semillas muestren de dos y hasta siete plúmulas de manera simultánea a la germinación (Espinoza et al., 1998); algunos de estos casos pueden mostrar también más de una radícula (Espinoza, V.J., datos sin publicar); estas condiciones son indicadores de la presencia de poliembrionía (PE) en este tipo de maíz, la cual ha demostrado ser de naturaleza heredable (Rebolloza etal., 2011).
La poliembrionía, presente en varios taxa de angiospermas y gimnospermas, tiene como consecuencia principal la producción de plúmulas múltiples y ha sido considerada una característica de valor agronómico (Webber, 1940; Ross y Wilson, 1969; Pesev et al., 1976; Bouquet, 1980; Nunez y Ocoy, 2008). En el proceso de germinación, el maíz común invierte normalmente todo su endosperma en el desarrollo de una sola plántula, mientras que los casos de cariopsis de naturaleza PE, de tamaño equivalente a la primera, debe invertir las reservas del endosperma en el desarrollo de dos o más plántulas.
Existe abundante información sobre desarrollo de la raíz en el maíz común (Hotchholdinger et al., 2004); el sistema radical se inicia con la aparición de la radícula y continúa con la expresión de raíces seminales laterales, en número variable; el crecimiento y desarrollo de éstas ocurre dentro de los cinco días a la siembra y cesa cuando la plántula alcanza el estado vegetativo de tres hojas (V3), de acuerdo a la terminología mundialmente aceptada de Ritchie et al. (1992), tiempo en que se extienden las raíces definitivas, denominadas "nodulares" o de corona, generadas a partir de los nudos basales en el tallo, las cuales se hacen aparentes desde la etapa V1 (Hochholdinger, 2004; 2009).
La anatomía de la radícula y raíces laterales en maíz muestran la organización típica de las monocotiledóneas (haces vasculares dispersos). El arreglo histológico comprende la epidermis, número variable de capas del córtex (de ocho a 15), endodermis, periciclo y un cilindro vascular con varios haces vasculares y floemas (Gould y Shaw, 1983). La organización longitudinal de este grupo de raíces es descrita en términos de zonas especializadas de desarrollo, y generalmente incluye la cofia, meristemo apical, zona de alargamiento distal, zona de alargamiento celular y zona de maduración, la cual está determinada por la formación de pelos radicales (Ishikawa y Evans, 1995; Hochholdinger, 2004; 2009).
Por otra parte, en la actualidad no se cuenta con información sobre desarrollo de radícula(s) en plántulas PE. En este contexto se presenta aquí un estudio anatómico de radícula(s) a los tres días de edad y se corrobora la presencia de dos o más radículas por semilla y su consecuencia a nivel anatómico, en dos poblaciones experimentales de alta frecuencia poliembriónica, denominadas C o UA-IMM-NAP (de porte normal) y D o UA-IMM- BAP (porte enano), generadas en el Instituto Mexicano del Maíz "Dr. Mario E. Castro Gil" de la Universidad Autónoma Agraria Antonio Narro (IMM-UAAAN), bajo la hipótesis de que la PE está asociada a procesos especiales en el desarrollo del sistema radical seminal.
MATERIAL Y MÉTODO
Los estudios de morfología y anatomía de radícula se llevaron a cabo en el laboratorio de Anatomía e Histología Vegetal, Departamento de Botánica, de la Universidad Autónoma Agraria Antonio Narro, sede en Saltillo, Coahuila, México (UAAAN). El material genético fueron nueve genotipos, producto de cruzas directas y recíprocas de tres poblaciones de maíz, denominadas C y D (desarrolladas en el IMM-UAAAN) y la E o Tuxpeño-HO, muestra otorgada por el Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT). Las poblaciones poliembriónicas C y D son materiales del tipo maíz mexicano, semidentado, de grano blanco, mientras que la población No-PE es un material de la raza tuxpeño, especializada en alto contenido de aceite en el grano. Los cruzamientos se llevaron a cabo siguiendo el método I de Griffing (1956), cuyos genotipos fueron, además de las tres poblaciones paternas (C, D y E), tres cruzas directas (CxD; CxE; y DxE) y tres recíprocas (DxC; ExC; y ExD). De sus progenies, se obtuvieron dos muestras aleatorias de 24 semillas, las cuales se hicieron germinar en rollos de papel germinador (tacos de germinación), dispuestas en un horno a 28 ± 2°C por tres días; radículas de esta edad fueron la base para los estudios de morfología.
Dos secciones de radícula de 2.5 cm de longitud fueron obtenidas después de 2 cm de la base de la semilla, en dirección al ápice radicular; el procedimiento se practicó en tres especímenes en cada uno de los nueve genotipos. Las radículas fueron colocadas en fijador FAA por 45 días; los tejidos ya fijados se sometieron a proceso de deshidratación y luego a inclusión en parafina, siguiendo la metodología de Johansen (1940) y Aguilar et al. (1996). El siguiente paso fue realizar los cortes transversales o longitudinales de radícula con el micrótomo rotatorio American Optical 820. La coloración se realizó con safranina fast-green. El grosor de los cortes fue de 16 μm. Para el análisis detallado de las imágenes se utilizó el software Axio Vision 4.7 de Carl Zeiss.
Una vez obtenidos los especímenes de radícula de tres días para los estudios anatómicos, se dejó que el resto de las plántulas siguieran su desarrollo en los tacos de germinación hasta alcanzar 10 días de edad, tiempo suficiente para apreciar y cuantificar el número de plúmulas y radículas por semilla germinada, en cada uno de los nueve genotipos. Las variables de estudio fueron: frecuencia de poliembrionía (FPE), longitud de radícula en cm (LR), número de radículas múltiples y su frecuencia (NRM; FRM). Las variables en cortes longitudinales de radícula(s) fueron: número de primordios de radícula (NPR), longitud de células corticales (LCC), capas del córtex (NCC), diámetro de radícula (DR), y número de haces vasculares (NHV). La longitud de radícula (LR) se tomó como referencia para analizar las tendencias de desarrollo de las características longitudinales, i.e. número de primordios radiculares (NPR) y la longitud de células del córtex (LCC), mientras que la variable diámetro de radícula (DR) fue la referente para analizar las tendencias de desarrollo de características transversales, i.e. número de capas corticales (NCC) y número de haces vasculares (NHV).
Análisis estadístico. Se utilizó un diseño completamente al azar, tres repeticiones; tres especímenes por genotipo. El modelo estadístico fue Yij = μ + ti + εij..; donde Yij representa la ij- ésima observación de cualquier variable, μ es la media general del experimento, ti representa el i-ésimo genotipo, y εij representa el ij- ésimo error experimental (Snedecor y Cochran, 1980).
RESULTADOS
Poliembrionía (PE)
Los valores de las variables obtenidos para plántulas a los 10 días de edad se presentan en el cuadro 1. Aquí, los nueve genotipos fueron agrupados como sigue: grupo PE (genotipos C y D), grupo PE x PE (cruzas directas y recíprocas entre C y D), grupo No-PE (tuxpeño, E), y grupo PE x No-PE (cruzas directas y recíprocas de E con C y D).

Se observó poliembrionía sólo en las poblaciones C y D, y las cruzas entre ellas (cuadro 1). La característica (FPE) fue cuantificada en el tiempo mínimo necesario para identificar fácil y claramente la condición de dos o más plántulas por semilla. En estos casos, la PE se presentó en frecuencias altas (superior a 60%); sin embargo, y de acuerdo a lo esperado, el carácter no se expresó en las F1 provenientes de cruzamientos entre genotipos PE x No-PE, ya que se ha demostrado que la PE es un carácter recesivo.
La variable frecuencia de radículas múltiples (FRM) del sistema radical seminal es también un fenómeno extraordinario en maíz, y se presentó sólo en genotipos PE y los cruzamientos entre ellos, en proporción promedio de 14%, valor inferior a la FPE; es decir, no todas las semillas que exhiben plúmulas múltiples pueden presentar también radículas múltiples. Una explicación al respecto es la condición de penetrancia incompleta del caracter PE (Rebolloza et al., 2011).
Morfología y anatomía de radícula(s)
Las variables relativas a la morfología y anatomía de radícula(s) de tres días de edad se consignan en el cuadro 2. La longitud promedio de las radículas fue de 8.7 cm; se encontró un incremento en esta variable en las combinaciones híbridas PE x E, las cuales presentaron un promedio de 9.2 cm, mientras que los genotipos PE tuvieron un promedio inferior (8.2 cm) a la media general y al genotipo E. De manera similar, la variable LCC presentó sus valores máximos en la hibridación de E con genotipos PE (promedió de 211 μm), condición superior a los valores que exhibieron los genotipos PE (promedio, 183 μm). La correlación de LR con NPR (r = 0.26) y LCC (r = 0.16) fueron positivas, bajas y no significativas; es decir, que una mayor longitud de la radícula no se traduce en mayor cantidad de primordios radiculares, ni en una mayor longitud de las células del córtex. Es notable que el NPR promedio en los genotipos PE sea el más bajo, mientras que los que fueron observados en E y sus cruzas como polinizador de madres PE, resultaron significativamente más altos.

Los primordios radiculares iniciaron su desarrollo a los tres días de edad (Fig. 1A) y no presentaron diferencias significativas entre genotipos. No obstante, la mayor densidad de los mismos fue registrada en híbridos entre E y los PE, tanto en cruzas directas como recíprocas; el promedio de éstos fue 11 primordios en 5 cm de radícula, valor superior a los que exhibieron los materiales PE como tales.
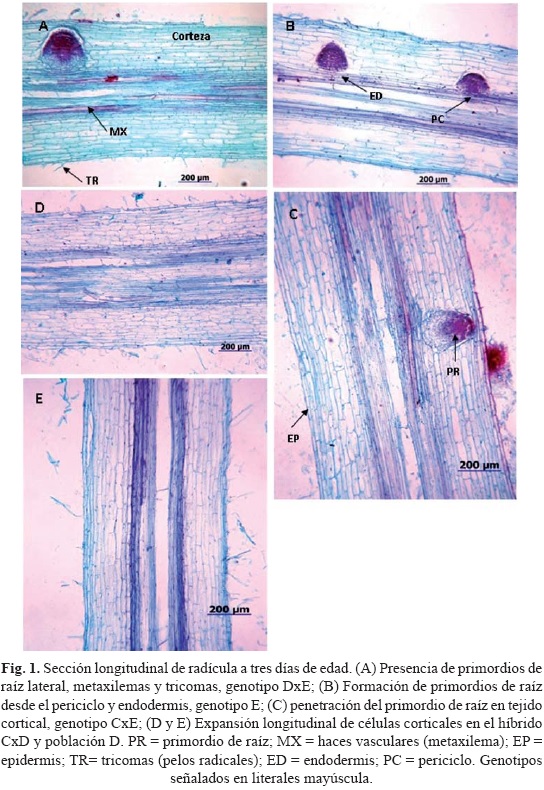
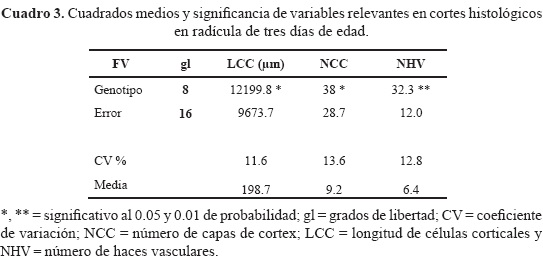
El DR (diámetro de radícula) no se correlacionó con NCC (r = 0.04) es decir, que el genotipo con mayor diámetro no presentó el mayor número de estratos del córtex. La asociación de DR con NHV fue positiva y significativa (r = 0.62, P ≤ 0.05) resaltando la asociación entre estas dos variables, aunque el DR, por sí solo, no presentó diferencias entre genotipos.
El análisis de varianza (cuadro 3) presentó diferencias estadísticas entre genotipos (P ≤ 0.05) sólo en las variables LCC, NCC y NHV; la comparación de medias (Tukey, α = 0.05) exhibió que el genotipo DxE fue el más alto para la variable LCC y que CxD presentó valores nominales altos para LR, no obstante que este híbrido ocupó el último lugar en LCC (Figs. 1D, 2a). En cuanto a NCC, las combinaciones más sobresalientes fueron DxE y CxE (Fig. 2b). Por otra parte, es razonable suponer que el NCC se correlacione positivamente con DR, aunque en este estudio, los genotipos de mayor número de estratos del córtex, no presentaron el mayor diámetro de radícula. Por último, para NHV la combinación CxD presentó la mayor cantidad de estas estructuras (Fig. 2c), incluso muy superior a genotipos de su misma condición PE.

Es relevante señalar que en esta etapa de desarrollo del sistema radical seminal, los genotipos de maíz PE mostraron diferentes tipos en la formación de radículas múltiples, los cuales se presentaron como independientes entre sí, o con cierto grado de fusión (Fig. 3A). Entre las variantes de radículas múltiples observadas fue posible documentar los casos siguientes: 1) dos radículas independientes, que se mostraron completamente normales; 2) dos radículas que comparten tejido cortical, pero cilindro vascular independiente; 3) dos radículas que compartían tanto el tejido cortical como el cilindro vascular. Es importante subrayar que estas tres variantes de radícula múltiple observadas corresponden a casos generados a partir de una semilla (Fig. 3D). En contra parte, los maíces comunes (E o sus cruzas con C y D) desarrollaron invariablemente sólo una radícula por semilla, anatómicamente normal (Figs. 3E, F).

DISCUSIÓN
La PE tiene implicaciones en el desarrollo inicial de la plántula; aquí, es destacable que sólo casos de genotipos PE presentaron plúmulas y radículas múltiples, lo cual puede considerarse como extraordinario, ya que el maíz común desarrolla invariablemente una de cada una de estas estructuras a la germinación. Además, dadas las frecuencias de las dos características, se puede plantear que la PE en los genotipos C y D presentan más de un tipo o versión del carácter, como las señaladas por Erdelska (1996) quien a partir de análisis histológicos establece que la poliembrionía en maíz se da en función del origen y la localización de los embriones en la semilla, y que los tipos probables son, 1) embriones gemelos provenientes de sacos embrionarios múltiples que pueden localizarse en lados opuestos o a distancia en la semilla, que no comparten tejido alguno y germinan por separado; 2) gemelos o tripletes que provienen de células individuales del aparato ovocélula, que pueden estar íntimamente unidos pero separados por capas epidérmicas, que comparten endosperma, pero poseen plúmulas y radículas por separado; y 3) poliembrionía por cleavage (multiplicación de embriones a partir de una célula fecundada), originada de manera espontánea o inducida, que comparten un suspensor común, parte del escutelo y capas superficiales de la radícula; por ello, germinan con sólo un complejo radicular pero con plúmulas por separado.
De acuerdo con lo anterior, la PE de los maíces C, D y sus cruzas, son mayormente del tipo (3), notable por ser casos de plúmulas múltiples y radícula única; la FRM en ellas se presentó en una proporción menor a casos PE, ya que el promedio de FRM fue de 14% mientras que la FPE fue de 63%, lo que pudiera indicar que cerca del 80% de éstos son tipo cleavage. Las proporciones de PE encontradas en este trabajo son prácticamente iguales a las señaladas por Espinoza et al. (1998).
Es oportuno señalar que el tamaño de las semillas PE es similar a los No-PE utilizadas en este trabajo (promedio de 280 mg ± 25); en los genotipos PE, la presencia de dos o más embriones en las semillas pudiera significar una disminución de tamaño del endosperma y, por lo tanto, una reducción en el volumen de reserva de nutrimentos; de ser así, se presentaría un menor volumen de nutrimentos a la germinación para abastecer los requerimientos de más de una plántula por semilla; probablemente, los casos de radículas múltiples pudieran ayudar a superar esta desventaja, aumentando la capacidad de absorción de agua y minerales en las primeras etapas de desarrollo de las plántulas.
Los resultados permitieron corroborar la presencia exclusiva de PE en las poblaciones C, D y cruzas entre ellas; también, que esta característica, en cruzamientos que generan a F1 con materiales normales (No-PE) se manifiesta como un carácter recesivo. Al respecto, la PE ha sido interpretada como un fenómeno gobernado por genes simples mendelianos, o como un carácter cuantitativo. Entre los primeros, está el reporte de Pilu (2000), y como carácter cuantitativo se tiene el registro de Espinoza et al. (1998); sin embargo, debido al comportamiento inconsistente en la fijación de la PE, Rebolloza et al. (2011) presentan una revisión de la genética del carácter, y proponen que la herencia de la PE corresponde a un fenómeno de interacción génica entre dos loci, del tipo epistasis doble recesiva, con penetrancia incompleta en la expresión del carácter.
Morfología y anatomía de radícula(s)
Los resultados permitieron corroborar que la radícula en maíz se hace visible como estructura morfológica al tercer día de la germinación; la radícula acompañada de raíces laterales seminales representan la mayor proporción de la plántula en las primeras dos semanas de desarrollo y es el sistema proveedor de agua y nutrientes en esta etapa; lo anterior, de acuerdo a lo reportado por Hochholdinger (2004; 2006; 2009).
La LR a esta edad alcanzó una media general de aproximadamente 9 cm; la mayor longitud fue alcanzada por las combinaciones híbridas de genotipos PE con E, mientras que los primeros tuvieron promedio menor a la media general y por debajo de E. Sin embargo, hay que hacer notar que en los casos de genotipos PE de radículas múltiples (dos a cuatro por semilla, presente en 14% de los casos) se consignó el promedio de longitud de las radículas y no la suma de ellas; esto, con el propósito de hacerlas comparables a las radículas simples. Por otra parte, la longitud de la radícula no fue un indicativo determinante para las variables longitudinales NPR y LCC. La falta de asociación de LR con NPR y LCC significó que la densidad de primordios observados en la radícula, ni la longitud de células del córtex se incrementaran por la mayor longitud de radícula. Se pudo corroborar que los primordios radicales son formados desde el periciclo y a veces de la endodermis, datos en concordancia con lo mencionado por Drubrovsky e Ivanov (1984), y que inician su desarrollo a los tres días, resultados similares a los reportados por Hoecker et al. (2006), quienes mencionan que las raíces laterales de radícula en maíz inician su desarrollo aproximadamente a los cuatro días y se hacen visibles al quinto día.
La mayor cantidad de primordios fue observada en híbridos resultantes entre genotipos PE con E, en cualquier sentido de cruza. Al comparar datos de radículas de genotipos sólo PE se debe tomar en cuenta que la semilla genera dos o más plúmulas y/o radículas al germinar; esto implica un gasto energético mayor, a partir del mismo monto o menos de las reservas nutrimentales que portan las semillas normales; éstas hacen gasto para una sola plántula y un solo sistema radical seminal; de esta manera, es explicable que en esta etapa; la semilla con embrión múltiple al germinar reduzca de algún modo la velocidad de sus procesos de desarrollo.
La radícula y los primordios de ésta, una vez emergidos, poseen la capacidad de formar raíces secundarias, terciarias y de más alto orden; éstas contribuyen a la densidad y arquitectura de la raíz, lo cual fortalece el anclaje temprano y la absorción de agua y nutrientes (Lynch, 1995; Hochholdinger y Feix, 1998; Hochholdinger, 2004; 2009). De esta comprensión del sistema radical seminal, se puede suponer que el aparente retraso en el desarrollo de plántula en genotipos PE pudiera afectar, de algún modo, el establecimiento de la plántula en las etapas muy juveniles.
El sistema radical seminal (radícula y las raíces laterales), es suficiente para formar una planta madura y fértil; esto fue demostrado por la acción del mutante monogénico recesivo rtcs que limita la aparición de las raíces laterales seminales y nodulares (Hetz et al., 1996). En este sentido, es posible y recomendable seleccionar genotipos PE con aptitud para desarrollar, a mayor velocidad, un mejor sistema radical seminal.
Desde el punto de vista transversal el DR influyó positivamente la aparición de un mayor número de haces vasculares (NHV) en algunos genotipos, como el caso del híbrido CxD, el cual mostró valores mayores en esta variable, influenciado mayormente por la condición de dos radículas unidas, que comparten tejidos en su estructura anatómica. Por otra parte, el hecho de que el DR no haya presentado diferencias entre genotipos pudiera significar la poca efectividad de la característica en el desarrollo de raíz en esta etapa, porque de tenerlo DR estaría correlacionada con el incremento en volumen de la radícula. Estudios previos han encontrado que es la longitud y no el diámetro de radícula lo que potencia el desarrollo de plántula, ya que una mayor elongación de la radícula logra incrementar de manera eficiente la superficie exploratoria y la absorción de las raíces (Hoecker et al., 2006).
Las células del córtex forman uno de los tejidos que componen en mayor parte la radícula en maíz (Hochholdinger, 2004; 2009); se sabe también que la LCC es el mayor factor de la elongación de la radícula (Hoecker et al., 2006). En este trabajo, la LCC fue mayor en la hibridación entre genotipos PE con E, aunque no queda plenamente claro si esta mayor longitud en los híbridos influye positivamente en cuanto a longitud de radícula. Esto se aprecia en la cruza CxD, que es del tipo PE, la cual se presenta entre los promedios más altos para LR, pero es el genotipo de menor valor en cuanto a LCC; esta condición sugiere que la LR pudiera estar asociada también al número de células contenidas en este órgano.
El número de capas del córtex (NCC) fue alta en las combinaciones híbridas PE x E. Esta respuesta parece tener una relación genética al combinar germoplasma contrastante donde la fuente materna es de naturaleza PE. Por otra parte, es razonable que NCC correlacione positivamente con el DR; sin embargo en este estudio los genotipos con mayor NCC no presentaron el mayor DR. De cualquier modo, el número de capas del córtex observados en los especímenes analizados están dentro de la amplitud conocida (8-15 estratos celulares) como se informa por Hochholdinger (2004; 2009).
Dentro del cilindro vascular de la radícula, los haces vasculares tienen la capacidad de transportar agua y nutrientes (Hochholdinger, 2004; 2009). Los resultados en este trabajo permiten señalar que la combinación CxD sobresale con la mayor cantidad de estas estructuras, incluso muy superior a los genotipos de su misma condición PE. Esta situación no puede ser atribuida a alguna clase de heterosis, ya que sus progenitores comparten un mismo origen genético y su mayor diferencia se refiere al porte de planta (enana vs. normal); por lo tanto, la expresión superior puede estar influida por su condición PE, ya que en los tres especímenes analizados presentaron radículas dobles fusionadas en tejido vascular. En contraparte, las muestras de sus similares (C, D y DxC) sólo mostraron un caso de esta particularidad en sus radículas.
Destacable y novedoso en esta etapa de desarrollo de los genotipos PE de maíz fue la expresión de diferentes versiones de radículas múltiples, separadas o con cierto grado de fusión, exclusivas de genotipos PE. En este contexto, Erdelska (1996) menciona dos de tres tipos de PE en maíz que describen el fenómeno; una de ellas corresponden a ovocélulas que están cercanamente adheridas pero separadas estrictamente por el tejido epidérmico con radículas independientes; la otra manifiesta un suspensor común, en el cual se comparten capas superficiales de la radícula; sin embargo, este autor no hace mención de la fusión de tejidos vasculares, como el que se documenta en este trabajo, que agrega un dato al concepto de tejidos compartidos en la producción de radículas múltiples en maíz.
CONCLUSIONES
La poliembrionía en maíces IMM-UAAAN, poblaciones C, D y cruzas entre ellas, es un carácter que influye de manera notable el desarrollo del sistema radical seminal obligando, en proporción importante, a la emisión de dos y hasta cuatro radículas, las cuales pueden emerger de manera separada o unidas en una especie de cableado, que comparten varios tejidos, incluso parte del cilindro vascular. La hibridación con material genéticamente distante (No-PE) genera progenies que no presentan PE ni casos de radículas múltiples, ratificando que el carácter que los influye (la PE) es de naturaleza recesiva; en general, el desarrollo del sistema radical seminal en la etapa de tres días de edad, es más rápido en los genotipos no-poliembriónicos, probablemente por que en las semillas PE reparten sus recursos nutrimentales en la emisión de dos o más plúmulas.
AGRADECIMIENTOS
Al personal del Laboratorio de Histología Vegetal del Departamento de Botánica de la Universidad Autónoma Agraria Antonio Narro por apoyar los trabajos de esta investigación.
LITERATURA CITADA
Aguilar M.M. y Coutino, 1996. Manual General de Técnicas Histológicas y Citoquímicas. Coordinación de Servicios Editoriales, Facultad de Ciencias. UNAM. México, DF, 130 p. [ Links ]
Bouquet, A., 1980. "Effect of some genetic and environmental factor son spontaneous polyembryony in grapes (Vitis vinifera L.)". Vitus, 19(2): 134-150. [ Links ]
Dubrovsky, J. y Ivanov, V., 1984. "Certain mechanisms of lateral root initiation in germinating maize roots". Phys. Biochem. Cultivated Plants, Kiev, 16: 279-284. [ Links ]
Erdelska, O., 1996. "Polyembryony in maize-histological analysis". Acta Soc. Bot. Poloniae vol. 65(1-2) CTOP. 001123-00125. [ Links ]
Espinoza, J., Vega, C., Navarro, E. y Burciaga, G., 1998. "Poliembrionía en maíces de porte normal y enano". Agron. Mesoam., 9(2):83-88. [ Links ]
Gould, F.W. y R.B. Shaw. 1983. Grass Systematics. 2nd. ed. College Station, Texas A & M Univ. Press., 397 pp. [ Links ]
Griffing, B., 1956. "Concept of general and specific combining ability in relation to diallel crossing systems". Aus. J. Bio. Sei., 9: 463-493. [ Links ]
Hetz, W., Hochholdinger, F., Schwall. M. y Feix, G., 1996. "Isolation and characterization of rtcs a mutant deficient in the formation of nodal roots". Plant J., 10: 845-857. [ Links ]
Hochholdinger, F. y Feix, G., 1998. "Early post-embryonic root formation is specifically affected in the maize mutant lrtl". Plant J., 16: 247-255. [ Links ]
Hochholdinger, F., 2004. Genetics of root formation in maize (Zea mays L.) "Reveals Root-Type specific developmental programs". Ann. Bot., 93: 359-368. [ Links ]
Hochholdinger, F., 2006. "Research interest: Genetic analysis of root formation in maize (Zea mays)". Available in http://www.uni-tuebingen.de/genetics/hochh/hochh.htm. Accessed 25. January 2007. [ Links ]
Hochholdinger, F., 2009. "The maize root system: Morphology, Anatomy, and genetics". J.L. Bennetzen and S.C. Hake (eds.), Handbook of maize: Its Biology. Springer Science, 145-160. [ Links ]
Hoecker, N., Keller, N., B., Piepho H. P., Hochholdinger, F., 2006. "Manifestation of heterosis during early maize (Zea mays L.) root development". Theor. Appl. Genet., 112: 421-429. [ Links ]
Ishikawa, H. y Evans, M.L., 1995. "Specialized zones of development in roots". Pl. Physiol., 109: 725-727. [ Links ]
Johansen, D.A., 1940. Plant Microtechniques. McGraw-Hill. New York, USA. [ Links ]
Lynch, J., 1995. "Root architecture and plant productivity". Pl. Physiol., 109: 7-13. [ Links ]
McPhee, K., 2005. "Variation for seedling root architecture in the core collection of pea germplasm". Crop Sci., 45: 1758-1763. [ Links ]
Nunez, T.C. y E.T. Ocoy, 2008. "Polyembryony in makapuno (Cocos nucífera L.) hybrids". Phillipine Jour. Crop Sci., 122-123. [ Links ]
Pesev, N., R. Petrovic, Lj. Zecevic y M. Milosevic, 1976. "Study of possibility in raising maize inbred lines with two embryos". Theor. Appl. Genet., 47: 74-51. [ Links ]
Pilu, R., 2000. "The twin trait maize". Maize Gen. Coop. News, 74: 51. [ Links ]
Rebolloza, H.H., J. Espinoza V., D. Sámano G. y V. M. Zamora V, 2011. "Herencia de la poliembrionía en dos poblaciones experimentales de maíz". Rev. Fitotec. Mex., 34(1): 27-33. [ Links ]
Ritchie, S.W. Hanaway, J.J. y Benson, G.O., 1992. How a corn plant develops. Special report No. 48. Iowa State University. Ames, IA, USA. [ Links ]
Ross, W.M. y J.A. Wilson, 1969. "Polyembryony in sorghum". Crop Sci., 9: 842-843. [ Links ]
Snedecor, G.W. y W.G. Cochran. 1980. Statistical Methods. The Iowa State University Press. Ames, Ia. USA. [ Links ]
Tuberosa, R. y Salvi, S., 2007. "Form the QTLs genes controlling root traits in maize". J.H.J. Spiertz, P.C. Struik and H.H. van Laar (eds.), Scale and Complexity in Plant Systems Research: Gene-Plant-Crop Relations, 15-24. [ Links ]
Webber, J.M., 1940. "Polyembryony". Bot. Rev., VI(11): 575-598. [ Links ]

