










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPolibotánica
versión impresa ISSN 1405-2768
Polibotánica no.32 México ago. 2011
Micromorfología de ceras epicuticulares en hojas maduras de Sabal yapa Wright ex Becc. (Arecaceae)
Micromorphology of epicuticular waxes in mature leaves of Sabal yapa Wright ex Becc. (Arecaceae)
Martha Pérez-García1 y José David Sepúlveda-Sánchez2
1 Departamento de Biología, División de Ciencias Biológicas y de la Salud, Universidad Autónoma Metropolitana Iztapalapa (UAM-I). Av. San Rafael Atlixco núm. 186. Col. Vicentina, Iztapalapa, DF, CP 09340.
2 Laboratorio de Microscopía Electrónica del edificio de Ciencia y Tecnología Ambiental (CYTAM), Universidad Autónoma Metropolitana Iztapalapa (UAM-I). Av. San Rafael Atlixco núm. 186. Col. Vicentina, Iztapalapa, DF, CP 09340. Correo electrónico: mpg@xanum.uam.mx
Recibido: 22 enero 2010.
Aceptado: 31 marzo 2011.
Resumen
Se reporta la presencia de cera epicuticular sobre la cutícula de la epidermis de las superficies adaxial y abaxial de la lámina de la hoja de Sabal yapa Wright ex Becc., observada con el microscopio electrónico de barrido (MEB). Las muestras foliares fueron colectadas en una vegetación secundaria de la selva mediana subperennifolia en Quintana Roo y en dos huertos de Yucatán. Se describió la micromorfología de la cera (siguiendo la clasificación de Barthlott et al., 1998); se observaron dos tipos de cristales de ceras en ambas superficies epidérmicas. El primer tipo se caracteriza por cristaloides denominados plaquetas irregulares, planos, sobresaliendo perpendicularmente desde la superficie, con bordes sinuosos y por su orientación se considera que son del tipo de plaquetas paralelas descritas como "tipo Convallaria". El otro tipo de cera se describe como cubiertas continuas sobre los estomas y son denominadas capas lisas. Son necesarios más estudios en especies de palmas, y contar con una definición clara de las ceras de este grupo, así como de su significado e importancia.
Palabras clave: ceras epicuticulares, cristaloides, cutícula, MEB, micromorfología, Sabal yapa.
Abstract
This paper reports the presence of epicuticular wax on the epidermal cuticle of the adaxial and abaxial surfaces of the leaf blade of Sabal yapa Wright ex Becc. Leaf samples were collected in secondary growth of subdeciduos forest in the State of Quintana Roo and in two home gardens in the State of Yucatan. Wax micromorphology observed with scanning electron microscopy (SEM) showed two types of crystals (following the classification of Barthlott et al., 1998) on both epidermal surfaces. The first type is characterized by crystalloids called irregular platelets, which are flat with sinuate margins and protrude perpendicularly from the surface. In their orientation they are arranged like the parallel platelets described as the "Convallaria-Type". The other type consists of continuous coverings over the stomata that are called smooth layers. Further studies on palm waxes, as well as a refined classification, are necessary to compare and clearly define their micromophology and their role and importance.
Key words: crystalloids, cuticle, epicuticular waxes, micromorphology, Sabal yapa, SEM.
INTRODUCCIÓN
Todas las partes primarias de la planta (excepto las raíces) están cubiertas por una cutícula que es la interfase entre las plantas y su medio ambiente, y que a su vez está revestida por una capa continua de cera en su superficie. Esta capa protege a la planta de varios factores ambientales como la sequía y el daño por radiaciones UV y actúa como una defensa contra patógenos como insectos, bacterias y hongos (Taiz y Zieger, 1991; García et al., 1995; Rashotte et al., 1997).
Las ceras están embebidas en una matriz polímera (cera intracuticular) y también depositadas sobre la superficie (ceras epicuticulares). Una de sus funciones más importantes es servir como una barrera contra la pérdida descontrolada de agua (Riederer and Schreiber, 1995); las ceras vegetales son las más repelentes al agua de todos los lípidos (Mauseth, 2003; Raven et al., 2005).
En muchas plantas la cera es exudada sobre la superficie de la cutícula, ya sea en láminas lisas, costras o como otras formas que se pueden extender hacia arriba a partir de la superficie cuticular. Esta estructura micromorfológica muestra una enorme variedad de estructuras tridimensionales funcionales (Barthlott et al., 1998, 2003; Koch et al., 2004).
Estas superficies frecuentemente desarrollan una propiedad de autolimpieza llamada 'efecto lotus' (efecto loto) y se caracterizan por el incremento de la repelencia al agua, por la reducción de la adhesión de las partículas contaminantes a la superficie foliar y son de gran interés tecnológico (Barthlott y Neinhuis, 1997; Guo y Liu, 2007).
Debido a que las ceras son cristalinas, la forma variada de los cristales está determinada por su composición química, pero por su tamaño pequeño, la composición química de un cristal de cera individual es aún desconocida (Koch et al., 2004).
El arreglo irregular de los bastones, plaquetas u otras formas, puede transformar a las superficies foliares en pantallas solares efectivas. Pero también se ha observado que ciertos contaminantes como el ozono y la lluvia ácida interfieren con el desarrollo de la cutícula y de la cera, causando daño considerable a la epidermis y a los tejidos internos (Baker y Hunt, 1986; Farmer, 1993; Shepherd y Griffiths, 2006).
Con la introducción del microscopio electrónico de barrido (MEB), muchas superficies cuticulares pudieron ser estudiadas más detalladamente. Una revisión posterior de las clasificaciones de estas ceras fue hecha por Jeffree (1986) quien resumió la mayor parte de la literatura publicada y propuso 14 tipos morfológicos de ceras basado en los resultados de las investigaciones de cerca de 480 taxa. Más adelante, en un estudio hecho en cutículas de plantas con semillas se identificaron 23 tipos de ceras y en este mismo se refieren a las ceras como cristales y el término de cristales de cera es ampliamente utilizado a partir de este trabajo (Barthlott et al., 1998).
En las palmas, a pesar de ser plantas con muchos y variados usos sobre todo en zonas rurales, no se encontró casi ningún estudio sobre micromorfología de ceras epicuticulares, excepto para las palmas de importancia económica como en el caso de la cera de carnauba (Vandenburg y Wilder, 1970; Barthlott et al., 1998), aunque sí hay algunas especies de palmas (García et al., 1995; Rashotte et al., 1997; Escalante et al., 2002), pero no del género Sabal.
El objetivo de este trabajo fue el de detectar la presencia de cera y describir su micromorfología tanto en la cutícula adaxial como abaxial, de una especie de palma que crece en la península de Yucatán.
METODOLOGÍA
Se colectaron dos hojas de Sabal yapa en cada uno de los siguientes sitios: vegetación secundaria de selva mediana subperennifolia (Miranda y Hernández, 1963) del municipio de Felipe Carrillo Puerto, Quintana Roo y en dos huertos familiares de Maxcanú, municipio de Maxcanú, Yucatán. El municipio de Felipe Carrillo, se localiza en el centro del estado de Quintana Roo a los 19°34'43" latitud N y 88°02'43" longitud O y se ubica a los 30 m.s.n.m. El municipio de Maxcanú está situado entre los paralelos 20°33' latitud N y 89°53' longitud O y se ubica a los 21 m.s.n.m. (Figs. 1 y 2).


Sabal yapa Wright ex Becc., es una especie de palma que llega a medir hasta 25 m de altura, con tronco regularmente grueso y anillado. Crece tanto en vegetación natural como asociada a la actividad antropogénica. Sus hojas son filíferas, alternas y dispuestas helicoidalmente. Las láminas foliares están compuestas por numerosos segmentos, de 90 cm a 1.5 m de longitud y son más o menos rígidos, se fusionan en la base, formando una palma larga. Cada segmento tiene tres nervaduras principales, una central y dos laterales. La lámina posee una costa que es recurvada. La hástula es adaxial, triangular, prominente de 4 a 8 cm de longitud y de 4 a 6 cm de ancho en la parte media, convexa abaxialmente con la superficie adaxial cóncava y con numerosos canales longitudinales (Figs. 3 y 4).


Las muestras de los segmentos de la lámina se obtuvieron de la parte media de los segmentos foliares, en secciones de 1 cm por lado y fueron fijadas en glutaraldehido al 2% en buffer de fosfato; el material vegetal fue procesado de acuerdo con Bozzola y Rusell (1992) y que se presenta en la tabla 1; las imágenes fueron observadas con el Microscopio Electrónico de Barrido (MEB) marca JEOL, modelo JSM-5900 LV.
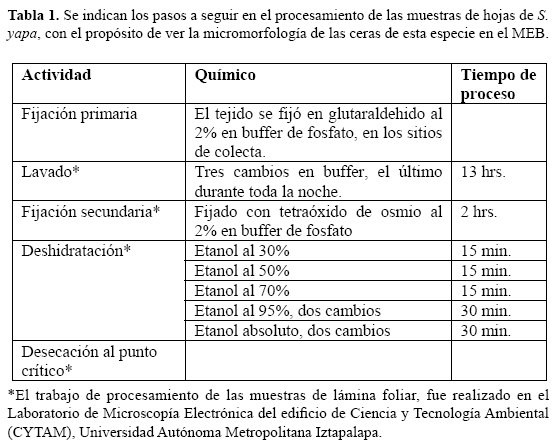
En 1998 Barthlott et al. publicaron un artículo en donde elaboraron una clasificación y establecieron una terminología para los cristales de cera; éstas fueron utilizadas para la identificación y descripción micromorfológica de las ceras de las superficies de las láminas foliares de S. yapa.
RESULTADOS
Fueron revisados los trozos de lámina de la hojas de S. yapa colectadas en los cuatro sitios ya mencionados en el microscopio electrónico de barrido y se observó que no había diferencia en cuanto a morfología de los cristales de cera, entre las hojas colectadas en cada sitio de colecta, ni entre las superficies abaxial y adaxial.
La micromorfología de estas hojas presentó dos tipos de cristales de ceras en ambas superficies, adaxial y abaxial, no se observó ninguna diferencia en forma y orientación de los cristales en ambas caras de la lámina foliar. El primer tipo se caracteriza por cristaloides cuya micromorfología y orientación es similar pero no igual a las denominadas plaquetas irregulares. Éstos son cristaloides planos, más o menos membranosos y con bordes sinuosos, no interconectados, dispuestos sobre las nervaduras de la lámina de la hoja, y por su orientación se considera que son del tipo de plaquetas paralelas (Figs. 5, 6, 8 y 9). Éstas fueron descritas como "tipo Convallaria" en la clasificación que fue utilizada. El otro tipo de cera, fue observada únicamente sobre los estomas de ambas superficies (lámina anfiestomática), que son intercostales y presentan otro tipo de cera que es una capa cerosa más o menos lisa, gruesa y no está esculpida (Figs. 5, 6, 7, 8, 9 y 10 ).






Muchas de las palmas crecen a mayor altura que el resto de la vegetación de la selva, o también lo hacen en lugares perturbados por actividades antrópicas, están muy expuestas a la luz solar y se esperaría que las ceras epicuticulares tuvieran morfología, fisiología y composición química que les brindara mayor protección contra la radiación solar y la temperatura excesiva; no se pudo saber si la disposición de las ceras en estas hojas tiene alguna función ya que es necesario hacer trabajo experimental, que no fue hecho, y en relación a la composición química de los cristaloides es desconocida para este género.
DISCUSIÓN
El tipo de cristaloides en Convallaria majalis L., con orientación paralela dio lugar a este tipo de plaquetas paralelas o "tipo Convallaria" y los estudios indican que son características de la familia de las lilifloras. Las plaquetas y las placas son cristaloides de amplia distribución entre las plantas y por eso son de poco significado sistemático, a menos que estén ordenadas en patrones de orientación característicos; las condiciones del medio ambiente influyen en la micromorfología de las ceras y sobre este aspecto aún hay escasa información (Engel y Barthlott, 1988).
No fue posible establecer si las ceras juegan un papel importante, relacionado con la protección contra la luz y temperatura excesivas en estas palmas, y debido a su estructura micromorfológica común a varias especies de plantas, se podría inferir, sin asegurar, que no tienen importancia taxonómica.
Los resultados muestran además que, en especial en las palmas, hace falta investigación sobre ceras epicuticulares, tanto morfológica como química y su papel ecofisiológico en los ambientes donde se desarrollan.
AGRADECIMIENTOS
Damos las gracias al Laboratorio de Microscopía Electrónica del Centro Nacional de Investigación y Capacitación Ambiental (CENICA). Instituto Nacional de Ecología, SEMARNAT, ubicado en el edificio de Ciencia y Tecnología Ambiental, Universidad Autónoma Metropolitana Iztapalapa, por el uso de su microscopio electrónico de barrido.
LITERATURA CITADA
Baker, E. A. y G. M. Hunt, 1986. "Erosion of waxes from leaf surfaces by simulated rain". New Phytol., 102: 161-173. [ Links ]
Barthlott, W. y C. Neinhuis, 1997. "Purity of the sacred lotus, or escape from contamination in biological surfaces". Planta, 202: 1-8. [ Links ]
Barthlott, W., C. Neinhuis, D. Cutler, F. Ditsch, I. Meusel, I. Theisen y H. Wilhelmi, 1998. "Classification and terminology of plant epicuticular waxes". Bot. J. of Linnean Society, 126: 237-260. [ Links ]
Barthlott, W., I. Theisen, T. Borsch, C. Neinhuis, 2003. "Epicuticular waxes and vascular plant systematics: integrating micromorphological and chemical dat". In: Stuessy T. F., Mayer V., Hõrandl E. (eds.) Deep morphology: toward a renaissance of morphology in plant systematics. Regnum Vegetabile vol. 141. Ruggell, Liechtenstein: Ganter Verlag, 189-206. [ Links ]
Bozzola, J.J. y L. Rusell, 1992. Electron Microscopy. Principles and Techniques for Biologists. John and Bartlett Publishers. Boston, USA. [ Links ]
Bussotti, F. y P. Grossoni, 1997. "European and Mediterranean oaks (Quercus L.; Fagaceae): SEM characterization of the micromorphology of the abaxial leaf surface". Botanical Journal of the Linnean Society, 124: 183-199. [ Links ]
Engel, T. y W. Barthlott, 1 988. "Micromophology of epicuticular waxes in Centrosperms". Plant Systematics andEvolution, 161: 71-85. [ Links ]
Escalante, E.F., M.R. Gamboa, J.G. Lecher, G.A. Arroyo, D. Zizumbo, C. Oropeza y L.M. Peña, 2002. "Major components from the epicuticular wax of Cocos nucifera. Rev. Soc. Quim. Mex., 46(3): 247-250. [ Links ]
Farmer, A.M., 1993. "The effects of dust on vegetation—a review". Environmental Pollution, 79: 63-75. [ Links ]
García, S., H. Heinzen, C. Hubbuch, R. Martínez, X. De Vries y P. Moyna, 1995. "Triterpene metil ethers from palmae epicuticular waxes". Phytochemistry, 39: 1381-1382. [ Links ]
Guo, Z. y W. Liu., 2007. "Biomimic from the superhydrophobic plant leaves in nature: binary structure and unitary structure". Plant Science, 172: 11031112. [ Links ]
Jefree, C.E., 1986. "The cuticle, epicuticular waxes and trichomas of plants, with reference to their structure, functions and evolution". In, B.E. Juniper y S.R. Southwood (eds.) Insects and the Plant Surface. Edward Arnold. London. pp. 23-63. [ Links ]
Koch, K, C. Neinhuis, H-J. Ensikat y W. Barthlott, 2004. "Self assembly of epicuticular waxes on living plant surfaces imaged by atomic force microscopy (AFM)". J. Exp. Bot., 55(397): 711-718. [ Links ]
Mauseth, J.D., 2003. Botany. An Introduction to Plant Biology. Jones and Bartlett Publishers, Inc. U.S.A. [ Links ]
Miranda, F. y E. Hernández, 1963. "Los Tipos de Vegetación de México y su Clasificación". Bol. Soc. Bot. Méx., 28: 29-179. [ Links ]
Rashotte, A., M. Jenks, T. Nguyen y K. A. Feldman, 1997. "Epicuticular wax variation in ecotypes of Arabidopsis thaliana". Phytochemistry, 45(2): 251-255. [ Links ]
Raven, P.H., R.F. Evert y S.E. Eichhorn, 2005. Biology of Plants. W.H. Freeman and Company Publishers. U.S.A. [ Links ]
Riederer, M. y L. Shreiber, 1995. "Waxes-the transport barriers of plant cuticles". In: Hamilton R. J., (ed.) Waxes: chemistry, molecular biology and functions. The Oily Press, pp. 131-156. [ Links ]
Shepherd, T. y D. W. Griffiths, 2006. "The effects of stress on plant cuticular waxes". New Phytologist, 171: 469499. [ Links ]
Taiz, L. y E. Zeiger, 1991 . "Surface protection and secondary defense compounds". In: Plant Physiology. Benjamin/Cummings Redwood City, California, USA. [ Links ]
Vandenburg, L.E. y E.A. Wilder, 1970. "The structural constituents of carnauba wax". Journal of the American Oil Chemistry Society, 47: 514-518. [ Links ]

