










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkPolibotánica
Print version ISSN 1405-2768
Polibotánica n.32 México Aug. 2011
Características funcionales de los principales bosques de Cuba oriental
Functional characteristics of the major forests of eastern Cuba
Orlando J. Reyes* y Euclides Fornaris-Gómez
Centro Oriental de Ecosistemas y Biodiversidad (BIOECO) CITMA, José A. Saco 601, esq. a Barnada, CP 90 100, Santiago de Cuba, Cuba. *Correo electrónico: joel@bioeco.ciges.inf.cu
Recibido: 6 agosto 2008.
Aceptado: 6 mayo 2011.
Resumen
Se estudió el funcionamiento de ocho de los principales ecosistemas de Cuba Oriental, los que abarcan el mayor rango de variación de las condiciones ecológicas (altitudinales, rocas, suelos, precipitación) y de la vegetación. Para el estudio de cada uno de los ecosistemas se utilizó el método de Herrera & Rodríguez (1988), con la modificación que se utilizó la fórmula (L+F)/H para conocer la velocidad general de descomposición del mantillo. Se corrobora que la esclerofilia y la esclerorrizia aumentan en función de la acentuación de las condiciones ecológicas estresantes. Los menores valores de la relación (L+F)/H (ecosistemas eueremazimóticos), se observan en ecosistemas muy húmedos y con suelos muy pobres, donde se observan las mayores tensiones ambientales.
Palabras clave: bosques tropicales, funcionamiento, esclerofilia, mantillo, descomposición.
Abstract
The functioning of eight main ecosystems of eastern Cuba was studied, which cover the widest range of variation in ecological conditions (altitude, rocks, soils, rainfall) and vegetation. For the study of individual ecosystems, the method of Herrera and Rodriguez (1988) was used, modifying the formula (L+F)/H to determine the overall rate of decomposition of the litter. It confirms that sclerophylly increases with the accentuation of stressful environmental conditions. The lowest values of the ratio (L+F)/H (eueremazimotic ecosystems) are observed in highly humid and very poor soils, in which there are major environmental stresses.
Key words: decomposition, functioning, litter, sclerophylly, tropical forests.
INTRODUCCIÓN
Desde hace mucho tiempo se conoce la importancia de lo que ocurre en el piso del bosque como parte integral del ecosistema. Uno de los principales procesos que tiene lugar en éste, es la descomposición, término empleado para referirse a todos los cambios biológicos, químicos y físicos que ocurren en el mantillo. Los ecólogos que se ocupan de estos estudios han prestado especial atención a la descomposición de la hojarasca, en relación con el ciclo de los nutrientes y con la productividad del suelo (Martín et al., 1994, 1996). Es evidente la relación entre la descomposición de la hojarasca y la disponibilidad de nutrientes, aspecto determinante en la producción de la biomasa total.
El presente estudio se enfoca en las relaciones ecológico-funcionales establecidas en los ecosistemas forestales, teniendo como antecedentes diferentes trabajos (Garrido et al., 1989; Prause & de Lifschitz, 2001; Prause et al., 2003; Ribeiro et al., 2004), fundamentalmente los realizados en Cuba, principalmente en la parte occidental del país (Rodríguez & Ulehlová, 1986; Rodríguez, 1988; Herrera & Rodríguez, 1988; Herrera et al., 1988); en la región oriental existen solamente los trabajos de Fornaris et al. (2000 a, b).
Cuba Oriental es la región más diversa con relación a altitud, clima, geología, suelos, flora, vegetación y fauna, condicionando gran variedad de formaciones vegetales (Reyes & Acosta, 2005 a, b; Reyes, 2006), es por ello que tiene especial importancia profundizar los conocimientos sobre las respuestas funcionales, con el fin de establecer estrategias de conservación y desarrollo de los bosques.
MATERIAL Y MÉTODOS
a) Características de las áreas de estudio
La región oriental presenta áreas climáticas desde menos de 500 mm de lluvia y 11 meses secos, hasta otras con más de 3 500 mm y 200 mm en el mes menos lluvioso, constituyéndose, de esta manera, en la zona más diversa del archipiélago cubano.
La zona costera suroriental, donde se encuentra el matorral costero y precostero, por estar protegida de los vientos alisios por las montañas, sufre el efecto de una acentuada sombra pluvial, contribuyendo esto a que en la mayor parte del área se observan condiciones extremas [temperaturas de 24 a 26°C; radiación solar muy intensa; poca lluvia (500-800 mm) con distribución irregular (9-11 meses secos) (Montenegro, 1991a, b, c; Lapinel, 1989); elevada evaporación (Crespo, 1989) y vientos marinos; además, hay lapiez o rendzinas muy superficiales, asi como calizas u otros materiales muy percolantes.
En la parte alta de la Sierra Maestra, donde se presentan el bosque nublado (1 500 m.s.n.m. 0 más) con suelos ferralíticos amarillentos lixiviados con una CIC (valor de T) de 2.5 a 18.75 meq/100 g y la CCB (valor de S) generalmente varía entre 1.44 y 5.02 meq/100 g, y la pluvisilva montana (800 a 1 400 m.s.n.m.) con un suelo ferralítico rojo lixiviado, donde se observa una CIC (valor de T) entre 5.6 y 20.5 meq/100 g y la CCB (valor de S) varía entre 2.8 y 6 meq/100 g (Renda, 1989). Aquí llueve entre 1 600 y 2 000 mm, con un periodo menos lluvioso de noviembre a abril (Trusov et al., 1983) y con gran influencia de las precipitaciones horizontales (nieblas y nubes bajas) (Reyes et al., 2005). La temperatura media, con un gradiente de 0.62°C/100 m de elevación en la ladera norte y 0.66°C/100 m en la ladera sur (Montenegro, 1991a), fluctúa en la cima del Pico Turquino (1 972 m.s.n.m.) entre 13.2 y 13.8°C en enero y entre 17.9 y 18.3°C en julio.
Climáticamente el macizo Sagua Baracoa, parte noreste de Cuba oriental, tiene características peculiares. Desde el interior de las Alturas de Moa hasta el río Duaba, donde se presentan la pluvisilva submontana sobre suelos de mal drenaje [en ofiolitas, con suelos ferríticos amarillentos con una CIC (valor de T) de 1.46 meq/100 g, (Ruiz, 1988)] y la pluvisilva de baja altitud sobre rocas metamórficas (en tobas, con suelos ferralíticos rojo lixiviados, pobres y ácidos) caen de 3 400 a 3 800 mm de precipitación media, con años aislados de hasta 5000 mm (Montenegro, 1991d), es a su vez, la zona más lluviosa de Cuba. El bosque siempreverde mesófilo se caracterizó en el valle Palenque-Bernardo-Tribilin, con alrededor de 1 700 mm de lluvia y suelos pardos mesotróficos.
El bosque semideciduo micrófilo se estudió en las terrazas costeras, sobre margas y con alrededor de 800 mm de lluvia (Montenegro, 1991c), mientras el bosque semideciduo mesófilo, con alrededor de 1200 mm de precipitación media, en la base del Pico Turquino, ambas en el sur de la Sierra Maestra.
b) Metodología
Para el análisis de cada uno de los ocho ecosistemas estudiados se utilizó el método descrito por Herrera & Rodríguez (1988), donde se describen los rangos de cada parámetro (esclerofilia, esclerorrizia, estructura y altura de la vegetación, presencia de estera radical y descomposición de la necromasa); sin embargo, se utilizó una modificación de la fórmula usada por dichos autores L/(F+H) para conocer la velocidad general de la descomposición del mantillo, por la siguiente (L+F)/H, pues se considera que explica mejor la relación entre los componentes poco o parcialmente descompuestos y los ya transformados en humus.
Para la microfilia se usó una plantilla con las superficies de los tipos de hojas de acuerdo a Borhidi (1996). Para la esclerofilia se colectaron cinco muestras de hojas maduras sin peciolo, de cinco especies que proporcionan el 75% o más de la hojarasca (peso fresco, PF); se secaron hasta peso constante a una temperatura de 90 a 95°C (peso seco, PS), calculándose la relación peso seco/peso fresco (PS/PF). Para la esclerorrizia se tomaron cinco réplicas de raicillas de menos de 1 mm de grosor, se lavaron hasta eliminar el suelo adherido y se prensaron suavemente con papel de filtro; posteriormente se secaron obteniéndose la relación PS/PF. El valor de dicha relación se considera para ambos de la forma siguiente:

Para el mantillo se recolectaron entre cinco y siete muestras, con un marco de 0.25 x 0.25 m (0.0625 m2) ó 0.50 x 0.50 m (0.25 m2); la primera medida fue necesaria en ecosistemas de difícil acceso y una enorme cantidad de mantillo (bosque nublado y pluvisilva submontana sobre suelos de mal drenaje) y donde, por el lapiez, fue imposible obtener una superficie mayor (matorral costero y precostero), en los demás se utilizó la segunda medida. Se tuvo en cuenta que las colectas se hicieran en zonas con un dosel fisonómica y florísticamente lo más homogéneas posible.
Se tomó todo el material orgánico hasta llegar al suelo y se recogió cada fracción por separado, luego, mediante humedecimiento, se retiró el suelo adherido a la muestra. Se separaron las capas L [hojas enteras (HE), hojas fragmentadas (HF), flores, frutos y ramitas], F (fragmentos de hojas, FH) y H [humus bruto grueso (HBG) y humus bruto fino (HBF)]. Con los valores obtenidos se estudió la relación entre las fracciones con vistas a conocer la descomposición de una respecto a la otra, además se calculó el valor de (L+F)/H, el que se consideró de la siguiente forma:

RESULTADOS
Tamaño de las hojas
Se presentan hojas mesófilas en la pluvisilva de baja altitud sobre rocas metamórficas, con abundante pluviosidad y protegida del viento; en el bosque siempreverde mesófilo (mesofítico), con suelos ricos y precipitación favorable; en el bosque semideciduo mesófilo, con condiciones ecológicas medias y en la pluvisilva montana más protegida del viento, los demás ecosistemas son micrófilos. En el grupo de ecosistemas micrófilos se destaca el matorral costero y precostero, con nueve meses secos y lapiez y/o rendzinas; se observan también el bosque semideciduo micrófilo con suelos pardos y alrededor de 800 mm de precipitación anual; la pluvisilva submontana sobre suelos de mal drenaje de Monte Iberia con suelos extremadamente pobres y alrededor de 3 000 mm de lluvia y la pluvisilva montana, influenciada fuertemente por el viento.
Esclerofilia y esclerorrizia
Respecto a la esclerofilia (cuadro 1), se observa que los ecosistemas más tensionados (bosque nublado, matorral costero y precostero y pluvisilva submontana sobre suelos de mal drenaje) tienen hojas euesclerófilas. En los demás cinco ecosistemas, la disminución del valor de la esclerofilia está relacionada con el decremento de la tensión, debido al aumento de la humedad y el mejoramiento de las condiciones del suelo; en este grupo, el bosque semideciduo micrófilo es esclerófilo, aunque su valor está muy cercano a mesoesclerófilo, categoría que presentan el bosque semideciduo mesófilo y la pluvisilva montana en general. El bosque siempreverde es hipomesoesclerófilo con pocas tensiones y por último la pluvisilva de baja altitud sobre rocas metamórficas con más de 3 000 mm es oligoesclerófila. La esclerorrizia sólo pudo ser determinada en bosques con estera radical, por ello, éstas no se presentan donde faltan datos. Como se observa (cuadro 1), el bosque nublado es euesclerorrízico, cuyas cifras son las mayores encontradas en el archipiélago cubano, mientras que la pluvisilva submontana sobre suelos de mal drenaje es esclerorrízica. En la pluvisilva montana la esclerorrizia se comporta igual a la esclerofilia (mesoesclerorrízico), mientras el bosque siempreverde es oligoesclerorrízico. Los bosques en condiciones ecológicas favorables son fundamentalmente hipoesclerorrizicos, mientras que los que se presentan en condiciones estresantes son de mesoesclerorrizicos a euesclerorrízicos.

El mantillo
Como se observa (cuadro 2), según su cuantía, la capa L puede ser separada en dos grupos: los de mayores valores, con más de 1 000 g/m2, y los que alcanzan menor cantidad, con cifras sensiblemente más pequeñas. El primer grupo coincide con ecosistemas donde existen grandes tensiones edafoclimáticas (de diferentes tipos), por lo que en algún momento el proceso de descomposición de las primeras fracciones se ralentiza. A su vez, presentan mayores cifras, tanto de las ramitas (se considera que en la pluvisilva submontana sobre suelos de mal drenaje y en la pluvisilva de baja altitud sobre rocas metamórficas las grandes lluvias facilitan su caída), como de los componentes de más fácil descomposición (hojas, flores y frutos). Por el contrario, el grupo con valores más pequeños coincide generalmente con tensiones menos acentuadas y menor cantidad de ambos elementos.
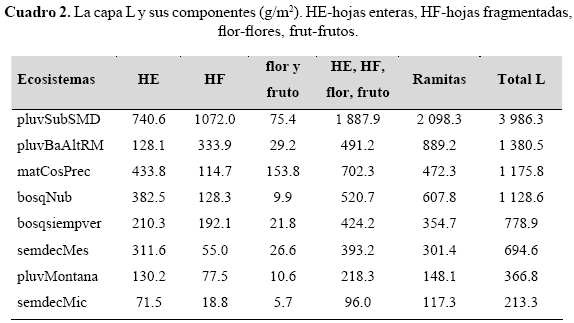
Las hojas enteras (HE) alcanzan cifras muy altas en los ecosistemas más tensionados (pluvisilva submontana sobre suelos de mal drenaje, matorral costero y precostero, bosque nublado), cuyas hojas son a su vez euesclerófilas. La mayor cuantía se observa principalmente en la pluvisilva submontana sobre suelos de mal drenaje, la que tiene 1.7 veces la del matorral costero y precostero que le sigue en cantidad y 5.8 veces la de la pluvisilva de baja altitud sobre rocas metamórficas (oligoesclerófila), relativamente cercana y con lluvias prácticamente semejantes (más de 3 000 mm). Con excepción del bosque semideciduo micrófilo, que es secundario y tiene muy poca cantidad de todos los elementos, la cantidad de hojas enteras disminuye en función de la esclerofilia.
Las hojas fragmentadas (HF) (cuadro 2), donde el proceso de descomposición está en pleno desarrollo, presentan los mayores valores (y por tanto constituyen una ralentización del proceso) en los ecosistemas situados en la zona más lluviosa del archipiélago cubano, pluvisilva submontana sobre suelos de mal drenaje (euesclerófila) y pluvisilva de baja altitud sobre rocas metamórficas (oligoesclerófila), donde, como ya se expuso, reciben más de 3 000 mm sin un periodo seco; se considera que el exceso de lluvia dificulta (independientemente de la esclerofilia) la descomposición de esta fracción (HF). En los demás ecosistemas, aunque a veces existen otros factores atenuantes, hay un periodo seco definido.
Las mayores cifras de las fracciones más descompuestas (FH+H), se encuentran en los ecosistemas más altamente tensionados (pluvisilva submontana sobre suelos de mal drenaje y bosque nublado), no sólo por la gran cantidad de humus (H) (tanto HBG como HBF), sino también por los fragmentos de hojas (FH) (cuadro 3). La pluvisilva submontana sobre suelos de mal drenaje, el ecosistema más tensionado, presenta 11.8 veces más de estos elementos (FH+H), 11.4 más de humus y 14.4 más de FH que la pluvisilva de baja altitud sobre complejo metamórfico; además, presenta 66.5 veces más de dichas fracciones (FH+H) y 190.8 del humus (H) que el bosque siempreverde con las menores tensiones ambientales. A su vez, el bosque nublado alcanza valores 30.9 veces mayores de estas fracciones (FH+H), 56.3 veces más de humus (H) y 3.1 veces más de FH que la pluvisilva montana, que es el ecosistema que queda inmediatamente debajo del mismo. La pluvisilva de baja altitud sobre complejo metamórfico, cuya principal tensión en el mantillo es el exceso de lluvia, es en este conjunto de fracciones (FH+H), 5.6 veces más y en humus (H) 16.7 que el bosque siempreverde expuesto.

Respecto a los fragmentos de hojas (FH), la mayor cantidad se encuentra en las zonas más tensionadas por la extrema pobreza del suelo, interrelacionada con los altos valores de humedad en el mantillo, sea por lluvia o porque presentan gran influencia de las precipitaciones horizontales (pluvisilva submontana sobre suelos de mal drenaje y bosque nublado). La primera tiene 14.4 y el segundo 2.5 veces más, respectivamente, de esta fracción, que la pluvisilva de baja altitud sobre complejo metamórfico, que le sigue en cantidad.
La mayor cuantía del humus bruto grueso (HBG), del humus bruto fino (HBF) y por consiguiente del humus total (H), se encuentra en los mismos ecosistemas y las mismas condiciones expuestas en los fragmentos de hojas (FH). Los menores valores, independientemente de la esclerofilia, se encuentran en sitios con un periodo seco acusado.
En la pluvisilva de baja altitud sobre rocas metamórficas, que como ya se expuso pertenece a la zona lluviosa y sin periodo seco, el HBG, el HBF y el H, es 13.6, 20.0 y 16.7 veces mayor respectivamente, que el bosque siempreverde, relativamente cercano, pero con la mitad de las precipitaciones y un periodo seco determinado.
Relación entre las fracciones
Cuando se estudia la velocidad en la descomposición del mantillo, en general, a través de la relación (L+F)/H, se observa que según la misma, los ecosistemas estudiados (cuadro 4), pueden dividirse en cuatro grupos: eueremazimóticos (descomposición muy lenta, menos de 1.00 en dicha relación), eremazimóticos (lenta, de 1.00 a 4.99), mesozimóticos (media, de 5.00 a 9.99) y taquizimóticos (rápida, más de 10.00). Los menores valores en esta relación, se presentan en los ecosistemas húmedos, sobre todo en los que se observan las mayores tensiones ambientales (pluvisilva submontana sobre suelos de mal drenaje y bosque nublado, eueremazimóticos).
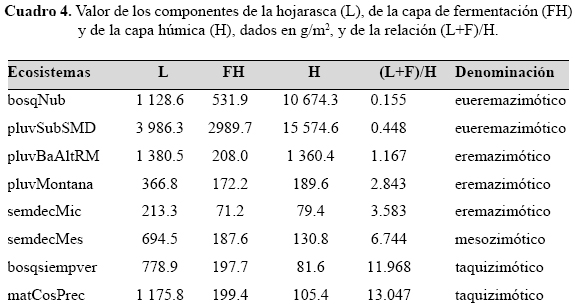
En general, no se presenta una relación directa con la esclerofilia, pues aunque los eueremazimóticos son euesclerófilos, se observan ecosistemas eremazimóticos, tanto oligoesclerófilos, mesoesclerófilos, como esclerófilos. A su vez, en los taquizimóticos, hay también euesclerófilos e hipomesoesclerófilos.
Al estudiar la relación HF/HE (cuadro 5) se encuentran valores mayores a la unidad, lo que expresa que en las hojas fragmentadas (HF) se produce una considerable ralentización del proceso de descomposición, en la pluvisilva submontana sobre suelos de mal drenaje y en la pluvisilva de baja altitud sobre rocas metamórficas, lo que coincide, como ya se expuso, con la zona de mayor pluviosidad del archipiélago cubano, sin época seca. A su vez, los valores menores se hallan en los bosques semideciduos, con la consiguiente caída de las hojas en una época seca definida.

Cuando se observa la relación FH/HF (cuadro 5), con excepción de la pluvisilva de baja altitud sobre rocas metamórficas, cuya ralentización del proceso de descomposición del mantillo se inicia en la hojas fragmentadas (HF), en los demás, dicha ralentización comienza o se intensifica en los fragmentos de hojas (FH), alcanzando los valores más elevados en el bosque nublado y los bosques semideciduos. Aunque las menores relaciones están en los ecosistemas con más baja esclerofilia, posteriormente se mezclan, lo que denota que no existe una relación directa entre ambos.
La mayor cuantía de la relación HBG/FH se observa en el bosque nublado (cuadro 5), que incluso sobrepasa en 4.1 veces a la pluvisilva de baja altitud sobre rocas metamórficas y en 5.7 veces a la pluvisilva submontana sobre suelos de mal drenaje, que le siguen en esta relación y que tienen cifras superiores a la unidad. A su vez, en los ecosistemas con cifras inferiores a la unidad, que significa más velocidad de descomposición que los fragmentos de hojas se observa un periodo seco, el menor valor de esta relación se encuentra en el bosque siempreverde el que presenta condiciones óptimas para la descomposición de esta fracción.
Por el contrario, en la relación HBF/HBG (cuadro 5), se observa que los valores superiores a la unidad, lo que representa ralentización del HBF, se encuentran en los ecosistemas con mayor cantidad de lluvia (pluvisilva submontana sobre suelos de mal drenaje y pluvisilva de baja altitud sobre rocas metamórficas) y sin época seca; a su vez, los otros seis ecosistemas, con cifras menores a la unidad y que significa mayor velocidad de descomposición que la otra fracción, tienen un periodo seco definido. Las dos proporciones, marcadamente menores, representan ecosistemas con condiciones particulares, el matorral costero y precostero presenta una extrema época de sequía (nueve meses), por lo que disminuye la formación del HBF y éste se incorpora rápidamente al suelo y la pluvisilva montana, donde debido a la gran pendiente, las lluvias arrastran el material más fino.
DISCUSIÓN
El funcionamiento de la vegetación depende del conjunto de condiciones ecológicas y define su estrategia de desarrollo y supervivencia. Herrera & Rodríguez (1988) expusieron un grupo de factores que actúan como tensiones sobre el bosque, de los que depende las características intrínsecas del mismo.
Microfilia, esclerofilia y esclerorrizia
Es generalmente aceptado que los tamaños y áreas foliares disminuyen a medida que se reduce la disponibilidad de humedad y de nutrientes (Lerch, 1965; Walter, 1970; Herrera et al., 1988; Boeger & Wisniewski, 2003). De los datos encontrados, se fortalece la idea que la microfilia es una consecuencia compleja y que el conjunto de factores edafoclimáticos actúan interrelacionados y no sólo la humedad, pues es micrófila la pluvisilva submontana sobre suelos de mal drenaje, con un suelo extraordinariamente pobre (1.4 meq/100 g, Ruiz, 1988), 3 000 mm de lluvia y sin época seca y es mesófila la pluvisilva de baja altitud sobre rocas metamórficas, con la misma cantidad de precipitaciones y suelos mejores. A su vez, en la pluvisilva montana Reyes et al. (2005) encontraron que debido a su acción desecante, el viento influye en el aumento de las especies con hojas micrófilas en el estrato arbóreo.
El conjunto de factores ambientales extremos que actúan de forma interrelacionada en la zona costera suroriental, provocan grandes tensiones sobre la vegetación, lo cual se manifiesta en la sencilla estructura y la acusada microfilia de las comunidades costeras y cercanas a la costa (matorral costero y precostero, bosque semideciduo micrófilo). En los demás bosques micrófilos, actúa principalmente la poca profundidad del suelo y en lugares expuestos el viento. La esclerofilia y la esclerorrizia son características importantes de la vegetación y expresan la interrelación del conjunto de factores que influyen en el ecosistema. Según Esau (1969) las hojas más ricas en esclereidas son más duras y esclerófilas, por lo que entonces la esclerofilia (PS/PF) es mayor. Jordan (1989) expone que las hojas escleromórficas ocurren frecuentemente en bosques tropicales pobres en nutrientes. Además, el déficit hídrico y nutrimental pueden actuar de conjunto (Santiago Martín et al., 2001). Como se observa (cuadro 1), hay coincidencia con estas aseveraciones (Fornaris et al., 2000 a, b), los bosques con hojas predominantemente euesclerófilas (bosque nublado, matorral costero y precostero y la pluvisilva submontana sobre suelos de mal drenaje), son los ecosistemas con mayores tensiones, tanto edáficas como climáticas, mientras en los demás disminuye la esclerofilia en función de la reducción del estrés ambiental. En otros bosques anteriormente estudiados en Cuba (Herrera & Rodríguez, 1988) también se encuentran estas características y denota la complejidad de estas interrelaciones.
También la dureza de las raicillas (esclerorrizia) depende de las tensiones a que está sometido el ecosistema, sobre todo el subsistema radical (Herrera & Rodríguez, 1988). Esto se comprueba en los datos expuestos, donde dichas tensiones aumentan desde el bosque siempreverde mesófilo (mesofítico y oligoesclerorrízico) sobre suelos ricos (Fornaris et al., 2000 a), pasando por suelos pobres y condiciones ecológicas moderadas (pluvisilva montana), suelos extraordinariamente pobres (Ruiz, 1988) y exceso de lluvia (pluvisilva submontana sobre suelos de mal drenaje), hasta el conjunto de tensiones ambientales del bosque nublado, donde también hay suelos muy pobres; estos dos últimos con muy bajo grado de saturación de bases.
El mantillo y su descomposición
Al estudiar las hojas enteras (HE), se constata que alcanzan valores superiores en los ecosistemas euesclerófilos, ello ratifica lo expuesto por Herrera & Rodríguez (1988), que las hojas con mayor esclerofilia presentan más dificultad en descomponerse y en ser utilizadas por la meso y microbiocenosis. Ello se produce independientemente de la humedad, ya que tanto lugares áridos (matorral costero y precostero) como muy lluviosos (pluvisilva submontana sobre suelos de mal drenaje) se comportan de forma semejante. Además, la disminución de las cifras de hojas enteras (HE) en función de la esclerofilia, demuestra que es en esta fracción que se cumple lo expuesto por dichos autores.
En las hojas fragmentadas (HF), las que ya se encuentran reblandecidas y están siendo usadas como fuente de energía, se ratifica lo expuesto por Rodríguez & Ricardo (1983) de que un periodo seco es importante en la descomposición de la hojarasca.
En este estudio se corrobora que en el grupo de fracciones donde el proceso de descomposición es más avanzado (FH + H), el conjunto de tensiones edafoclimáticas, interrelacionadas, definen dicha descomposición, pues éste es más lento y por tanto hay mayor cantidad de mantillo, a medida que el estrés ambiental es más acentuado (pluvisilva submontana sobre suelos de mal drenaje y bosque nublado). La extrema carencia en nutrientes en el suelo, así como el exceso de humedad, sea por la gran cantidad de lluvia con ausencia de periodo seco, como por la influencia de las precipitaciones horizontales sobre la gran cantidad del mantillo, dificultan el trabajo de la meso y la microbiocenosis; constituyendo a su vez las principales tensiones sobre la descomposición del mantillo en estos ecosistemas tropicales.
Se confirma, por las cifras de estas fracciones en los cinco últimos ecosistemas (cuadro 3), que son apreciablemente menores, que la existencia de un periodo seco definido, como fue anteriormente expuesto, facilita la descomposición de dichas fracciones.
El proceso de descomposición del mantillo, el que es extraordinariamente complejo (Lupi et al., 2002; Prause et al., 2003; Petisco et al., 2005), forma parte de la dinámica o funcionamiento de la vegetación y depende de la interacción de un conjunto de factores abióticos y bióticos.
Como bien han fundamentado Singh & Singh (1987), más tarde descrito por Gallardo (1994), al parecer, el alto contenido de lignina es la razón por la cual ocurre el retardo en la descomposición de los fragmentos de hojas (FH). A este componente de la hojarasca es al que algunos autores han denominado fracción recalcitrante (Álvarez-Sánchez & Becerra, 1996; Fornaris et al., 2000a) debido a lo difícil de su descomposición. Se considera, que en las condiciones de este estudio, la dificultad en la descomposición de dichos fragmentos de hojas (FH), se debe principalmente a la falta de aireación por el exceso de lluvia o de precipitaciones horizontales; Del Valle-Arango (2003) observó una relación entre la anaerobiosis y la lentitud de la descomposición.
Cuando se analizan las cifras del componente húmico (H), se observan cuestiones muy interesantes. Los menores valores se encuentran en los ecosistemas de lugares más secos, aumentando dicho componente a medida que el exceso de humedad se constituye en un factor limitante. Como las capas húmicas están debajo de las L y FH, es más fácil conservar un exceso de humedad, ello se confirma en la pluvisilva submontana sobre suelos de mal drenaje y en el bosque nublado.
La relación entre las distintas fracciones del mantillo expresa las complejidades de la descomposición y permite detectar donde dicho proceso se ralentiza y/o se acelera y por lo tanto analizar sus causas. Herrera & Rodríguez (1988) expusieron que en general, la velocidad de descomposición del mantillo dependía de la esclerofilia de las hojas. Sin embargo, este proceso es muy complejo y depende de un conjunto de factores que no pueden ser determinados totalmente en este trabajo. Ejemplo, son eremazimóticos un conjunto de ecosistemas que varían desde más de 3 000 mm de lluvia y ningún mes con menos de 200 mm, suelos ácidos y pobres y oligoesclerófilos, hasta con alrededor de 900 mm, seis meses secos, suelos ricos en carbonatos y esclerófilos. A su vez, son taquizimóticos el bosque siempreverde, muy poco tensionado, con lluvias favorables, tres a cuatro meses secos, suelos mesotróficos e hipomesoesclerófilo y el matorral costero y precostero, altamente tensionado, con alrededor de 700 mm y nueve meses secos, elevadas temperaturas, creciendo sobre el lapiez o diente de perro y euesclerófilo. Sin embargo (cuadro 4), los cuatro ecosistemas con valores menores de la relación (L+F)/H (eremazimóticos y eueremazimóticos) constituyen sitios con alta precipitación, sin meses secos y/o zonas elevadas (a partir de los 800 m.s.n.m.) con lluvias favorables, poca evaporación y precipitaciones horizontales la mayoría de los días del año. Los cuatro ecosistemas con mayor velocidad en la descomposición, tienen periodos secos definidos, a veces acusados, cuya importancia fue anteriormente expuesta. Respecto al proceso de descomposición en general (L+F)/H, se observa que la velocidad del proceso desciende con el aumento de la humedad y la intensificación de la pobreza del suelo. Es conocido (Martín et al., 1993) que dicha descomposición se realiza con mayor intensidad cuando hay una humedad apropiada (Zech & Kogel-Knabner, 1994) y en suelos ricos, retardándose con la baja disponibilidad de nutrientes (Martín et al., 1993; Carmona et al., 2006; Graça & Canhoto, 2006); ello fortalece el criterio de que la tensión nutrimental tiene gran efecto sobre el ecosistema (Herrera et al., 1988; Jordán, 1989).
Se considera que en las condiciones tropicales los principales factores que determinan la lentitud del proceso de descomposición son: el exceso de humedad, la pobreza de nutrientes, el pH muy bajo y la alta esclerofilia; hay otros que actúan localmente, como las bajas temperaturas en las montañas elevadas. Los mismos actúan limitando la actividad de los microorganismos y la fauna que participan en la descomposición del mantillo. Debido a la gran cantidad e intensidad de esos factores, los menores valores de la relación (L+F)/H se observan en el bosque nublado y en la pluvisilva submontana sobre suelos de mal drenaje (0.155 y 0.448 respectivamente), en los cuales el proceso de descomposición es muy lento; estos valores son los menores encontrados hasta el momento en Cuba (Herrera & Rodríguez, 1988; Fornaris et al., 2000 a, b; Reyes et al., 2005; Matos, 2007).
Cuando se examina (cuadro 5) la relación HF/HE, se observa también que el exceso de humedad del mantillo, producto de las continuas lluvias, independientemente de la esclerofilia, provoca una relación mayor que la unidad, es decir, ralentización en la descomposición de las hojas fragmentadas (HF).
El alto valor de la relación HBG/FH en el bosque nublado (más de 1 500 m.s.n.m.), c onfirma que las bajas temperaturas ralentizan el proceso de descomposición, lo que también fue observado por Varela et al. (2002), y que para que éste se desarrolle de forma óptima, es preciso una temperatura y humedad adecuadas (Martín et al., 1993, 1996). Además, en los valores de la relación expuesta y en los de la HBF/HBG de los ecosistemas con mayor cantidad de lluvia, se observa mayor lentitud relativa en la descomposición de ambas fracciones.
En el proceso de descomposición del mantillo se le ha brindado poca importancia a las ramitas; en algunos ecosistemas ellas representan mayor peso que la parte foliar (cuadro 3), varios autores han trabajado en ello (Menéndez, 1990; Lastres, 1988; Álvarez-Sánchez & Guevara, 1993; Muñoz-Cruz & Álvarez-Sánchez, 1995), debiéndose intensificar estos estudios, completando así los conocimientos sobre este proceso.
CONCLUSIONES
Se ratifica que el valor de la esclerofilia y la esclerorrizia aumentan en función de las tensiones a que es sometido el ecosistema, principalmente el déficit hídrico y/o la pobreza en nutrientes del suelo.
El proceso de descomposición del mantillo es extraordinariamente complejo. Tanto en la capa L, como en la FH y la H, las mayores cifras de biomasa se encuentran en los ecosistemas con más tensiones edafoclimáticas.
Respecto al proceso completo de descomposición, o sea, la relación (L+F)/H, se encuentran valores menores a la unidad (ecosistemas eueremazimóticos) en áreas muy húmedas, combinadas con la extrema pobreza del suelo; mientras las cifras más elevadas de dicha relación (ecosistemas taquizimóticos) se encuentran en suelos ricos, con una humedad óptima y una corta estación menos lluviosa.
El exceso de humedad (tanto por la gran cantidad de lluvia sin un periodo seco, como por la influencia de las precipitaciones horizontales) que produce anaerobiosis, constituye el principal factor en la ralentización de la descomposición del mantillo en estos ecosistemas tropicales.
LITERATURA CITADA
Álvarez-Sánchez, J. & S. Guevara, 1993. "Litterfall Dinamics in a Mexican Lowland Tropical Rain Forest". In Tropical Ecology, 34(2): 127-142. [ Links ]
Alvarez-Sánchez, J. & R. Becerra, 1996. "Leaf Decomposition in a Mexican Tropical Rain Forest". Biotropica, 28(4b): 657-667. [ Links ]
Boeger, M.R.T. & C. Wisniewski, 2003. "Comparação da morfología foliar de espécies arbóreas de três estadios sucessionais distintos de floresta om-brófila densa (Floresta Atlântica) no Sul do Brasil". Revista Brasileira de Botânica, 26(1):10 pp. [ Links ]
Borhidi, A., 1996. Phytogeography and Vegetation Ecology of Cuba. 2a ed. Budapest. Akadémiai Kiadó. 926 pp. [ Links ]
Carmona, M.R., M. Aguilera, C.A. Pérez & I. Serey, 2006. "Actividad respiratoria en el horizonte orgánico de suelos de ecosistemas forestales del centro y sur de Chile". Gayana Botanica, 63(1):1-12. [ Links ]
Crespo, S.E., 1989. "Evaporación media anual". En Nuevo Atlas Nacional de Cuba. Mapa 42. pp. VI.4.1. [ Links ]
Del Valle-Arango, J.I., 2003. "Descomposición de la hojarasca fina en bosques pantanosos del pacífico colombiano". Interciencia, 28(3):148-153. [ Links ]
Esau, K., 1969. Anatomía vegetal. Ed. Revolucionaria. Instituto Cubano del Libro. La Habana. 390 pp. [ Links ]
Fornaris, E., O.J. Reyes & F. Acosta, 2000a. "Características fisonómicas y funcionales de la Pluvisilva de baja altitud sobre complejo metamórfico de la zona nororiental de Cuba". Biodiversidad de Cuba Oriental 4, Editorial Academia. pp. 44-51. [ Links ]
----------, 2000b. "Características fisonómicas y funcionales del Bosque siempreverde mesófilo de las Cuchillas del Toa, Guantánamo, Cuba". Biodiversidad de Cuba Oriental 4, Editorial Academia. pp. 36-43. [ Links ]
Gallardo, J.F., 1994. "Dinámica de la descomposición orgánica en sistemas conservacionistas". Memorias VII Congreso. España. pp. 31-37. [ Links ]
Garrido, M.V., J.M. Arco & J. Escudero, 1989. "Ciclo de nutrientes: fenología de abscisión y dinámica de la descomposición en distintas especies caducifolias y perennifolias". Options Méditerrannéenes. Séries Séminaires, 3: 163-166. [ Links ]
Graça, M.A.S. & C. Canhoto, 2006. "Leaf litter processing in low order streams". Limnetica, 25(1-2): 1-10. [ Links ]
Herrera, R.A. & M.E. Rodríguez, 1988. "Clasificación funcional de los bosques tropicales". En Herrera, R.A., Menéndez, L., Rodríguez, M.A., García, E.E. Ecología de los bosques siempreverdes de la Sierra del Rosario, Cuba. Montevideo. ROSTLAC. pp. 574-626. [ Links ]
Herrera. R.A., M.E. Rodríguez, E. Furrazola, E.E. García, R.P. Capote & M. Ruiz, 1988. "Génesis y significación ecológica de las esteras radicales en los bosques tropicales". In Memorias del Primer Simposio de Botánica. Habana. vol. 5. pp. 63-89. [ Links ]
Jordan, C.F. Ed., 1989. "An Amazonian Rain Forest". The Structure and Function of a Nutrient Stressed Ecosystem and the Impact of Slash and Burn Agriculture. Man and Biosphere Series. Vol. 2. 176 pp. [ Links ]
Lapinel, B., 1989. "Temperatura media anual del aire en julio". En Nuevo Atlas Nacional de Cuba. Mapa 17. pp. VI.2.4. [ Links ]
Lastres, O., 1988. "Dinámica de las reservas orgánicas y energéticas de la hojarasca en un bosque tropical semideciduo en Cuba". Tesis en opción a Doctor en Ciencias Biológicas. Instituto de Ecología y Sistemática. 182 pp. [ Links ]
Lerch, G., 1965. Pflanzenòkologie. Akademie Verlag, Berlin. 171 pp. [ Links ]
Lupi, A.M., M. Conti & R. Fernández, 2002. "Evolución de las fracciones de la MO en diferentes sistemas de manejo de residuos forestales". En Memorias XVIII Congreso Argentino de la Ciencia del Suelo. 5 pp. [ Links ]
Martín, A., J.F. Gallardo & I. Santa Regina, 1993. "Dinámica de la descomposición de hojas de rebollo en cuatro ecosistemas forestales de la Sierra de Gata (Provincia de Salamanca, España): Índices de descomposición". Invest. Agr. Sist. Recur. For., 2(1):5-17. [ Links ]
Martín, A., I. Santa Regina & J.F. Gallardo, 1994. Ciclos biogeoquímicos en un bosque perenne de encina (Quercus rotundifolia) en las proximidades de Salamanca (España): Retorno potencial. Consejería Med. Amb. Junta Castilla y León. Valladolid. pp. 151-160. [ Links ]
Martín, A., J.F. Gallardo & I. Santa Regina, 1996. "Dinámica de las Fracciones Orgánicas en la Descomposición de Hojas de Rebollos y Castaño". Silva Lusitana, 4(2):199-216. [ Links ]
Matos, R., 2007. "Funcionamiento Ecológico de la pluvisilva submontana sobre suelos de mal drenaje de Monte Iberia". Tesis en opción al título de Ingeniero Forestal. Ministerio de Educación Superior. Centro Universitario de Guantánamo. Facultad Agroforestal de Montaña. [ Links ]
Menéndez, L., 1990. "Estructura y biomasa del bosque siempreverde tropical de la Sierra del Rosario". Tesis Doctoral Ciencias Biológicas. Inst. Ecología y Sistemática. La Habana, Cuba. [ Links ]
Montenegro, U., 1991a. "Temperatura media anual". En Atlas de Santiago de Cuba. Academia de Ciencias de Cuba. Mapa 26. [ Links ]
----------, 1991b. "Insolación media periodo seco". En Atlas de Santiago de Cuba. Academia de Ciencias de Cuba. Mapa 25. [ Links ]
----------, 1991c. "Precipitación media anual". En Atlas de Santiago de Cuba. Academia de Ciencias de Cuba. Mapa 33. [ Links ]
----------, 1991d. Condiciones climáticas de las cuencas de los ríos Toa y Duaba de la provincia de Guantánamo. Inst. Meteorología, ACC, Santiago de Cuba. 45 pp. [ Links ]
Muñoz-Cruz, M. & J. Álvarez-Sánchez, 1 9 9 5. "Branch fall variation and over time in a mexican Rain Forest. UNAM". Brenesia, 43-44: 1-7. [ Links ]
Petisco, C., A. García, B.R. Vázquez, I. Zabalgogeazcoa, S. Mediavilla & B. García, 2005. "Determinación de lignina y celulosa en hojas de plantas leñosas mediante NIRS: Comparación de métodos estadísticos. Producciones Agroganaderas: Gestión eficiente y conservación del medio natural" (vol. I). XLV Reunión Científica de la SEEP (Sesión: Producción Animal). pp. 97-104. [ Links ]
Prause, J. & A.P. de Lifschitz, 2001. "Constantes de descomposición de hojas y la relación lignina/celulosa en cuatro especies forestales de la Reserva Estricta de Colonia Benítez-Chaco". Cátedra de Edafología. Facultad de Ciencias Agrarias. UNNE. Argentina. [ Links ]
Prause, J., A.P. de Lifschitz & D.M. Toledo, 2003. "Dinámica de la mineralización de N, P, y K en hojas de Schinopsis balansae Engl. sobre un suelo forestal del Parque Chaqueño Húmedo". Quebracho, 10: 46-52. [ Links ]
Renda, A., 1989. "Particularidades edafológica-forestales de la región central de la Sierra Maestra. La Habana". Tesis Doctoral Ciencias Agrícolas, ISCA Habana. [ Links ]
Reyes, O.J., 2006. "Clasificación de la vegetación de la Sierra Maestra". Biodiversidad de Cuba Oriental, Vol. VIII. pp. 23-41. [ Links ]
Reyes, O.J. & F. Acosta Cantillo, 2005a. "Vegetation. Cuba: National Park La Bayamesa". In Rapid Biological Inventories: 13. D. Maceira, A. Fong, W. S. Alverson & T. Wachter, Eds. pp. 113-119 and Appendice 150-161. [ Links ]
Reyes, O.J. & F. Acosta Cantillo, 2005b. "Vegetation. Cuba: Alejandro de Humboldt National Park". In Rapid Biological Inventories: 14. D. Maceira, A. Fong, W. S. Alverson & T. Wachter, Eds. pp. 158-172. [ Links ]
Reyes. O.J., O. Pelicié, C. Vinent, L. Mireya, E. Semanat & E. Fornaris, 2005. "Estudio fisonómico y funcional de las Pluvisilvas Montanas de la Gran Piedra". Foresta Veracruzana, 7(2):7-14. [ Links ]
Ribeiro, M.D., M. Álvarez, P. Riolobos & S. Cirujano, 2004. "Descomposición de helófitos en un humedal semiárido hipertrófico". Anales del Jardín Botánico de Madrid, 61(1): 53-61. [ Links ]
Rodríguez, M.E., 1988. "Ciclo de los nutrientes y estrategias para su conservación en el ecosistema". En Ecología de los Bosques Siempreverdes de la Sierra del Rosario. Montevideo. ROSTLAC. pp. 709-731. [ Links ]
Rodríguez, M.E. & N. Ricardo, 1983. "Descomposición de la hojarasca en tres lugares del Bosque Siempreverde de la Estación Ecológica Sierra del Rosario, Provincia Pinar del Río, Cuba". Ciencias Biológicas, 9: 55-65. [ Links ]
Rodríguez, M.E. & B. Ulehlová, 1986. "Ciclo de los macronutrientes en el sistema detritus-suelo en dos ecosistemas de bosque en Sierra del Rosario". Rev. Jardín. Bot. Nac. Univ. Habana, 7(1): 63-72. [ Links ]
Ruiz, J., 1988. "Particularidades de la formación y uso agrícola de los suelos del macizo montañoso Sagua Baracoa. La Habana". Tesis doctoral C. Agrícolas, Inst. Suelos, MINAGRI. 36 pp. [ Links ]
Santiago Martín, R., F. Olivares Ponce, C. Sánchez Sánchez & J. Arroyo Hailuoto, 2001. Bosques Mediterráneos. Ed. Depto. de Ecología de la Fac. Ciencias Biológicas de la Univ. Complutense de Madrid. 10 pp. [ Links ]
Singh, J.S. & S.P. Singh, 1987. "Forest Vegetation of the Himalaya". In: The Botanical Review. The New York Botanical Garden, 53(1):80-191. [ Links ]
Trusov, J.J., A. Izquierdo & L.R. Díaz, 1983. Características espaciales y temporales de las precipitaciones atmosféricas en Cuba. La Habana. Academia de Ciencias de Cuba, Inst. Geografía. 150 pp. [ Links ]
Varela, A., P. Barriga & J.A. Ahumada, 2002. "Comparación de factores abióticos relacionados con la descomposición de hojarasca entre fragmentos y no fragmentos de bosque Altoandino nublado (Sabana de Bogotá, Colombia)". Ecotrópicos, 15(2): 185-193. [ Links ]
Walter, H., 1970. Vegetationszonen und Klima. VEB Gustav Fisher Verlag Jena. 244 pp. [ Links ]
Zech, W. & I. Kogel-Knabner, 1994. "Patterns and regulation of organic mater transformation in soil: litter decomposition and humification". In Flux control in biological systems. Schulze, E.D. (Ed.) Academic Press, San Diego. pp. 303-334. [ Links ]

