










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkPolibotánica
versão impressa ISSN 1405-2768
Polibotánica no.31 México Mar. 2011
Caracterización morfológica de Padina boergesenii (Dictyotaceae, Phaeophyceae) en la costa mexicana del golfo de México y mar caribe
Morphological characterization of Padina boergesenii (Dictyotaceae, Phaeophyceae) on the mexican Coast of the Gulf of Mexico and Caribbean sea
Alejandrina G. Ávila-Ortiz1, Luz Elena Mateo-Cid2 y A. Catalina Mendoza-González2
1 Herbario FEZA. Facultad de Estudios Superiores Zaragoza, UNAM. Batalla 5 de Mayo y Fuerte de Loreto. Col. Ejército de Oriente C.P. 09230 México, DF. Correo electrónico:
2 Departamento de Botánica, Escuela Nacional de Ciencias Biológicas, IPN. Prolongación de Carpio y Plan de Ayala, Col. Santo Tomás. CP 11340 México, DF.
Correspondencia:
agao@xanum.uam.mx
Recibido: 10 diciembre 2009.
Aceptado: 20 septiembre 2010.
Resumen
Se realizó la caracterización morfológica de Padina boergesenii Allender y Kraft para México, con base en el estudio de los talos esporofitos y gametofitos masculino y femenino, estos últimos se reconocen por primera vez en el litoral mexicano del Golfo de México y Mar Caribe. Se revisaron un total de 78 ejemplares, 47 del herbario ENCB, 30 del herbario FEZA y uno del UAMIZ. Los especímenes correspondieron a 14 localidades litorales y 20 insulares de Veracruz, Campeche, Yucatán y Quintana Roo, México, recolectados durante los años 1960 a 2007. Del total de ejemplares examinados, el 95% fueron esporofitos, el 5% gametofitos, dos femeninos y uno masculino, así como un talo con ambos tipos de gametangios de Isla Chica, Yucatán. Este último hallazgo sugiere que esta especie puede ser dioica o monoica. La morfología del talo de Padina boergesenii es variable, ya que en localidades litorales crece generalmente en la zona intermareal y las láminas tienden a ser divididas; mientras, que en localidades insulares también habita la zona intermareal y las láminas son enteras o poco divididas.
Finalmente, este taxón se distribuye desde Veracruz hasta Quintana Roo, México.
Palabras clave: Dictyotales, esporofito, gametofito, morfología.
Abstract
A morphological characterization of Padina boergesenii Allender & Kraft was carried out based on variation in sporophytes and gametophytes of male and female plants from Mexican populations from the Gulf of Mexico and Caribbean Sea. We analyzed 78 herbarium specimens from ENCB (47 vouchers), FEZA (30) and UAMIZ (1). The specimens were taken during 1960 to 2007 at 14 coastal and 20 insular localities from Veracruz, Campeche, Yucatan and Quintana Roo. Of the specimens examined 95% were sporophytic thalli; only 5% were gametophytic, two female and one male. The one thallus seen with both types of gametangia came from Isla Chica, Yucatan, suggesting that P. boergesenii can have monoecious or dioecious thalli. In coastal localities the thalli become shallowly split into fan-shaped segments, whereas in insular localities they are semicircular with entire margins and only rarely split into segments. Finally, this taxon, which inhabits the intertidal zone, is distributed in Mexico from Veracruz to Quintana Roo.
Key words: Dictyotales, gametophyte, morphology, sporophyte.
INTRODUCCIÓN
El género Padina es un alga parda (Dictyotaceae, Phaeophyceae) con distribución pantropical (Womersley, 1987). Para las costas mexicanas del Golfo de México y Mar Caribe se han registrado seis especies: P. boergesenii Allender et Kraft, P. gymnospora (Kützing) Sonder, P. haitiensis Thivy, P. pavonica (Linnaeus) Thivy, P. perindusiata Thivy y P. sanctae-crucis Børgesen (Ortega et al., 2001). La diferenciación entre las especies mencionadas se basa en el número de capas de células medulares del talo, grosor de la lámina en el margen, zona media y basal, disposición de los soros esporangiales en uno o en ambos lados de la lámina, así como su relación con los pelos feofíceos. En la mayoría de las diagnosis de estas especies se incluye información sobre los soros esporangiales, pero no así de los gametangiales (oogoniales y anteridiales). Sin embargo, es común que los caracteres, sobre todo los vegetativos se empalmen, lo que crea confusión al momento de la determinación taxonómica de las especies (Taylor, 1960; Littler y Littler, 2000). En este sentido, en el presente estudio se proporciona información sobre la variación morfológica de Padina boergesenii, basada en la comparación de talos recolectados en diferentes ambientes y localidades litorales e insulares del Golfo de México y Mar Caribe. Además, se registran por primera vez los talos gametofitos femenino y masculino para P. boergesenii.
Padina boergesenii se caracteriza por la presencia de dos a tres capas de células a todo lo largo de la lámina, organizada en una capa de células medulares y dos corticales. Además posee otras característica del Orden Dictyotales como talos parenquimáticos, con crecimiento marginal y células con plastos numerosos y discoidales sin pirenoides, su ciclo de vida es digenético, isomórfico con alternancia entre un gametofito haploide y un esporofito diploide, reproducción sexual oogámica (Graham y Wilcox, 2000). Presenta una distribución amplia en el litoral del Golfo de México y Mar Caribe mexicano (Mendoza-González et al., 2000, Ortega et al., 2001, = P gymnospora (Kützing) Vickers = Huerta, 1961; Huerta y Garza-Barrientos, 1964; Díaz-Martín et al., 1998).
La mayoría de las especies de Padina son dioicas, entre ellas: P. antillarum (Kützing) Piccone (Wynne y De Clerck, 1999), P. crispata Thivy, P. durvillei Bory de Saint Vincent, P. mexicana Dawson (Ávila-Ortiz, 2003; Avila-Ortiz y Pedroche, 2005), P. gymnospora (Kützing) Sonder, P. sanctae-crucis Bergesen y Padina boergesenii (Allender y Kraft, 1983; Taylor, 1960; Littler y Littler, 2000). Solamente se ha descrito a P. australis Hauck (Abbott y Huisman, 2004) y P. mexicana Dawson (Lawson y John, 1987) como monoicas. Un caso particular es el de Padina pavonica, que fue registrada como dioica y monoica por Gómez-Garreta et al. (2007). Sin embargo, aún se desconocen los talos gametofitos fértiles en otras especies: P. crassa Yamada, P. ele-gans Kho ex Womerley, P. fraseri (Greville) Greville, P. haitiensis Thivy, P. japonica Yamada, P. minor Yamada, P. perindusiata Thivy (Womersley, 1987), P. profunda. Earle y P. boryana Thivy (Chapmann, 1963; Dawson et al., 1964; Gaillard, 1967, 1975; Fagerberg y Dawes, 1973, Lawson y John, 1987; Phillips, 1988; Trono, 1997 y Abbott y Huisman, 2004).
El presente estudio tiene como objetivo caracterizar la morfología de los esporofitos y evidenciar la presencia tanto de los gametofitos femenino y masculino de Padina boergesenii, además de confirmar y complementar la distribución de esta especie en el Golfo y Caribe mexicano.
MATERIAL Y MÉTODO
Se revisaron un total de 78 ejemplares fértiles y maduros de Padina boergesenii, 50 depositados en los herbarios de la Escuela Nacional de Ciencias Biológicas (ENCB), Facultad de Estudios Superiores Zaragoza (FEZA) y de la Universidad Autónoma Metropolitana-Unidad Iztapalapa (UAMIZ) (Holgremm et al., 1990), 28 fueron recolectados, durante este trabajo, de 2004 a 2007, los especímenes fueron desprendidos del sustrato con una espátula y fijados en formol al 4%. Los ejemplares datan de 1960 a 2007 (tabla 1), los cuales corresponden a 14 localidades litorales y 20 insulares de los estados de Veracruz, Campeche, Yucatán y Quintana Roo, México (tabla 2, fig. 1). Los datos de fecha, ubicación, número de herbario, número de colecta, hábitat y datos reproductivos se indican en la (tabla 1).

De los 78 ejemplares, se tomaron muestras del margen, zona media y basal de la lámina, los cuales fueron rehidratados para hacerles cortes histológicos en sentido longitudinal con una navaja de doble filo, éstos fueron teñidos con verde yodo para evidenciar la presencia del indusio y apreciar los soros maduros en los esporofitos, se midieron los esporangios en división, en el caso de los soros anteridiales se tomó en cuenta los anteridios más desarrollados que por lo general se ubican en el centro del soro, en el caso del soro oogonial las medidas se tomaron de manera similar, descartando en todos los casos los talos inmaduros o vegetativos (Ávila-Ortiz y Pedroche, 2005), con los cortes se elaboraron preparaciones semipermanentes con miel Karo® y fenol. De cada ejemplar se tomaron cinco medidas de cada uno de los caracteres vegetativos y reproductivos que definen a esta especie. Para fotografiar las estructuras más representativas se utilizó un microscopio Olympus CX31 y una cámara digital Canon®.
RESULTADOS
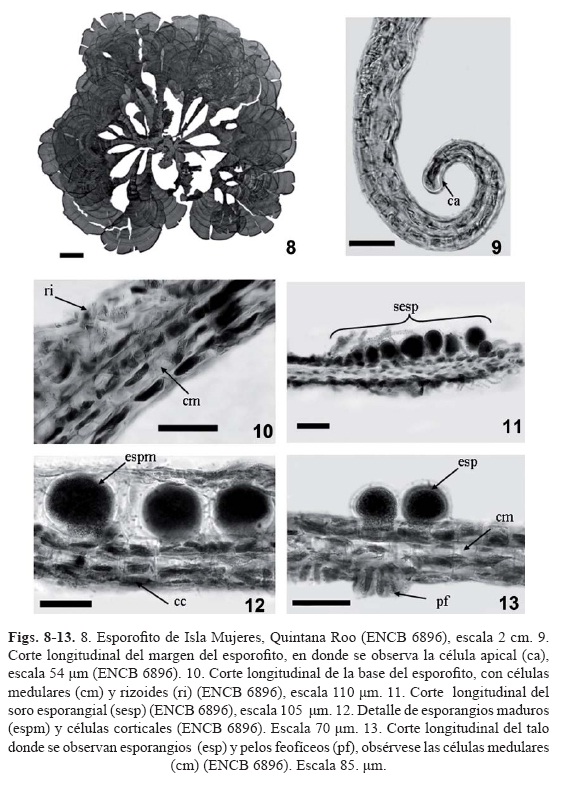
Los ejemplares revisados presentaron las características vegetativas y reproductivas de Padina boergesenii propuesta por Thivy (1959) y Allender y Kraft (1983), el carácter distintivo de esta especie es la presencia de dos a tres capas de células a lo largo de la lámina, ésta es la principal característica que distingue a P. boergesenii de las otras especies registradas en la zona de estudio. De los 78 especímenes, 74 fueron esporofitos (94.87%) y 4 gametofitos (5.13%), dos de ellos con oogonios, uno con anteridios y otro con oogonios y anteridios. Los gametofitos fértiles de Padina boergesenii solamente fueron observados en ejemplares de herbario provenientes de las islas Cozumel, Mujeres, Quintana Roo e Isla Chica, Yucatán (tabla 1, figs. 2-13 (3, 4, 5, 6, 7, 8, 9, 10, 11, 12)).

Descripción
Padina boergesenii Allender & Kraft 1983: 87-88
Localidad Tipo: Islas Vírgenes
Gametofito masculino figuras 2-3
Talo de 4 cm de largo, lámina lobada a dividida de 2 a 4 cm de ancho, color café claro, estípite de 0.5 cm de longitud. Ligera presencia de carbonato de calcio en la superficie interna del talo. En sección longitudinal presenta tres capas de células a todo lo largo de la lámina, excepto en el ápice. Margen con 14.8 μm de grosor. Células corticales internas de 7.4 μm de largo por 7.4 μm de ancho, las externas 14.8 μm de largo por 7.4 μm de ancho. Zona media de 74 μm de grosor, en esta zona las células internas 48.1 a 55.5 μm de largo por 18.5 μm de ancho, células corticales externas 37 μm de largo y 29.6 μm de ancho, célula medular de 55.5 a 66.6 μm de largo por 18.5 a 22.2 μm de ancho. Zona basal de 85.1 μm de grosor, en este caso las células corticales internas de 85.1 μm de largo por 37 μm de ancho, células corticales externas de 44.4 a 74 μm de largo por 25.9 μm de ancho, célula medular de 77.7 a 81.4 μm de largo por 25.9 μm de ancho. Soros anteridiales en bandas con 2 mm de distancia sobre la superficie externa. Anteridios de 48.1 a 51.8 μm de largo por 18.5 μm de ancho.
Gametofito femenino figuras 4-5
Talo de 4.0 a 10 cm de largo, lámina lobado-dividida de 1.0 a 6 cm de ancho, color café amarillento, estípite de 3 a 5 mm de largo. Presencia de carbonato de calcio en la superficie interior. En sección longitudinal presenta tres capas de células a todo lo largo de la lámina, excepto en el ápice. En el margen de 18.5 a 60.2 μm de grosor. Células corticales internas de 22.2 a 29.6 μm de largo por 11.1 a 33.3 μm de ancho, las corticales externas de 25.9 μm de largo por 7.4 a 25.9 μm de ancho. En la zona media, con 77.7 μm de grosor. Células corticales internas de 37 μm de largo por 25.9 μm de ancho, las externas de 59.5 a 111 μm de largo por 18.5 a 29.6 μm de ancho, célula medular 62.9 a 74 μm de largo por 37 a 59.2 μm de ancho. Zona basal de 62.6 a 74 μm de grosor. Células corticales internas 48.1 a 70.3 μm de largo por 22.2 a 33 μm de ancho, las externas de 29.6 μm de largo por 22.2 a 25.9 μm de ancho, célula medular de 40.7 a 70.3 μm de largo por 18.5 a 22.2 μm de ancho. Pelos feofíceos presentes en ambas superficies de la lámina, se localizan junto a los soros. Soros oogoniales en bandas con 3 a 5 mm de distancia en la superficie externa de la lámina, oogonios de 66-74 μm longitud y 25.9-74 μm de ancho.
Talo monoico figuras 6-7
Talo de 10 cm de largo, lámina lobado-dividida de 3 a 4 cm de ancho, color café amarillento, estípite anchamente cónico de 1 cm de largo y 3 mm de ancho, estuposo. Presencia de carbonato de calcio en la superficie interior. En sección longitudinal presenta tres capas de células a todo lo largo de la lámina, excepto en el ápice. En el margen con 70.3 μm de grosor. Células corticales internas de 51.8 μm de largo por 33.3 a 37 μm de ancho, células corticales externas de 29.6 μm de largo por 14.8 a 29.6 μm de ancho. Zona media de 85 μm de grosor. Células corticales internas de 40.7 a 44.4 μm de largo por 40.7 a 50.2 μm de ancho, las externas de 29.6 μm de largo por 11.1 a 33.3 μm de ancho, célula medular 66.6 μm de largo por 22.2 a 25.9 μm de ancho. Zona basal, con 70.3 μm de grosor. Células corticales internas 55.5 a 62 μ m de largo por 37 μm de ancho, las externas 70.3 μm de largo y 22.2 a 29.6 μm de ancho, célula medular de 33.3-85.1 μm de largo por 25.9 μm de ancho. Pelos feofíceos presentes en ambas superficies, soro anteridial de 355 μm de largo, anteridios 37 μm largo y 29.6 μm de ancho. Oogonios dispersos de 66.6 μm longitud y 74 μm de ancho.
Talo de 3.5 a 15 cm de largo, lámina lobado-dividida de 3 a 7 cm de ancho, color café amarillento, estípite cónico estuposo de 0.5 a 1.7 cm de longitud. Presencia de carbonato de calcio en la superficie interna de ligera a abundante. En sección longitudinal presenta tres capas de células a todo lo largo de la lámina, excepto en el ápice. Margen con 37 a 77 μm de grosor. Células corticales internas de 25.9 a 44.4 μm de largo por 18.5 a 37 μm de ancho, las externas de 22.2 a 33.3 μm de largo por 18.5 a 25.9 μm de ancho. Zona media con 80 a 111 μm de grosor. Células corticales internas de 51.8 a 74 μm de largo por 25.9 a 33.3 μm de ancho, célula medular de 51.8 a 74 μm de largo por 22.2 a 29.6 de ancho. Zona basal de 85.1 a 149 μm de grosor, células corticales internas de 59.2 a 99.9 μm de largo por 25.9 a 37 μm de ancho, las corticales externas de 37 a 44.4 de largo por 33.3 a 37 μm de ancho, célula medular de 59.2 a 99.9 μm de largo por 33.3 a 37 μm de ancho. Pelos feofíceos presentes en ambas superficies de la lámina, se localizan junto a los soros. Soros esporangiales en bandas de 2 a 3 mm de distancia sobre la superficie externa de la lámina. Esporangios de 87.5 a 155.4 μm de longitud y 74 a 136.5 μm de ancho.
DISCUSÓN
El ciclo de vida en el orden Dictyotales es con alternancia de fases isomórfico, en donde se presentan con mayor frecuencia talos esporofitos y escasamente talos gametofitos (Ramón y Friedmann 1966). Esta característica también fue observada por Allender "y" Kraft (1983), en 22 especies de Dictyotales de Australia, en su mayoría fueron esporofitos a excepción de los gametofitos de: Zonaria diesingiana J. Agardh (masculino), Padina boryana (femenino), P. crassa Yamada (femenino) y Spatoglossum macrodontun J. Agardh (masculino). Gómez-Garreta (1983) encontró talos con oogonios en: Dictyota dichotoma (Hudson) Lamouroux y D. spiralis Montagne pero no se refirió a los talos masculinos. Abbott y Huisman (2004) sólo encontraron gametofitos fértiles en cuatro especies de Dictyota de un total de 23 taxa de Dictyotales, incluyendo siete especies de Padina. Gómez-Garreta et al. (2007), registraron gametofitos fértiles de Padina pavonica (L.) Thivy para la costa en la península ibérica, destacando una proporción mayor de esporofitos en contraste con los gametofitos. Esta condición fue explicada en el trabajo experimental de Gaillard (1972) quien sólo obtuvo generaciones de esporofitos en cultivo, lo que sugirió la probabilidad de un fenómeno de apomeiosis en la célula madre de las esporas, dando como resultado que al germinar las esporas sólo se formaban talos diploides y por lo tanto la meiosis era rara. También esta autora consideró que en el ambiente natural de P. pavonica y factores ecológicos pueden influir en la mayor proporción de esporofitos sobre los gametofitos.
Lo encontrado en el presente estudio con Padina boergesenii, confirma que la frecuencia de talos gametofitos es muy baja en comparación con la de esporofitos, tal y como sucede en otras especies del género (Ávila-Ortiz & Pedroche, 2005).
En la zona de estudio, los esporofitos se presentaron con mayor frecuencia en abril, mayo y junio (primavera y verano) (tabla 1), disminuyeron a partir de agosto y fueron escasos de diciembre a marzo (invierno). Este resultado es similar a lo encontrado por De Clerck & Coppejans (1996), quienes observaron que esta especie se desarrollaba durante la primavera y desaparecía a finales del verano en el Golfo de Arabia.
Thivy (1959), reconoció gametofitos masculinos y femeninos en enero, que se relaciona con el verano de Mauritius e India. En este trabajo, los gametofitos fueron reconocidos en ejemplares depositados en el herbario ENCB, obtenidos durante el otoño, el masculino recolectado en septiembre de 1993 y un femenino en noviembre de 1984, ambos en Quintana Roo. El talo monoico pertenece a un ejemplar de herbario en Isla Chica, Yucatán recolectado en julio de 1960, que corresponde al verano de México con soros anteridiales maduros y escasos oogonios aislados sin indusio (fig. 7); en el caso de P. pavonica en los talos monoicos, los soros anteridiales presentan pelos feofíceos y alternan con soros oogoniales que además tienen indusios (Gómez-Garreta et al., 2007).
La talla máxima de los talos de Padina boergesenii, se registraron durante los meses de mayo a julio, con 12 a 17.5 cm de alto y las menores en octubre a diciembre de 3.0 a 5.0 cm (tablas 2 y 3).
Este hecho coincide con las temperaturas cálidas superficiales del agua de mar durante primavera y verano (Mendoza-González y Mateo-Cid, 1992). Como se observa en las figuras 14 a 19 (15, 16, 17, 18), las láminas de los esporofitos muestran una notable variación morfológica, ya que se encontraron talos con láminas enteras (figs. 18, 19) de 3 a 8 cm de alto; y talos con láminas divididas que medían desde 5 hasta 17.5 cm de alto (figs. 14-17). Cabe mencionar que los talos de mayor talla fueron recolectados en localidades insulares como Isla Pájaros e Isla Mujeres (tabla 3) y los de menor talla se ubicaron en localidades litorales como en Playa Penacho del Indio y Playa Escondida (tabla 2). Taylor (1960) también hizo tal apreciación sobre el tamaño pequeño en la flora litoral en comparación con las de las islas. Por lo que respecta a los gametofitos, el talo masculino midió 4 cm (fig. 3) y los femeninos 4 y 10 cm de alto (fig. 4), el talo monoico tenia una talla de 10 cm de alto (fig. 6), estos hallazgos confirman que esta especie puede ser dioica o monoica. La variación en la talla y morfología de los talos esporofíticos y gametofíticos de Padina boergesenii es registrada por primera vez en el área de estudio.


Padina boergesenii es una especie de distribución tropical y se encuentra en Florida, Antillas Mayores, Antillas Menores, Cuba, Colombia, Venezuela, Panamá, Eritrea, Libia, Sudáfrica, Italia, Israel, Sri Lanka, Oman, Pakistán, Indonesia, Australia, Nueva Zelanda, Hawaii, Guam y Fiji (Littler y Littler, 2000; Guiry y Guiry, 2010). En México, P. boergesenii se ubica en la costa de Veracruz, Campeche, Yucatán y Quintana Roo (fig. 1), aunque se presenta con mayor frecuencia en Veracruz y Quintana Roo, quizá el sustrato sea un factor importante para su establecimiento de esta especie, ya que en Veracruz existen puntas rocosas de origen volcánico (Ortega et al., 2001), mientras que en Quintana Roo hay plataformas rocosas de origen calcáreo (Mendoza-González y Mateo-Cid, 1992). En el caso de Campeche y Yucatán, los sustratos en donde se obtuvieron los especímenes de P. boergesenii, fueron en esqueletos de coral, rocas de escolleras y otros sustratos artificiales y en su gran mayoría se encuentran grandes extensiones de playas areno-limosas, lo que probablemente sea el motivo de la escasa presencia de esta especie en ambos estados.
AGRADECIMIENTOS
Las autoras agradecen al Programa de Apoyo a Proyectos de Investigación e Innovación Tecnológico con la clave (PAPIIT-IN211505-3), por el apoyo en la compra de equipo y salidas a campo. Se agradece al Instituto Politécnico Nacional y a la Facultad de Estudios Superiores Zaragoza, por proporcionar las instalaciones y equipo necesario para el desarrollo de este estudio. La tercera autora agradece la beca otorgada por la Comisión de Operación y Fomento de Actividades Académicas (COFAA). A los alumnos S. Díaz-Martínez y U. García participantes y becarios en este proyecto. A Carlos A. Sánchez Mendoza por la ayuda en la elaboración de las láminas.
LITERATURA CITADA
Abbott, I.A. y J.M. Huisman, 2004. "Marine green and brown algae of the Hawaiian Islands". Bishop Museum Bulletin in Botany, 4: 1-259. [ Links ]
Allender, B.M. y G.T. Kraft, 1983. "The marine algae of Lord Howe Island (New South Wales); the Dictyotales and Cutleriales (Phaeophyta)". Brunonia, 6: 73-130. [ Links ]
Ávila-Ortiz, A., 2003. "Una variedad nueva de Padina mexicana (Dictyotaceae) para el Pacifico Tropical Mexicano". Hidrobiológica, 13: 69-74. [ Links ]
Ávila-Ortiz, A. y F.F. Pedroche, 2005. "El género Padina (Dictyotaceae, Phaeophyceae) en la región tropical del Pacífico mexicano". En: Monografías Ficológicas. Vol 2. Eds A. Sentíes G. and K. M. Dreckmann. Universidad Autónoma Metropolitana-Iztapalapa e Instituto de Botánica Sao Paulo, Brasil. p. 139-171. [ Links ]
Chapman, V.J., 1963. "The marine algae of Jamaica. Part 2. Phaeophyceae and Rhodophyceae". Bulletin Institute Jamaica Science Series, 12: 1-201. [ Links ]
Dawson, E.Y., C. Acleto y N. Foldvik, 1964. "The seaweeds of Peru". Beihefte zur Nova Hedwigia, 13: 1-111. [ Links ]
De Clerck, O. y E. Coppejans, 1996. "Marine algae of the Jubail Marine Wildlife Sanctuary, Saudi Arabia". En: (F. Krupp, A.H. Abuzinda & I.A. Nader, (eds.). A Marine Wildlife Sanctuary for the Arabian Gulf. Environmental Research and Conservation Following the 1991. Gulf War Oil Spill. NCWCD, Riyadh y Senckenberg Research Institute, Frankfurt a. M. p. 199-289. [ Links ]
Díaz-Martín, M.A., E. Torres-Mejía y J. Espinoza-Avalos, 1998. "Lista de algas del Área de Protección Yum Balam, Quintana Roo, México". Revista Biología Tropical, 46(3): 487-492. [ Links ]
Earle, S., 1969. "Phaeophyta of the eastern Gulf of Mexico". Phycologia, 7: 71254. [ Links ]
Fagerberg, W.R. y C. Dawes, 1973. "An electron microscope study of the spo-rophytic and gametophytic plants of Padina vickersiae Hoyt". Journal of Phycology, 9: 199-204. [ Links ]
Gaillard, J., 1967. "Étude monographique de Padina tetrastromatica (Hauck)". Bulletin de l'Institut fondamental d'Afrique Noire, 29: 447-463. [ Links ]
––––––––––, 1972. "Quelques remarques sur le cycle reproducteur des Dictyotales et sur ses variations". Societé Botanique de France, Mémoires, 1972: 145-150. [ Links ]
––––––––––, 1975. "Padina sanctae-crucis Boergesen, Padina japonica Yamada, Padina haitiensis Thivy et leurs affinities". Botaniste, 57: 85-103. [ Links ]
Gómez-Garreta, A., 1983. "Estudio fenológi-co de la vegetación marina de la isla de Mallorca". Tesis Doctoral. Universidad Complutense de Madrid. 269 p. [ Links ]
Gómez-Garreta A., J. Rull., M.C. Barcelò y M.A. Ribera, 2007. "On the presence of fertile gametophytes of Padina pavonica (Dictyotales, Phaeophyceae) from the Iberian coast". Anales del Jardín Botánico de Madrid, 64(1): 27-33. [ Links ]
Graham, L.E. y L.W. Wilcox, 2000. Algae. Prentice Hall, Nueva Jersey. 640 p. [ Links ]
Guiry, M.D y W.D. Guiry, 2010. AlgaeBase version 4.2. World-wide electronic publication, National University of Ireland, Galway. http://www.algaebase.org [ Links ]
Holmgren, P.K., N.H. Holmgren y L.C. Barnett, 1990. "Index herbarium Part I". The Herboria of the World. New York. 693 pp. (Regnum Vegetabile Vol. 20). [ Links ]
Huerta, M. L., 1961. "Flora marina de los alrededores de la Isla Pérez, Arrecife Alacranes, sonda de Campeche, México". 1961. Anales de la Escuela Nacional de Ciencias Biológicas, México, 10: 11-22, 1 fig., 1 cuadro. [ Links ]
Huerta, M.L. y M.A. Garza Barrientos, 1964. "Algas marinas de la Barra de Tuxpan y de los arrecifes Blanquilla y Lobos". Anales de la Escuela Nacional de Ciencias Biológicas, México, 13: 5-21, 1 fig., 1 cuadro. [ Links ]
Lawson, G.W. y D.M. John, 1987. "The marine algae and coastal environment of Tropical west Africa" (Second Edition). Nova Hedwigia, 93: 1-415. [ Links ]
Littler, D.S y Littler, M.M., 2000. Caribbean Reef Plants. Off Shore Graphics. Inc. China. 542 p. [ Links ]
Mendoza-González, A.C. y L.E. Mateo-Cid, 1992. "Algas marinas bentónicas de la isla Mujeres, Quintana Roo, México". Acta Botánica Mexicana, 19: 37-61. [ Links ]
Mendoza-González, A.C., L.E. Mateo-Cid y R.B. Searles. 2000. "New records of benthic marine algae from Isla Cozumel, México: Phaeophyta and Chlorophyta". Bulletin of Marine Science, 66(1): 119-130. [ Links ]
Ortega, M.M., J.L. Godínez y G. Garduño-Solórzano, 2001. Catálogo de algas Bénticas de las Costas mexicanas del Golfo de México y Mar Caribe. Comisión de Estudio para la Biodiversidad y Universidad Nacional Autónoma de México, México DF. 594 p. [ Links ]
Phillips, J.A., 1988. "Reproduction in Southern Australian species of the Dictyotales (Phaeophyta)". Botanica Marina, 31: 437-445. [ Links ]
Ramón, E. y I. Friedmann, 1966. "The ga-metophyte of Padina in the Mediterranean". Proceedings of the International Seaweeds Symposium, 5: 183-196. [ Links ]
Santelices, B., 1977. Ecología de algas marinas bentónicas. Universidad Católica de Chile, Santiago. 784 p. [ Links ]
Taylor, W.R.,1960. Marine Algae of Eastern Tropical and Subtropical Coasts of Americas. University of Michigan Press. Ann. Arbor. 870 pp. [ Links ]
Thivy, F., 1959. "On the morphology of the gametophytic generation of Padina gymnospora (Kützing) Vickers". Journal of the Marine Biological Association of India, 1: 69-66. [ Links ]
Trono, J.F., 1997. Field guide and atlas of the seaweeds resources of the Philippines. Bookmark. Makati City. [ Links ]
Womersley, H.B.S., 1987. The Marine Benthic Flora of Southern Australia. Part. II. Dep., of Bot. South Australian Government. Print. Div. Adelaide. 484 pp. [ Links ]
Wynne, M.J. y De Clerck, O., 1999. "First reports of Padina antillarum and P. glabra (Phaeophyta, Dictyotaceae) from Florida, with a key to the western Atlantic species of the genus". Caribbean Journal of Science, 35(3-4): 286-295. [ Links ]

