










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPolibotánica
versión impresa ISSN 1405-2768
Polibotánica no.29 México mar. 2010
Composición y estructura de la vegetación en tres sitios del estado de Nuevo León, México
Composition and structure of vegetation in three sites in the state of Nuevo Leon, Mexico
Humberto González-Rodríguez1, Roque G. Ramírez-Lozano2, Israel Cantú-Silva1, Marco V. Gómez-Meza3 y José I. Uvalle-Sauceda1
1 Facultad de Ciencias Forestales, Universidad Autónoma de Nuevo León, Apartado Postal 41, Linares, Nuevo León, 67700, México. Correo electrónico: humberto@fcf.uanl.mx
2 Facultad de Ciencias Biológicas, Universidad Autónoma de Nuevo León. San Nicolás de los Garza, Nuevo León, México.
3 Facultad de Economía, Universidad Autónoma de Nuevo León. Monterrey, Nuevo León, México.
Recibido: 28 mayo 2009.
Aceptado: 2 diciembre 2009.
Resumen
En verano de 2004, en tres sitios del matorral espinoso tamaulipeco, se determinó el estatus de la vegetación arbustiva y arbórea. Los sitios fueron localizados en los municipios de Los Ramones (S1), China (S2) y Linares (S3), Nuevo León, México. En cada sitio se establecieron, aleatoriamente, tres transectos (50 m x 7 m), en los cuales se estimaron los valores relativos de abundancia, dominancia, frecuencia y valor de importancia (VI) de los árboles y arbustos presentes. Además, se determinaron parámetros dasométricos como altura y diámetro de copa. La diversidad de especies para cada sitio se estimó con el índice de Shannon Wiener. La similitud entre sitios se determinó con el índice de Jackard. Se registraron un total de 1 741 individuos. Las plantas encontradas pertenecen a 20 familias, predominando los géneros y especies de la familia Fabaceae (10) seguidos por Euphorbiaceae (4) y Rhamnaceae (4), Rutaceae (3) y Cactaceae (2). En las restantes 15 familias sólo presentaron un solo tipo de individuo. El índice de Shannon mostró que no hay diferencias en la biodiversidad entre sitios; sin embargo, el índice de Jackard mostró igualdad entre especies sólo entre S1 y S2. Las especies de plantas más frecuentes fueron Acacia rigidula (255) seguida por Viguera stenoloba (171), Havardia pallens (167), Karwinskia humboldtiana (132), Forestiera angustifolia y Castela texana (125); representando el 56% del total encontrado y registran los valores más altos de los diferentes índices aplicados. Las especies menos frecuentes fueron: Condalia spahtulata (7), Ebenopsis ebano (7), Condalia hookeri (6), Wedelia acapulcensis (5), Cordia boissieri (4), Acacia farnesiana y Yucca treculeana con (3), Helietta parvifolia (2), Croton torreyanus (1) y Prosopis sp. (1); representando estas últimas sólo un 2.2%. En el S3 se registró una riqueza específica de individuos de poco más de 100% que en S1 o S2. Linares fue el sitio que presentó mayor diversidad de especies, posiblemente debido a la alta precipitación históricamente registrada.
Palabras clave: matorral espinoso tamaulipeco, Linares, China, Los Ramones, Nuevo León, composición y estructura de la vegetación.
Abstract
During the summer of 2004, a study of the status of woody vegetation was performed at three sites in the Tamaulipan thornscrub. The sites were located in the municipalities of Los Ramones (S1), China (S2) and Linares (S3) in the state of Nuevo Leon, Mexico. At each site three random transects (50 m X 7 m) were laid out, and relative abundance, relative dominance and relative frequency were estimated along each transect. In addition, dasometric parameters, such as height and crown diameter, were determined. Plant diversity was estimated by the Shannon-Wiener index, and similarity between sites was calculated using the Jackard index. A total of 1 741 individual plants belonging to 20 families were registered. Fabaceae had the highest number of species (10) followed by Euphorbiaceae (4) and Rhamnaceae (4), Rutaceae (3) and Cactaceae (2). The other 15 families were represented by only one species. The Shannon index showed that there were no statistical differences in biodiversity between sites; however, the Jackard index showed similarity among species only between S1 and S2. The most frequent species in the three sampling sites were Acacia rigidula (255) followed by Viguera stenoloba (171), Havardia pallens (167), Karwinskia humboldtiana (132), Forestiera angustifolia and Castela texana (125). This group represented about 56% of the total. Less frequent species were: Condalia spathulata (7), Ebenopsis ebano (7), Condalia hookeri (6), Wedelia acapulcensis (5), Cordia boissieri (4), Acacia farnesiana and Yucca treculeana with (3), Helietta parvifolia (2), Croton torreyanus (1) and Prosopis sp. (1), and they represented only about 2%. S3 registered the highest species richness with a little more than 100% of that of S1 or S2. That Linares was the site with the greatest species diversity might have been due to the higher precipitation recorded historically.
Key words: Tamaulipan thornscrub, Linares, China, Los Ramones, Nuevo León, composition and structure of vegetation.
INTRODUCCIÓN
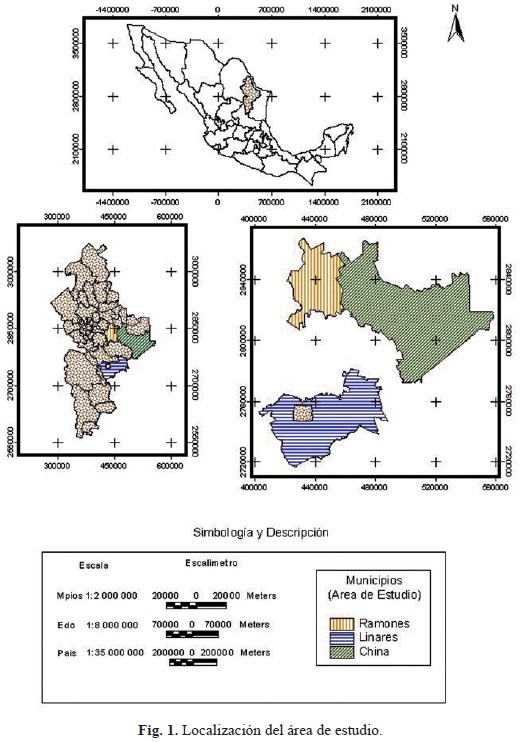
El ecosistema denominado matorral espinoso tamaulipeco (MET) cubre una superficie de 200 000 km2 (Fig. 1) del noreste de México (Coahuila, Nuevo León y Tamaulipas) y sur de Texas. Abarca de la Llera de Canales y los límites sureños de la Sierra Azul en Tamaulipas al altiplano Edwards en Texas y de las faldas de la Sierra Madre Oriental hasta la costa del Golfo de México (Alanís, 2006). Se clasifica en cuatro grandes asociaciones: 1) bajo espinoso e inerme, 2) mediano principalmente espinoso y subinerme, 3) alto subinerme y espinoso y 4) matorral crasirrosulifolio espinoso. Tiene una gran diversidad de plantas, dentro de las cuales las xerófitas son dominantes. Es un importante ecosistema que cumple funciones vitales por la protección que brinda a los recursos suelo y agua y su función como hábitat de la fauna silvestre (Alanís et al., 2008).
Las plantas presentes en el MET se distinguen por un amplio rango de grupos taxonómicos mostrando diferencias en patrones de crecimiento, diversidad en la longevidad foliar, dinámicas de crecimiento y desarrollo fenológico (McMurtry et al., 1996; Northup et al., 1996). La precipitación de la región varía de 400 a 800 mm y la evapotranspiración potencial anual es de aproximadamente 2 200 mm (González et al., 2004). En el caso particular del estado de Nuevo León, la distribución del clima y suelo no es uniforme. La distribución irregular de la precipitación y temperatura en el estado ha ocasionado que se puedan identificar regiones áridas y semiáridas (INEGI, 2002).
Sin embargo, la amplia diversidad de plantas nativas del estado de Nuevo León, es un ejemplo típico de una gran plasticidad en la respuesta a factores físico-químicos extremos (Reid et al., 1990). Especies arbustivas como chaparro prieto (Acacia rigidula), mezquite (Prosopis glandulosa), anacahuita (Cordia boissieri), tenaza (Havardia pallens), huizache (Acacia farnesiana), panalero (Forestiera angustifolia), granjeno (Celtis pallida), cenizo (Leucophyllum frutescens), guayacán (Guaiacum angustifolium), palo verde (Cercidium macrum), guajillo (Acacia berlandieri), nopal (Opuntia engelmanni), entre otras, generalmente forman parte de las especies dominantes de algunos tipos vegetativos propios de esta zona (INEGI, 2002; Estrada et al., 2004). En particular, muchos árboles y arbustos de especies leguminosas son utilizadas como alimento para el ganado dado que permanecen verdes y con un contenido relativamente alto de nutrientes a lo largo del año (Ramírez, 2009). Sin embargo, el uso de la vegetación como alimento por las grandes especies de herbívoros domésticos (ganado bovino) en las áreas naturales ha provocado modificaciones en la composición florística y la estructura de la formación vegetal (Turner, 1999); esta transformación de la estructura vegetal ha ocasionado cambios importantes en el balance de las poblaciones silvestres. El impacto ecológico que ha provocado la pérdida de biodiversidad se asocia también a problemas de deforestación y cambios en el uso del suelo. El efecto más importante para los manejadores y propietarios de ranchos diversificados es el desplazamiento de las poblaciones de fauna silvestre hacia lugares que puedan proporcionar los requerimientos básicos como alimento, agua, cobertura vegetal y espacio vital, para obtener un desarrollo vigoroso y saludable de los animales silvestres (Ramírez, 2004).
La identificación de especies vegetales del matorral, es relevante en el entendimiento de la estructura de la vegetación, la diversidad de especies, abundancia y la dinámica de las comunidades vegetales. La caracterización de la vegetación es importante debido a que la distribución de las especies no es similar y la aparición de una especie puede ser nula o abundante, por tener limitaciones de distribución por algún factor edáfico o climático. De esta forma, la información generada puede ser un medio para hacer propuestas de conservación o regeneración en áreas reducidas o fragmentos de vegetación del matorral (Rosenzweigh, 1995). Sin embargo, no se han llevado a cabo estudios recientes sobre diversidad en el matorral espinoso mediano, principalmente espinoso y subinerme, de los municipios del centro del estado de Nuevo León. Los objetivos de este estudio fueron determinar y comparar los índices diversidad y riqueza de las especies presentes del matorral espinoso mediano, en tres sitios de muestreo localizados en los municipios de China, Los Ramones y Linares.
MATERIAL Y MÉTODOS
El estudio fue realizado durante el verano de 2004 en tres sitios localizados en el estado de Nuevo León, México (Fig. 1). El primer sitio (S1) fue ubicado en el Rancho "El Abuelo" en el municipio de Los Ramones (25° 40' N; 99° 27' O), con una superficie de 100 ha y una elevación de 200 m.s.n.m. El clima es semiárido con verano cálido. La temperatura media anual es de 22°C. La precipitación media anual es de aproximadamente 700 mm. El segundo sitio (S2) fue localizado en el rancho "Zaragoza", en el municipio de China (25° 31' N; 99° 16' O), con una superficie de 300 ha y una elevación de 200 m.s.n.m. El clima es seco y cálido a través del año. La precipitación promedio total anual es de 500 mm, con una temperatura media anual de 22°C. El tercer sitio (S3) fue localizado en el Campus Experimental de la Facultad de Ciencias Forestales de la Universidad Autónoma de Nuevo León (24° 47' N; 99° 32' O), con una superficie de 500 ha y una elevación de 350 m.s.n.m., localizado en el municipio de Linares. El clima es subtropical y semiárido con verano cálido. La temperatura media mensual del aire varía de 15 a 22°C y la precipitación anual media es aproximadamente de 800 mm (Reid et al., 1990). La distancia entre S1 y S2 es de aproximadamente 54 km, de S2 a S3 es de 131 y de S1 a S3 es de 185 km. En los tres sitios, temperaturas de 40°C son comunes durante el verano. En general, se agrupan bajo un patrón climático similar con picos de precipitación máxima durante mayo, junio y septiembre. El principal tipo de vegetación se conoce como MET o también nombrado como matorral espinoso subtropical. Los suelos dominantes son vertisoles profundos, gris oscuro, limo-grisáceos, limo-arcillosos, con Montmorillonita, que se contraen y se expanden perceptiblemente en respuesta a los cambios en el contenido de humedad del suelo (INEGI, 2002).
En cada sitio de muestreo se establecieron al azar tres unidades de muestreo con una longitud de 50 m x 7 m y separadas a una distancia aproximada de 500 m. En cada unidad se cuantificó el número de ejemplares de cada especie vegetal (cuando la mitad o más del individuo se localizaban dentro de los límites de la unidad de muestreo se considero dentro; en caso contrario no se tomaba en cuenta). Además, se determinaron los parámetros dasométricos de altura (m) y diámetro (m) de copa (este último tomando en cuenta la medición del largo por el ancho de la especie y tomada de norte a sur y de oriente a poniente).
La diversidad, en su definición, considera tanto al número de especies como también al número de individuos (abundancia) de cada especie existente en un determinado lugar. En este estudio se utilizó el índice de Shannon, uno de los más frecuentes para determinar la diversidad de plantas de un determinado hábitat (Matteucci et al., 1999). Para aplicar este índice, el muestreo debe ser aleatorio y todas las especies de una comunidad vegetal deben estar presentes en la muestra. Para determinar la similitud entre sitios de muestreo se utilizó el índice de Jackard, cuyos datos cualitativos están basados en la presencia o ausencia de las especies en los sitios (Magurran, 1988). El valor de importancia (VI) en cada sitio se estimó por medio de la suma de tres indicadores ecológicos: dominancia, densidad y frecuencia. En este estudio, para estimar la densidad de las especies se utilizaron los datos de cobertura en lugar del área basal. Heiseke y Foroughbakhch (1985) recomiendan lo anterior cuando la mayoría de las especies presentes son arbustos con una gran cantidad de tallos con diámetros menores a 1 cm.
Los datos obtenidos con el índice de Shannon fueron analizados estadísticamente. Los resultados no presentaron homogeneidad de varianzas al igual que tampoco mostraron una distribución normal de acuerdo a las pruebas estadísticas de Kolmogorov-Smirnov (con la corrección de Lilliefors) y Shapiro-Wilk (Brown y Forsythe, 1974); por tanto, los datos fueron analizados mediante la prueba no paramétrica de Kruskal-Wallis (Stell y Torrie, 1980; Ott, 1993) para detectar diferencias estadísticas en el índice de Shannon entre los sitios. Todos los procedimientos estadísticos fueron realizados mediante el uso del paquete estadístico SPSS versión 9.0 (SPSS, Inc., Chicago, IL, USA).
RESULTADOS Y DISCUSIÓN
Se registraron 1 741 individuos de las diferentes especies de árboles y arbustos en los nueve transectos establecidos. La altura promedio de las plantas fue de 1.18, 1.62 y 2.52 m para S1, S2 y S3, respectivamente (cuadro 1). Las especies de plantas encontradas pertenecen a 20 familias, predominando las especies de la familia Fabaceae (10), Euphorbiaceae (4) y Rhamnaceae (4), Rutaceae (3) y Cactaceae (2); en las restantes 15 familias sólo presentaron una especie. La abundancia de las plantas de la familia Fabaceae es probable que se deba a que en el estado de Nuevo León algunos taxa son abundantes sobre grandes extensiones, como Acacia rigidula, A. constricta, A. berlandieri y Prosopis glandulosa, asociados a diversas comunidades de matorral xerófilo (Estrada y Jurado, 2005; González et al., 1997) al describir la fisonomía y la composición florística de la vegetación secundaria de cinco áreas con clima, suelo y topografía similares, pero con historiales de uso de suelo diferentes, Espinoza y Návar (2005) al evaluar la estructura de la vegetación en 15 cuadrantes, establecidos en un gradiente de productividad en cinco sitios y Alanís et al. (2008) al analizar la diversidad de especies en áreas con distinto historial de actividades antropogénicas, también reportaron una mayor abundancia de especies de la familia Fabaceae en el MET del municipio de Linares, N.L., México. Estos autores concluyeron que la presencia de mayor número de este tipo de especies pudiera explicarse, parcialmente, por sus bajos y diferentes requerimientos nutricionales, la diferenciación en la explotación del recurso espacio-tiempo y a los diferentes patrones fisiológicos, entre otros.
El S2 tuvo menor riqueza específica (335 especies) que el S1 (487) y el S3 (919). Sin embargo, la prueba de Kruskal-Wallis demostró que los datos obtenidos de diversidad al aplicar el índice de Shannon, los sitios no fueron significativamente diferentes entre sí con valores de 2.6, 2.8 y 2.4 para S1, S2 y S3, respectivamente. Resultados similares (2.1, 2.7, 2.5 y 2.5) fueron reportados por Domínguez (2009) quien cuantificó la diversidad de especies en cuatro sitos localizados en el MET caracterizados por diferencias en altitud y Espinoza y Návar (2005) en cinco sitios con diferente nivel de productividad en el MET, con 2.8 2.5, 2.7, 3.2 y 2.9, y argumentan que la falta de significancia estadística en el índices de Shannon pudiera deberse a que los cambios en la riqueza de especies se equilibran con los cambios en la abundancia. Es decir, mientras que el gradiente de riqueza de especies se mueve en una dirección, se observa que la abundancia se mueve en la dirección contraria. Alanís et al. (2008) reportaron valores menores (1.4, 2.1, 2.2 y 2.3, respectivamente) a los de este estudio, quizás por tratarse de que las áreas evaluadas corresponden a cuatro sitios (ganadería intensiva, ganadería extensiva, agricultura tradicional y matarasa) localizadas en el MET del municipio de Linares, con distinto historial de actividades antropogénicas con 21 años de abandono.
En general, la diversidad en los sitios de muestreo es caracterizada como intermedia en comparación con determinaciones similares obtenidas en análisis tipo para la región (Medina, 1995). Lo anterior se traduce en que el ecosistema de esa región en particular cuenta con una relevancia ecológica media. En este contexto, Segura et al. (2003), Prasad et al. (2007) y Alanís et al. (2008) señalan que la razón de la variación en el patrón de la diversidad de las especies es debida, en parte, a las condiciones microclimáticas del sitio, al ambiente físico y condiciones ecológicas del hábitat y en cierta medida a los cambios en el uso de suelo (ganadería extensiva).
El índice de Jackard muestra que S1 y S2 tienen mayor similitud de especies (75.8) que S1 y S3 (38.9) y S2 y S3 (36.1). Lo que significa que el S1 y S2 son muy similares en su composición de especies y el S3 destaca por su mayor número de individuos que lo hace desigual a los otros sitos quizás porque la precipitación registrada es mucho más abundante y por tratarse de un sitio sin disturbio antropogénico.
En el S1, las especies que mostraron mayor altura y cobertura fueron Cercidium macrum y Phaulothamnus spinescens y la que presentó los valores más bajos fue Yucca treculenta. Las de mayor abundancia fueron Acacia rigidula seguida por Karwinskia humboldtiana y Croton cortesianus; mientras que las que presentaron los valores menores fueron Lantana macropoda, Prosopis sp. y Yucca treculeana. Las especies con mayor dominancia fueron Phaulothamnus spinescens, Cercidium macrum y Bumelia celastrina mientras que las menos dominantes fueron Y. treculeana y Leucophyllum frutescens. Las plantas con mayor valor de importancia fueron Acacia rigidula seguida por Cercidium macrum y Karwinskia humboldtiana y las de menor fueron Yucca treculeana, Bernardia myricaefolia, Lantana macropoda, Ziziphus obtusifolia y Prosopis sp. (cuadro 1). En el S2, Yucca treculeana y Acacia rigidula fueron las de mayor altura y Schaefferia cuneifolia la de menor. Las plantas de mayor cobertura fueron Prosopis leavigata, Ebaopsis ebano y Acacia rigidula y Schaefferia cuneifolia la menor. Cercidium macrum fue la de mayor abundancia y Schaefferia cuneifolia fue la menor. Prosopis laevigata, Ebanopsis ebano y Yucca treculeana fueron más dominantes y Schaefferia cuneifolia, Randia rhagocarpa y Jatropha dioica la menos dominantes. Entre las especies con mayor frecuencia se encuentran Castela texana, Acacia rigidula y Bumelia celastrina. Las especies con mayor valor de importancia fueron Cercidium macrum y Acacia rigidula; sin embargo, Schaefferia cuneifolia, fue la de menor VI (cuadro 2). En el S3, Prosopis laevigata fue la de mayor altura y Randia rhagocarpa la de menor. La de mayor cobertura en el sitio fue Bumelia celastrina y la de menor fue Karwinskia humboldtiana. Las especies con los valores de abundancia más elevados fueron Viguiera stenoloba, Havardia pallens y Acacia rigidula, mientras que Prosopis laevigata fue menos abundante. Las especies más dominantes en el sitio fueron Bumelia celastrina, Helietta parvifolia y Acacia berlandieri. Por otra parte, Randia rhagocarpa, Karwinskia humboldtiana y Castela texana fueron las menos dominantes. Las especies con mayor frecuencia fueron Cordia boissieri, Acacia rigidula y Dyospiros texana y las menos frecuentes al momento del muestreo fueron Prosopis laevigata, Schaefferia cuneifolia, Leucophyllum frutescens, entre otras. Las especies con mayor valor de importancia fueron Havardia pallens, Viguiera stenoloba y Acacia rigidula, mientras que Schaefferia cuneifolia, Prosopis laevigata y Leucophyllum frutescens fueron menores (cuadro 3). Reid et al. (1990), al realizar un estudio sobre la variación florística y estructural en el MET, argumentaron que factores climáticos, topográficos y edáficos son los que originan los diferentes patrones de distribución de las especies. En este estudio es probable que estos mismos factores hayan contribuido a las variaciones en la estructura y composición de la vegetación en los tres sitios.
La arbustiva Acacia rigidula de la familia Fabaceae, en los tres sitios tuvo un alto VI. Asimismo, en otros estudios (Estrada et al., 2004) conducidos en el MET, la han identificado como una planta prevalente en la mayoría de los ecosistemas. Además, Alanís et al. (2008) reportaron que A. rigidula tuvo un alto valor de importancia en ecosistemas con historial de agricultura, ganadería y matarasa localizados en el municipio de Linares, N.L., México. González (1996) también reportó que junto con Acacia farnesiana y Acacia berlandieri, Acacia rigidula aparece como pionera después de un aprovechamiento. Por tanto, es muy probable que debido a su gran VI en la región, Acacia rigidula se identifique como un componente muy importante de la dieta de pequeños rumiantes en pastoreo como caprinos, ovinos y venado cola blanca (Ramírez, 2009).
CONCLUSIONES
En el sitio Los Ramones, las especies de plantas que presentaron mayor VI fueron Acacia rigidula, Karwinskia humboldtiana y Croton cortesianus; en el sitio China, fueron Forestiera angustifolia, Celtis pallida, Castela texana, Karwinskia humboldtiana, Croton cortesianus y en Linares, fueron Acacia rigidula, Castela texana, Lantana macropoda, Forestiera angustifolia, Leucophyllum frutescens, Zanthoxylum fagara y Karwinskia humboldtiana. Particularmente, en Linares se registro una mayor densidad específica de plantas lo que pudiera indicar que este sitio no está perturbado y que por su mayor precipitación, históricamente registrada, la diversidad de especies ha alcanzado un estadio sucesional avanzado. Sin embargo, la biodiversidad entre sitios no fue diferente por lo que se puede concluir que la vegetación tiende a ser homogénea.
LITERATURA CITADA
Alanís, R.E., 2006. Diversidad de especies arbóreas y arbustivas en áreas con distinto historial antropogénico en el matorral espinoso tamaulipeco, tesis de maestría. Facultad de Ciencias Forestales, Universidad Autónoma de Nuevo León. 66-67 pp. [ Links ]
Alanís, R.E.; Jiménez, P.J.; Aguirre, C.O.; Treviño, G.E.; Jurado, Y.E. y González, T.M., 2008. "Efecto del uso del suelo en la fitodiversidad del matorral espinoso tamaulipeco". Ciencia UANL, 11: 56-62. [ Links ]
Domínguez, T.G., 2009. Deposición de hojarasca y retorno potencial de nutrimentos en diferentes comunidades de vegetación, tesis de maestría, Facultad de Ciencias Forestales, Universidad Autónoma de Nuevo León, Linares, N.L. México. 49-71 pp. [ Links ]
Brown, M.B. y Forsythe, A.B., 1974. "Robust tests for the equality of variances". Journal of the American Statistical Association, 69: 364-367. [ Links ]
Espinoza, R. y Návar, J., 2005. "Producción de biomasa, diversidad y ecología de especies en un gradiente de productividad en el matorral espinoso tamaulipeco del Nordeste de México". Revista Chapingo, 11: 25-35. [ Links ]
Estrada, E.; Yen, C.; Delgado, A. y Villarreal, J.A., 2004. "Leguminosas del centro del estado de Nuevo León, México". Anales del Instituto de Biología, Universidad Nacional Autónoma de México, Serie Botánica. 75: 73-85. [ Links ]
Estrada, E. y Jurado E., 2005. ''Leguminosas del norte del estado de Nuevo León, México". Acta botánica Mexicana, 73: 1-18. [ Links ]
González, M. 1996. Análisis de la vegetación secundaria de Linares, N.L. México, tesis de maestría, Facultad de Ciencias Forestales, Universidad Autónoma de Nuevo León, Linares, N.L. México. 103 pp. [ Links ]
González, E. M.; Treviño, E.J. y Aguirre, O.A., 1997. "Fisonomía y composición de la vegetación secundaria en un área de matorral del noreste de México", Phytología, 83: 257-269. [ Links ]
González, R.H.; Cantú, S.I.; Gómez, M.M.V. y Ramírez, L.R.G., 2004. "Plant water relations of thornscrub shrub species, northeastern Mexico". Journal of Arid Environments, 58: 483-503. [ Links ]
Heiseke, D. y Foroughbakhch R., 1985. "El matorral como recurso forestal". Reporte Científico núm. 1, Facultad de Silvicultura y Manejo de Recursos Renovables, Linares, N.L., México. 1-31 pp. [ Links ]
INEGI., 2002. "Uso actual del suelo en los núcleos agrarios. Aspectos geográficos de Nuevo León". Consultado el 09 de mayo de 2009. Página Web del Instituto Nacional de Estadística, Geografía e Informática: http://nl.inegi.gob.mx/territorio/español/cartcat/uso.html. [ Links ]
Magurran, A., 1988. Ecological diversity and its measurements. Princeton University Press. New Jersey. 179 pp. [ Links ]
Matteucci, S.D.; Colma, A. y Pla, L., 1999. "Biodiversidad vegetal en el árido falconiano (Venezuela)". Interciencia, 24: 300-307. [ Links ]
McMurtry, C.R.; Barnes, P.W.; Nelson, J.A. y Archer, S.R., 1996. Physiological responses of woody vegetation to irrigation in a Texas subtropical savanna, La Copita Research Area. Consolidated Progress Report. Texas Agricultural Experiment Station-Corpus Christi, Texas A & M University System, College Station, Texas, USA. 33-37 pp. [ Links ]
Medina, M. del C., 1995. Fitodiversidad en relación al tamaño de fragmentos remanentes de matorral, en Linares N.L., México, tesis profesional. Facultad de Ciencias Forestales. Universidad Autónoma de Nuevo León, Linares, N.L. México. 44 pp. [ Links ]
Northup, B.K.; Zitzer, S.F.; Archer, S.R. y Boutton, T.W., 1996. A technique to allocate biomass of woody plants, La Copita Research Area. Consolidated progress report. Texas Agricultural Experiment Station - Corpus Christi, Texas A & M University System, College Station, TX. 47-50 pp. [ Links ]
Ott, L., 1993. An introduction to statistical methods and data analysis. 775 p. 2nd ed., Duxbury Press. Boston, Massachusetts. [ Links ]
Prasad, P.R.C.; Reddy, C.S. y Dutt, C.B.S., 2007. "Phytodiversity assessment of tropical rainforest of North Andaman Islands, India". Research Journal of Forestry, 1: 27-39. [ Links ]
Ramírez, R.G., 2004. Nutrición del Venado Cola Blanca. Ed. Publicaciones Universidad Autónoma de Nuevo León. México. 73-87 pp. [ Links ]
Ramírez, R.G., 2009. Nutrición de Rumiantes: Sistemas Extensivos. 2a Ed. Editorial Trillas. México. 216-224 pp. [ Links ]
Reid, N.; Marroquín, J.; Beyer-Münzel, P., 1990. "Utilization of shrubs and trees for browse, fuelwood and timber in the Tamaulipan thornscrub, northeastern Mexico". Forest Ecology and Management, 36: 61-79. [ Links ]
Rosenzweigh, M.L., 1995. Species Diversity in Space and Time. University of Cambridge Press. Great Britain at University Press. Cambridge. 50-72 pp. [ Links ]
Segura, G.; Balvanera, P.; Durán, E. y Pérez, A., 2003. "Tree community structure and stem mortality along a water availability gradient in a Mexican tropical dry forest". Plant Ecology, 169: 259-271. [ Links ]
Steel, R.G. y Torrie, J.H., 1980. Principles and procedures of statistics. 2nd ed. McGraw-Hill Book Co., New York, NY. 107-133 pp. [ Links ]
Turner, M.D., 1999. "Spatial and temporal scaling of grazing impacto n the species composition and productivity of Sahelian annual grasslands". Journal of Arid Environments, 41: 277-297. [ Links ]

