










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPolibotánica
versión impresa ISSN 1405-2768
Polibotánica no.27 México abr. 2009
Cambios en la estructura y composición de la vegetación relacionados con los años de abandono de tierras agropecuarias en la selva baja caducifolia espinosa de la reserva de Dzilam, Yucatán
Changes in the struture and composition of the vegetation relative to the years of abandonment of agricultural land in the tropical dry forest of the Dzilam reservation, Yucatan, Mexico
Jorge L. Leirana–Alcocer, Silvia Hernández–Betancourt, Luis Salinas–Peba y Leonardo Guerrero–González
Campus de Ciencias Biológicas y Agropecuarias, Universidad Autónoma de Yucatán Carretera Mérida–Xmatkuil Km. 15.5, AP 4–116 Itzimná CP 97100. Correo electrónico: ambienteprimero@yahoo.com.mx
Recibido: 20 octubre 2007
Aceptado: 21 enero 2009
Resumen
Las selvas bajas del norte de Yucatán son de gran importancia científica y conservacionista; sus condiciones de aridez, pedregosidad del suelo y su relativo aislamiento de otras regiones biogeográficas le permiten albergar una importante proporción de los endemismos de la península. No obstante lo anterior, esta área sufre de presiones por actividades humanas, principalmente la ganadería, agricultura y la extracción de piedra caliza. Por esta razón se hace necesario estudiar los cambios en la vegetación en las tierras agropecuarias después de su abandono. Se comparó la estructura y composición de árboles y arbustos de potreros derivados de selva baja caducifolia espinosa en Dzilam, Yucatán con diferentes tiempos de abandono. Se reportan los cambios en la composición y abundancia de las tres subfamilias pertenecientes a la familia Leguminosae (Papilonoideae, Mimosoideae y Caesalpinoideae) ya que es una familia muy importante en los trópicos secos y fue la más abundante y rica en nuestra área de estudio. Se encontró que la altura y la abundancia de individuos del estrato arbóreo y la riqueza del estrato arbustivo fueron las variables que cambiaron de manera más intensa con la edad de abandono (ANOVA, P < 0.05). Los sitios de pasto en uso y sitios con menos de cinco años de abandono mostraron una mayor semejanza en la composición de especies entre ellos que con los más antiguos. Las parcelas con más de 10 y 5 años de abandono formaron otro grupo, aunque con un bajo valor de similitud (Sorensen < 0.2). La familia Leguminosae fue la más rica en especies en los estratos arbustivo y arbóreo. Las leguminosas también se encontraron entre las más abundantes. Esto se explica por un suelo pobre en nutrientes y la historia de manejo intenso, ya que su capacidad de establecer simbiosis con microorganismos fijadores de nitrógeno les confiere una ventaja competitiva. Las abundancias relativas de las mimosoideas fueron las más altas en los sitios de reciente abandono, en tanto que las de papilonoideas dominaron en los sitios más antiguos. Las especies más abundantes en el sitio de más de 10 años resultaron ser Bursera simaruba, Metopium brownei y Plumeria rubra. Las primeras dos se encuentran en casi todos los tipos de vegetación terrestre de la Península de Yucatán. Las tres especies son resistentes a disturbios, como incendios o destrucción asociada a la explotación maderera y poseen una alta capacidad de regeneración, lo que puede indicar una historia de uso intenso de este tipo de vegetación.
Palabras clave: selva baja caducifolia espinosa, Yucatán, tierras postagrícolas, estructura de la vegetación, Leguminosae.
Abstract
The dry forests in the northern Yucatan Peninsula are highly important not only for science but also for the conservation of biodiversity because of their unique biogeographical and ecological characteristics. The combination of arid climate, stony soil and relative isolation from other biogeographical regions, has resulted in their sheltering an important proportion of the peninsula's endemic taxa. Nevertheless, this region suffers from such human pressures as cattle ranches and the exploitation of limestone banks. This scenario makes it necessary to describe the ecological changes on the land once it has been abandoned after being used for agriculture, because there is no pristine forest in Yucatan. We compared the structure and composition of trees and shrubs on abandoned agricultural land of different ages. All sites were derived from tropical dry forest, a vegetation type which contains several endangered species. The changes in composition and relative abundance of the subfamilies belonging to the Leguminosae (Papilonoideae, Mimosoideae and Caesalpinoideae) were also reported, since this family was the most abundant at the study site. Average tree height and density and shrub richness showed larger changes with increasing number of years without management (ANOVA, P< 0.05). Younger plots (1–5 years and currently in use) were more similar among themselves than to older sites (more than five and more than ten years). The Leguminosae were the richest family in species in all the plots and one of the most abundant, possibly indicating a history of intense use. Agriculture tends to deplete nitrogen from the soil, which gives a competitive advantage to legumes. Papilonoideae were the most abundant in older sites (more than five and more than ten years), whereas Mimosoideae dominated recently abandoned sites (five years or less) and those currently in use. Metopium brownei, Bursera simaruba and Plumeria rubra were the most abundant species in the oldest site. The first two are found in almost all types of terrestrial vegetation on the Yucatan Peninsula, and all three are reported as being very resistant to disturbance, such as fire or damage associated with wood harvest, and to regenerate readily. Their presence may indicate a history of intense use of this region.
Key words: tropical dry forest, Yucatan, abandoned agricultural land, vegetation structure, Leguminosae.
INTRODUCCIÓN
Las selvas bajas caducifolias del norte del estado de Yucatán son de gran importancia ecológica y conservacionista; su relativo aislamiento de otras regiones biogeográficas, aunado a la aridez del clima (Orellana et al., 2003) y la pobreza en nutrientes de sus suelos (Bautista–Zúñiga, et al., 2003), han dado como resultado una flora y fauna particulares (Carnevali et al., 2003, Flores–Guido y Espejel, 1994) que contienen un alto porcentaje de los endemismos de plantas y animales del estado (Ibarra–Manríquez et al., 2002).
No obstante lo anterior, esta zona sufre de presiones por parte de actividades humanas tales como la ganadería, la agricultura y la explotación de piedra caliza, incluso dentro de las áreas destinadas a la protección. Esto hace que para su mejor gestión se necesiten conocer mejor los cambios ecológicos asociados al uso del suelo, su posterior abandono y la recuperación de la cobertura vegetal (Lamb et al., 2005).
La cobertura vegetal está asociada íntimamente al funcionamiento del ecosistema (Harris y Hobbs, 2001, Parkes et al., 2003, 2004) y por tanto puede servir de indicador de lo que ocurre con el resto de la biota; por eso es importante documentar la velocidad y dirección de los cambios en diversas variables de la vegetación para tener mejores bases en el manejo y conservación de la biodiversidad.
En este trabajo se describen los cambios en estructura y composición de los pastizales artificiales derivados de selva baja caducifolia espinosa con base en los años de abandono. Para ello se estimará la cobertura de las especies de los estratos arbóreo y arbustivo en parcelas con diferentes edades de abandono y también se estimará la relación entre el tiempo de abandono y variables estructurales tales como la abundancia de árboles y arbustos, la altura promedio de los árboles, la riqueza de especies y familias y la cobertura de árboles y arbustos. También se analizarán los cambios en la composición de la familia Leguminosae y las tres subfamilias que la componen, ya que ésta fue la familia más rica y abundante del área de estudio.
ÁREA DE ESTUDIO
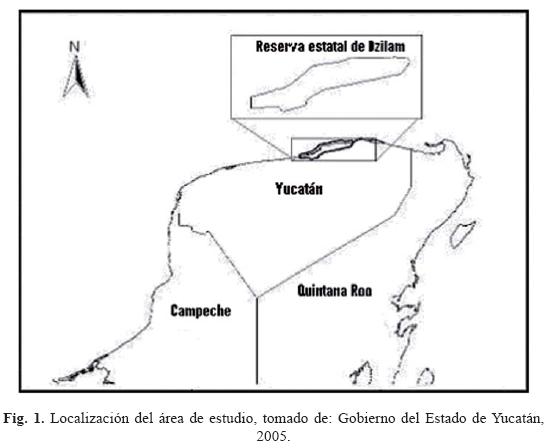
La reserva estatal de Dzilam se encuentra entre los municipios de Dzilam de Bravo y San Felipe, Yucatán, en la costa central del estado a 107 km al noreste de la ciudad de Mérida (UADY, 1999).
La superficie de la reserva es de 61 706.83 ha. y se localiza entre los meridianos 88° 47' 54.4'' y 88° 47' 49.7'' de longitud Oeste y entre los paralelos de 21° 26' 33.4'' y 21° 24' 51.2 " de latitud Norte (Gobierno del Estado de Yucatán, 2006).
El clima en el área es semiseco, muy cálido; en la clasificación de Koppen–García es BS1(h') w (x') (Orellana et al., 2003). La temperatura promedio anual de la zona va de los 25 a los 26°C y la precipitación promedio anual va de los 700 mm anuales a los 650 mm. Más del 65% del volumen de precipitación cae entre junio y noviembre (CNA, 2000).
El tipo de vegetación presente en el área de trabajo corresponde a una selva baja caducifolia espinosa, cuya característica más distintiva es la alta frecuencia de las familias Cactaceae, Leguminosae y Rubiaceae. Este tipo de vegetación está representada en la Península como una estrecha franja costera que corre desde la parte norte del estado de Quintana Roo, pasando por Yucatán hasta la porción norte del estado de Campeche (Flores–Guido y Espejel, 1994).
MÉTODOS
Se eligieron sitios con diferentes años de abandono en su uso según encuestas realizadas a los habitantes del lugar. A cada rango de años de abandono se le asignó una categoría que inició en I, para un pastizal en uso, II para un sitio con uno a cinco años de abandono, III para un sitio entre 5 y 10 años de abandono y IV, para los sitios con más de 10 años de abandono.
La distancia promedio entre los sitios de categoría I, II y III fue de 1.7 km. La distancia promedio entre el sitio IV y los demás fue de 37 km.
Para representar la heterogeneidad en la distribución de especies, en cada sitio se establecieron tres cuadrantes de 10 x 10 m con una distancia mínima entre cuadrantes de 100 metros. En el caso de la parcela de 5 a 10 años de abandono (categoría III), se observó una muy alta riqueza por cuadrante y una muy alta tasa de recambio de especies entre cuadrantes, por lo que se muestreó un cuadrante más.
Se censaron todos los individuos cuyas raíces y órganos aéreos estuvieran dentro de los cuadrantes. En los análisis se separan los individuos pertenecientes a los estratos arbóreo y arbustivo.
En el estrato arbóreo se incluyeron los individuos mayores a 2 m de alto (la altura se midió con una garrocha telescópica de dimensiones conocidas), con un diámetro a la altura del pecho (DAP) mayor a 2 cm, y cuyas ramificaciones comenzaran por lo menos a 1 m del suelo.
Se consideraron dentro del estrato arbustivo a los individuos con DAP's menores a 2 cm y una altura mayor de 50 cm. Cada individuo fue asignado solamente a un estrato, aunque hubieron especies que ocurrieron en ambos estratos.
Para cada cuadrante se registraron las siguientes variables denominadas "estructurales" que son importantes en la evaluación de ecosistemas terrestres (Harris y Hobbs, 2001, Parkes et al., 2003): 1. Densidad de individuos arbóreos., 2. Altura promedio de los individuos en el estrato arbóreo, 3. Cobertura total del estrato arbóreo, 4. Número de especies/cuadrante en el estrato arbóreo, 5. Densidad de arbustos, 6. Cobertura total del estrato arbustivo y 7. Número de especies/cuadrante en el estrato arbustivo.
Para ver si había diferencias de cada variable entre tratamientos, se realizaron ANO VAs y pruebas de rangos múltiples con el paquete Statgraphics Plus v 5.1. Se construyeron los gráficos de medias de cada variable.
En todos los casos el número de muestras (cuadrantes) totales fue de 13, por lo que los grados de libertad totales fueron de 12, de los cuales tres grados fueron entre grupos y nueve intragrupos.
Para determinar la importancia de las variables estructurales en la clasificación de los sitios en las categorías de edad de abandono, se llevaron a cabo análisis discriminantes (Martínez–Arias, 1999) con el paquete Statgraphics Plus v 5.1.
El análisis discriminante reclasifica a cada elemento, en este caso cuadrante, según sea más similar a los valores de los grupos originales. Si los elementos son reasignados a su grupo original en un mayor porcentaje que lo esperado al azar, las variables usadas son consideradas buenas discriminantes.
Se reporta el porcentaje de aciertos al asignar cada cuadrante a un grupo de edad; primero usando cada variable estructural una por una, y luego la combinación de variables que mejor diagnostica el rango de edad de los cuadrantes. También se reporta la significancia global de la clasificación evaluada con la prueba Q de Press, que toma en cuenta el número de aciertos, el tamaño de muestra y el número de grupos.
La diversidad taxonómica de árboles y arbustos de cada categoría de edad fue descrita usando los siguientes indicadores: Número de especies por cuadrante, índice Simpson de diversidad, número de familias y riqueza y abundancia relativa de la familia más abundante (Magurran, 2004, Peetchey y Gaston, 2006).
Para estimar la similitud en la composición de especies y familias entre sitios de diferente edad, se estimaron índices de Sörensen con el paquete MVSP 3.10b (Kovach Computing Services, 1999), estos índices fueron usados para construir dendrogramas de los sitios muestreados.
La familia Leguminosae se analizó por separado porque fue la más rica y abundante en el área de estudio. De esta familia se describió la proporción de especies de las subfamilias Caesalpinoideae, Mimosoideae y Papilonoideae dentro de cada categoría de edad.
RESULTADOS
En seis de las siete variables estructurales, los cuadrantes pertenecientes a diferentes grupos de edad presentaron diferencias significativas (P < 0.05). Solamente en la cobertura del estrato arbustivo no se encontraron diferencias significativas entre los grupos de edad de abandono.
En la figura 2 se presentan los gráficos de medias para todas las variables.
El porcentaje de aciertos del análisis discriminante, usando una variable estructural a la vez, se presenta en el cuadro 1. Se observa que la mejor clasificadora fue la altura promedio del estrato arbóreo, con un 100% de aciertos.
La combinación de variables que mejor explica la clasificación de los cuadrantes, es la pareja formada por la cobertura del estrato arbóreo con la riqueza específica del estrato arbustivo; con esta combinación se acertó el 100% de las veces.
En el cuadro 2, se presentan los valores de las funciones discriminantes que se obtuvieron usando estas variables.
Con respecto a la riqueza y diversidad de especies (cuadro 3) en el estrato arbóreo, las riquezas promedio por cuadrante no fueron significativamente diferentes entre los sitios, contrastes de bootstraps (P < 0.05). Sin embargo en el estrato arbustivo el sitio de edad III presentó un índice de Simpson significativamente más alto.
Es notoria la diferencia entre la riqueza total de ambos estratos y las de cada sitio muestreado; esto sugiere una alta tasa de recambio de especies entre sitios.
La diversidad de familias se resume en el cuadro 4. La dominancia más alta se dio en el estrato arbóreo de la edad III, donde la abundancia relativa de la familia más dominante fue de 48%, seguida de la categoría II de este mismo estrato con un 46% de individuos en la familia más dominante. En ambos casos fue la familia Leguminosae la más abundante.
En cuanto a la similitud en la composición de especies, los sitios I y II resultaron ser los más parecidos entre sí, aunque con un índice bajo (Sorensen < 0.3). Los sitios III y IV formaron el otro grupo, pero la similitud entre ellos resultó aún menor (Sorensen < 0.2) (figura 3). Esto indica que existe una alta tasa de recambio de especies entre sitios de diferentes edades.
En el estrato arbóreo de la categoría IV, la familia Apocynaceae fue la más abundante con el 21% de los individuos perteneciendo todos a una sola especie, la segunda más abundante fue Burseraceae (14%), con una especie y la tercera más abundante, Leguminosae (13%) con seis especies.
En cuanto al estrato arbustivo, se observa que la más alta dominancia ocurrió en la categoría I, con un 21% de individuos pertenecientes a la familia más abundante que fue la Malpighiaceae, representada con una sola especie (cuadro 4).
El dendrograma de similitud de familias muestra un patrón similar al de especies, solamente que en este caso todos los valores de similitud son más altos que con las especies. En este caso las categorías de edad más similares fueron la III y la IV (figura 4).
Observando la composición de leguminosas, se puede notar que los sitios con más años de abandono tienen un mayor porcentaje de especies en la subfamilia Papilonoideae, en tanto que en los sitios en uso o de abandono reciente, es la subfamilia Mimosoideae la más ricas en especies.
La subfamilia Caesalpinoideae resultó en general la menos rica, y mantuvo un bajo porcentaje de especies en todos los sitios (figura 5).

DISCUSIÓN
En este trabajo se observa que no todas las variables estructurales presentaron una tendencia monótona a aumentar con el tiempo de abandono. De hecho, en las que sí presentaron este comportamiento, solamente se observaron diferencias significativas en la altura de los árboles.
El tamaño (aquí representado por la altura) puede servir de indicador de la edad de los individuos, y por ende del tiempo que tiene el sitio sin ser talado (Moughalou, 2006). Sin embargo en este caso la altura que se midió fue un promedio de la de los árboles de cada cuadrante lo que podría causar que un sitio con pocos árboles, pero muy maduros sean clasificados como con más de 10 años.
Esto hace necesario combinar la altura con otras variables, como la riqueza del estrato arbóreo o la abundancia de arbustos, que también mostraron diferencias significativas entre los diferentes grupos de edad (Medellín et al., 2000, Moughalou, 2006) y una tendencia (aunque no significativa) a aumentar con el tiempo de abandono.
La riqueza del estrato arbóreo, aunque aumentó de manera monótona con el grupo de edad, no presentó diferencias significativas entre las tres categorías más antiguas. Por ello es necesario combinar esta variable con la altura del estrato arbóreo.
Es notoria la alta abundancia de individuos en el estrato arbóreo en la categoría II, pues fue significativamente mayor que en las categorías III y IV; esto no tiene una fácil explicación teórica y quizá se esté observando un fenómeno particular del área muestreada, resultado de su historia (uso menos intenso, tolerancia del usuario hacia individuos arbóreos) y ubicación geográfica (si es atravesado por aves y murciélagos que desechan semillas y propágulos), que modifican el efecto del tiempo de abandono.
Así mismo resulta interesante notar que la cobertura de árboles en la categoría II resultó significativamente más alta que la de la categoría III. Esto se debe a que la cobertura suele estar altamente correlacionada con la abundancia.
También es interesante notar que pese a tener más individuos, la categoría de edad II, tuvo una menor (aunque no significativamente) cobertura arbórea que el sitio IV, y que la diferencia (medida con la varianza) con la categoría III fue menor que en el caso del número de árboles.
Esto sugiere que el número de individuos en el estrato arbóreo no fue tan buen indicador de la madurez, ya que, como se mencionó, la tasa de ganancia de individuos puede estar influida por factores independientes del tiempo; mientras que en la cobertura, el tiempo puede ejercer una influencia más fuerte, pues combina los efectos del crecimiento individual con el de la ganancia de individuos.
El comportamiento de la riqueza de arbustos también es notable; primero porque existen diferencias entre todas las categorías de edad y segundo por el descenso en el paso de la categoría III a la IV. Explicado por un aumento rápido de especies en las primeras etapas de sucesión, seguido por un ligero descenso en las etapas más avanzadas (Moughalou, 2006, Rosenzweig, 1995); quizá éste sea el patrón observado en el estrato arbustivo, que suele estar conformado por especies de crecimiento rápido, alta capacidad de dispersión y por ende de colonización (Mueller–Dumbois, 2006, Orellana, 1999) pero que se ven afectadas por la competencia por luz y espacio con las especies del dosel de los bosques más desarrollados.
Además, en las etapas sucesionales intermedias pueden estar mezcladas especies de aparición temprana con las de etapas posteriores; conforme avanza el desarrollo del bosque, las especies de aparición más tempranas disminuyen por la exclusión de las más tardías, que suelen ser más competitivas y/o con mayor tolerancia a la sombra del dosel y otras fuerzas causantes de estrés (Walker y Del Moral, 2003).
El análisis discriminante (AD) confirma las tendencias observadas en los ANOVA; las variables que resultaron mejores indicadoras del grupo de edad fueron las que según los ANOVA presentaron diferencias significativas entre las cuatro (o en un caso tres) categorías de años de abandono.
En el plano ecológico resultó interesante revelar que las mayores diferencias con la edad se dan en la altura y abundancia del estrato arbóreo, así como en la riqueza de especies en el estrato arbustivo. Estas variables son indicadoras indirectas de los cambios funcionales de todo el ecosistema, pues la altura y abundancia de árboles son características determinantes del hábitat de grupos tales como las aves, reptiles y otros vertebrados (Medellín et al., 2000), y la riqueza y diversidad de arbustos proporciona una mayor diversidad de recursos para herbívoros, frugívoros y visitantes florales entre otros (Hernández–Betancourt, 2003, Proctor et al., 1996).
Según el análisis discriminante la altura del estrato arbóreo fue la mejor para distinguir entre sitios de diferente grupo de edad, estadísticamente esta variable resultó ser una buena indicadora de la madurez de la vegetación pero ya se mencionaron anteriormente sus limitaciones. Para describir los cambios en la estructura de la vegetación se deben incluir otras variables tales como la diversidad o la cobertura de árboles y arbustos, que son determinantes en la calidad del hábitat de otros organismos.
Las variables estructurales aquí estudiadas son consideradas como indicadoras de la madurez en ecosistemas arbolados, pues se adquieren después de muchos años de sucesión, pero se pueden perder muy rápidamente con la perturbación, especialmente si una gran extensión de hábitat es modificada para usos agrícolas (Parkes et al., 2003, 2004). Sin embargo, la correlación entre cada indicador y la edad del fragmento no siempre es perfecta, ya que además del tiempo pueden intervenir factores tales como la historia del uso del suelo, la disponibilidad de propágulos para recolonizar claros recién abiertos y la distancia a fragmentos de vegetación no perturbada, entre otros (Parkes et al., 2004).
Los bajos índices de similitud en general, sugieren una alta tasa de recambio total de especies; este recambio de especies se puede atribuir a dos factores principales: el espacio y el tiempo, ya que los cuadrantes de la misma edad estuvieron en promedio más cerca entre ellos que con cuadrantes de otra categoría. No obstante los sitios III y IV fueron más parecidos entre sí, pese a que la distancia entre ellos fue la mayor.
Para distinguir entre el efecto del espacio y el de la categoría de edad, se debería comparar este muestreo con otro diseñado para evaluar la heterogeneidad espacial; por desgracia los estudios espaciales a escala intermedia demandan un esfuerzo muy grande, por lo que la mayoría de los estudios que existen son a escalas más pequeñas (1 ha o menos) o suficientemente grandes como para poder usar imágenes aéreas y sistemas de información geográfica (Dale et al., 2002).
En cuanto al recambio temporal de especies, es posible que muchas especies típicas de bosques más maduros no hayan colonizado aún las áreas de más de diez años, ya que estas especies suelen tener limitaciones por la disminución de dispersores de semillas en ambientes perturbados (Martínez–Garza y Howe, 2003), bajas tasas de crecimiento o incluso ser excluidas del pool de especies en selvas con historia de uso muy intenso y extenso (Lamb et al., 2005), y lo que se esté observando en este trabajo sean los cambios de las etapas intermedias de la sucesión.
En el dendrograma de familias, a diferencia de lo observado en el dendrograma de especies, la rama formada por las categorías III y IV presentó un mayor índice de similitud que la rama formada por las categorías I y II es decir, el recambio de familias disminuyó con la edad. La explicación matemática es que al haber menos familias que taxa inferiores, es menos probable que una especie recién llegada a un sitio pertenezca a una familia diferente de las ya representadas.
Esto tiene consecuencias en la biodiversidad taxonómica del paisaje, encontrando que entre mayor sea la extensión de la perturbación, menor será la representación de familias que aparecen más tardíamente, con los consecuentes efectos en las funciones ecológicas.
Con el aumento del tiempo de abandono, se observa una tendencia a la disminución de la proporción de individuos de la familia más dominante, tanto en árboles como en arbustos; esto lleva a un aumento en la distancia taxonómica promedio entre individuos (Magurran, 2004). Un incremento de la distancia taxonómica de los árboles y arbustos puede tener profundas consecuencias ecológicas para la comunidad; pues se aumenta la capacidad de proveer de recursos a una mayor variedad de herbívoros, frugívoros y polinizadores, así como de generar una mayor heterogeneidad de microambientes propicios para el establecimiento de epifitas y otros organismos (Gaston, 1996, Peetchey y Gaston, 2006), así como influir en la heterogeneidad fenológica de plantas y animales (Belaoussof y Kevan, 2003).
Se ha relacionado la diversidad taxonómica con propiedades tales como la resiliencia y la productividad primaria de la vegetación (Tilman et al., 2001), teóricamente, por la capacidad que tiene un mayor número de taxa de llenar más nichos ecológicos y explotar los recursos (por ejemplo luz, nutrimentos y agua) de manera más eficiente. En ambientes estresantes como los sistemas semiáridos y con suelos tan delgados de las selvas secas, la diversidad funcional puede estar relacionada con la capacidad de aprovechar la humedad de diferentes capas de suelo, habilidades diferentes en el aprovechamiento y reciclaje de nutrientes (Escamilla et al., 2005) e interacciones de competencia y nodricismo; esta última de suma importancia para la germinación y sobrevivencia de especies que medran en los estratos inferiores de sistemas áridos (Cervera et al., 2006).
Es notorio que las especies más abundantes en el sitio de edad IV, fueron Plumeria rubra (flor de mayo), Metopium brownei (chechem) y Bursera simaruba (chaká) que se citan como especies características de casi todos los tipos de selvas de la península de Yucatán (Flores–Guido y Espejel, 1994, Rzedowski, 1978). Los climas en los que se encuentran estas especies van desde los Aw (cálidos subhúmedos con picos de precipitación en verano) hasta los BS (cálido o muy cálido semiárido) y los tipos de vegetación van desde las selvas medianas perennifolias hasta las selvas bajas caducifolias, y en el caso de M. brownei el matorral costero. Ibarra Manríquez et al., 2002 citan a M. brownei y B. simaruba como especies de distribución pan peninsular.
Las tres especies son toleradas e incluso promovidas por los habitantes del área maya, además de que toleran bien los climas semiáridos y se regeneran favorablemente de perturbaciones como las quemas y la destrucción asociada con la explotación maderera o agrícola (Macario et al., 1995).
Estas características, sugieren que hasta en los sitios con más años de abandono, aún se detectan los efectos de perturbaciones constantes, ya sean naturales o artificiales. Esto puede deberse a que toda la flora de la región de estudio (el pool de especies) lleva muchas generaciones siendo seleccionada por perturbaciones naturales o artificiales.
El chaká, Bursera simaruba tiene una gran importancia ecológica en todas las comunidades en las que se encuentra, ya que sus frutos y semillas proporcionan alimento a una gran variedad de aves y mamíferos como murciélagos, primates y roedores, por ejemplo se ha encontrado que las semillas de esta especie fueron las más dispersadas por el ratón de abazones Heteromys gaumeri, en selvas medianas del sur de Yucatán (Hernández–Betancourt, 2003).
La flor de mayo (Plumeria rubra), es un arbusto reportado para varios tipos de bosque tropicales, aunque no tantos como las otras dos especies mencionadas. Es notorio que al igual que ellas, pierda sus hojas durante la estación seca y sea resistente a los climas semiáridos. Presenta buena regeneración, pues al igual que el chaká, los individuos caídos pueden producir nuevas raíces y se puede regenerar con estacas. La flor de mayo es una especie tolerada y hasta propiciada en los solares y huertos familiares por su importancia cultural y ornamental. De manera que su abundancia puede indicar también una historia de uso intenso.
En cuanto al análisis de la familia Leguminosae, se encontró que ésta fue la más rica en especies en el sitio de estudio. Según Flores–Guido (2001) ésta es la familia más importante en la composición de la vegetación en la Península de Yucatán.
En los sitios estudiados, se encontró que la riqueza de Papilonoideae fue la más alta en los dos sitios con la categoría de edad más alta; cabe destacar que Flores–Guido, 2001 reporta que ésta es la subfamilia con más especies en las selvas de la Península de Yucatán.
En contraste, la subfamilia Mimosodeae fue la más rica en los sitios de uso más reciente; esto se puede atribuir a que en Yucatán, en esta subfamilia se agrupan varias especies de vida corta y muy dispersivas que aparecen desde el primer año de regeneración del monte, tras el abandono de tierras agrícolas (Flores–Guido, 2001).
La enorme proporción de especies e individuos de Leguminosae, se explica por la historia de manejo del monte; pues según Gómez Pompa (1971) citado en Flores–Guido (2001), muchas especies de esta familia pueden medrar en suelos empobrecidos por el lixiviado de nutrientes típico de las explotaciones agrícolas, gracias a que establecen una interacción simbiótica con bacterias fijadoras de nitrógeno.
De esta forma, la disminución de las leguminosas en las abundancias relativas de los cuadrantes de edad IV, es decir de más de 10 años, podría deberse a que los bosques tienden a acumular nutrientes conforme se acercan a la "madurez" (Krebs, 1994). Se supone que los procesos de reciclamiento de nutrientes se hacen más eficientes, y disminuye la lixiviación. Aun así, las leguminosas fueron la tercera familia en abundancia y la primera en riqueza en los sitios de edad IV.
Se puede concluir que aunque se observan importantes cambios en la diversidad taxonómica y estructura de la comunidad, las características de los sitios más antiguos (especies resistentes a las perturbaciones, alta abundancia de leguminosas y altas tasas de recambio de especies en etapas avanzadas de abandono) sugieren que no se ha alcanzado el llamado "clímax" de la vegetación.
AGRADECIMIENTOS
Los recursos económicos de este proyecto fueron aportados por Fondos Mixtos CONACYT–Yucatán mediante el proyecto 2004–09–30/2005–12–15 "Las Aves como indicadoras de la pérdida de Biodiversidad en la Reserva de Dzilam".
Los autores agradecen la colaboración de habitantes y autoridades de la reserva por su paciencia y generosidad. A los revisores y editores de esta revista por sus valiosos comentarios para mejorar el manuscrito.
LITERATURA CITADA
Bautista–Zúñiga, F., Batllori–Sampedro, E. Ortiz–Pérez, M.A., Palacio–Aponte, G. y Castillo–González, M., 2003. "Geoformas, agua y suelo de la Península de Yucatán". En: Colunga–GarcíaMarín, P y Larqué–Saavedra, A. (Editores). Naturaleza y Sociedad en el Área Maya. Academia Mexicana de Ciencias–CICY. pp. 21–36. [ Links ]
Belaoussof, S. & Kevan, P., 2003. "Are there ecological foundations for ecosystem health". The Environmentalist, 23: 255–263. [ Links ]
Carnevali, G., Ramírez, I.M. y González–Iturbide, J.A., 2003. "Flora y Vegetación de la Península de Yucatán". En: Colunga–García Marín, P y Larqué–Saavedra, A. (Editores). Naturaleza y Sociedad en el Área Maya. Academia Mexicana de Ciencias–CICY. pp 53–68. [ Links ]
Cervera, J.C., Andrade, J.L., Simá, J.L. y Graham, E.A., 2006. "Microhabitats, germination and establishment requirements for Mammillaria gaumeri (CACTACEAE) a rare species from Yucatán". J. Pl. Sci., 167: 311–319. [ Links ]
CNA, 2000. Datos Climatológicos por estación: Yucatán. www.cna.gob.mx [ Links ]
Dale, M.R.T., Dixon, P., Fortin, M.J., Legendre, P. Myers, D.E. & Rosenberg, M.S., 2002. "Conceptual and mathematical relationships among methods for spatial analysis". Ecography, 25: 558–577. [ Links ]
Escamilla, A., Quintal, F., Medina, F., Guzman, A. Pérez, E. y Calvo, L.M., 2005. "Relaciones suelo–planta en ecosistemas naturales de la Península de Yucatán: Comunidades dominadas por palmas". En: Bautista–Zúñiga, F. y Palacio, G. (Editores). Caracterización y manejo de los suelos de la Península de Yucatán, Implicaciones agropecuarias y ambientales. Universidad Autónoma de Campeche–Universidad Autónoma de Yucatán, pp. 159–172. [ Links ]
Flores–Guido, J.S., 2001. Leguminosae, florística, etnobotánica y ecología. Universidad Autónoma de Yucatán. Etnoflora Yucatanense, Fascículo 18. 320 pp. [ Links ]
Flores–Guido, J.S. y Espejel, I., 1994. Tipos de Vegetación de la Península de Yucatán. Universidad Autónoma de Yucatán. Etnoflora Yucatanense, Fascículo 3. 135 pp. [ Links ]
Gaston, K.J., 1996. "Species richness measure and measurements". En: Gaston K. J. (Editor) Biodiversity a Biology of Numbers and Difference. pp. 77–113. [ Links ]
Gobierno del Estado de Yucatán, 2006. Ficha técnica de la Reserva Estatal de Dzilam. Secretaría de Ecología. 18 pp. [ Links ]
Harris, J.A & R.J. Hobbs, 2001. "Clinical practice for ecosystem health: the role for ecological restoration". Ecosys Health, 7: 195–202. [ Links ]
Hernández–Betancourt, S., 2003. Dinámica Poblacional de Heteromis gaumeri (Allen y Chapman, 1897), en una selva mediana del sur de Yucatán, México. Tesis Doctoral en Ciencias Biológicas. Universidad Autónoma Metropolitana. 227 pp. [ Links ]
Ibarra–Manríquez, G., Villaseñor, J.L., Durán, R. y Meave, J.,2002. "Biogeographical análisis of the tree flora of the Yucatán Peninsula". J. Biogeography, 29: 17–29. [ Links ]
Kovach Computing Services, 1999. MVSP 3.10b. [ Links ]
Krebs, C.J., 1994. Ecology, the Experimental Analysis of Distribution and Abundance. Harper Collins. [ Links ]
Lamb, D., P.D. Erksine, y J.A. Parotta, 2005. "Restoration of tropical forest landscapes". Science, 310: 1628–1632. [ Links ]
Macario, P., E. García–Moya, J.R. Aguirre–Rivera y E. Hernández–Xolocotzi, 1995. "Regeneración natural de especies arbóreas en una selva mediana subperennifolia perturbada por una extracción forestal". Acta Bot. Mex., 32: 11–23. [ Links ]
Magurran, A.E., 2004. Measuring biological diversity. Blackwell Scientific Publications. [ Links ]
Martínez–Garza, C. & Howe, H.F., 2003. "Restoring tropical diversity: beating the time tax on species lost". J. Appl. Ecol., 40: 423–429. [ Links ]
Martínez–Arias, R., 1999. El análisis multivariante en la investigación científica. Cuadernos de Estadística Fascículo 1. Editorial la Muralla. 143 pp. [ Links ]
Medellín R.A, M. Equihua & M.A. Amín, 2000. "Bat diversity and abundance as indicators of disturbance in neotropical rainforests". Conserv. Biol., 14: 1666–1675. [ Links ]
Moughalu, J.I., 2006. "Tree species population dynamics in a secondary forest at Ile–Ife. Nigeria after a ground fire". Afr. J. Ecol., 45: 62–71. [ Links ]
Mueller–Dumbois, D., 2006. "Long–term sucesión and landscape change in Hawai'i: The 'Maui forest trouble' revisited". J. Veget. Sci., 17: 685–692. [ Links ]
Orellana, R., 1999. "Respuestas de las plantas al ambiente por medio de sus estrategias morfológicas y funcionales". En: Orellana, R., J.A. Escamilla y A. Larqué Saavedra (Editores). Ecofisiología vegetal y conservación de recursos genéticos. CICY. pp. 35–50. [ Links ]
Orellana, R., Islebe, G. y Espadas, C., 2003. "Presente, pasado y futuro de los climas de la Península de Yucatán". En: Colunga–García Marín, P y Larqué–Saavedra, A. (Editores)". Naturaleza y Sociedad en el Área Maya. Academia Mexicana de Ciencias–CICY. pp. 37–52. [ Links ]
Parkes,D.,G. Newell & D. Cheal, 2003. "Assessing the quality of native vegetation: the habitat hectares approach". Ecological Management and Restoration, 4: S29–S38. [ Links ]
––––––––––, 2004. "The development and raison d'être of 'habitat hectares': A response to McCarthy et al. 2004". Ecological Management and Restoration, 5: 30–31. [ Links ]
Peetchey, O.L. & K.J. Gaston, 2006. "Functional diversity: back to basics and looking forward". Ecol. Letters, 9: 741–758. [ Links ]
Proctor, M., Yeo, P. & Lack, A., 1996. "The natural history of pollination". Harper Collins Publishers. 479 pp. [ Links ]
Rosenzweig, M.L., 1995. Species Diversity in Space and Time. Cambridge. University Press. 436 pp. [ Links ]
Rzedowski, J., 1978. Vegetación de México. LIMUSA, México. 432 pp. [ Links ]
Tilman, D., J. Knops, D. Wedin & P. Reich., 2001. "Experimental and observational studies of diversity, productivity and stability". En: Kinzig, A.P, S.W. Pacala y D. Tilman (Editores). "The functional consequences of biodiversity". Princeton University Press. Monographs in Population Biology, 33: 42–52. [ Links ]
UADY., 1999. Atlas de Procesos Territoriales del Estado de Yucatán. Universidad Autónoma de Yucatán. [ Links ]
Walker, L.R. & R. Del Moral, 2003. Primary succession and ecosystem rehabilitation. Cambridge University Press. 442 pp. [ Links ]

