










Serviços Personalizados
Journal

Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkPolibotánica
versão impressa ISSN 1405-2768
Polibotánica no.27 México Abr. 2009
Small–scale environmental gradients in a pine–oak forest community in Nueva Colonia, Mezquitic, Jalisco, Mexico
Gradientes ambientales a pequeña escala de un bosque de pino–encino en Nueva Colonia, Mezquitic, Jalisco, México
Gregorio Nieves–Hernández1, J. Antonio Vázquez–García1, Yalma L. Vargas–Rodriguez2, Marcelino Vázquez–García1 and Jesús González–Gallegos1
1 Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara, Km 15, carretera Guadalajara–Nogales, Las Agujas, Zapopan, Jalisco, 45110, México. Correo electrónico: jvazquez@cucba.udg.mx
2 107 Life Sciences Building, Department of Biological Sciences, Louisiana State University, Baton Rouge, Louisiana, 70803, USA.
Recibido: 6 febrero 2008
Aceptado: 2 diciembre 2008
Abstract
We explored environmental variables related to pine–oak forest community structure at one locality in Jalisco, Mexico. We used an NMS ordination in conjunction with the Sørensen distance to identify the major small–scale community gradients along 25 contiguous quadrats (25 m x 25 m, 400 m2 each) of pine–oak forest in Nueva Colonia, Mezquitic, Jalisco, Mexico. The main matrix (25 stands x 7 tree species) included basal area data, and the environmental matrix consisted of 19 quantitative environmental variables. Community structure, through sociological ordination, showed a direct correlation with the vertical altitudinal gradient and apparent soil density, as well as with slope inclination across the horizontal gradient; it also showed an inverse correlation with cation exchange capacity, Ca + Mg, Mg, and altitude across the horizontal gradient. Direct gradient analyses showed an increase of cation exchange capacity, Ca+Mg, Mg and K with decreasing altitude along the vertical gradient (from north to south). Total nitrogen increased with decreasing altitude across the horizontal gradient (from west to east). In addition, we identified three main community groups using UPGMA cluster analysis; however, groups were weakly related to the ordination results and to the physical space. At the 1–hectare scale, species composition and basal area of pine–oak forest in Nueva Colonia can be explained by niche partitioning of altitude and soil gradients. The relevance of nitrogen for this community could be the result of habitat specialization or disturbance history.
Key words: calcium, magnesium, environmental gradients, habitat differentiation, niche partitioning.
Resumen
Nosotros exploramos cuáles variables ambientales se relacionan con la estructura de comunidades de pino–encino en Nueva Colonia, Mezquitic Jalisco, México. A través de una ordenación NMS y usando la distancia Sørensen se identificaron en la escala pequeña los principales gradientes de comunidades entre 25 parcelas contiguas (25 x 25 m, cada una de 400 m2) de bosque de pino–encino. La matriz principal (25 rodales x 7 especies de árboles) incluyó datos de área basal y la matriz ambiental consistió de 19 variables ambientales cuantitativas. La estructura de la comunidad, mediante la ordenación sociológica, mostró una correlación directa, sobre el gradiente vertical, con la altitud y con la densidad aparente del suelo; y, sobre el gradiente horizontal, con la inclinación de la pendiente; también mostró una correlación inversa, sobre el gradiente horizontal, con la capacidad de intercambio catiónico, Ca + Mg, Mg y con la altitud. El análisis de gradientes directos mostró, en el gradiente vertical (de norte a sur), un incremento de la capacidad de intercambio catiónico, Ca+Mg, Mg y K con la disminución de la altitud. El nitrógeno total, en el gradiente horizontal (de oeste a este), incrementó con la disminución de la altitud. Además, se identificaron tres grupos principales de comunidades usando el análisis de conglomerados de UPGMA, sin embargo, los grupos mostraron una relación débil con los resultados de la ordenación y con el espacio físico. A la escala de una hectárea, la composición de las especies y el área basal del bosque de pino–encino en Nueva Colonia pueden ser explicados por la partición de nicho en gradientes de altitud y de suelo. La relevancia del nitrógeno para esta comunidad podría resultar de la especialización de hábitat o del historial de disturbio.
Palabras clave: calcio, magnesio, gradientes ambientales, diferenciación de hábitat, partición de nicho.
INTRODUCTION
Many studies of large–scale plant community patterns have supported the individualistic hypothesis of plant community organization (Gleason, 1926) in both, temperate areas (Whittaker, 1956, 1960; Curtis and McIntosh, 1951; Peet, 1980) and subtropical to tropical areas (Vázquez–García, 1995, 1998; Cuevas, 2002), some other authors document vegetational discontinuities (Beals, 1969; Kitayama, 1992). However, studies of small–scale community patterns to assess whether or not the individualistic hypothesis is supported, have largely been neglected.
Understanding patterns of species distribution and abundance across landscapes have been central goals in Ecology. Ecological studies have focused on determining whether species specialize to partition environments or whether distributions are the result of stochastic processes (Brokaw and Busing, 2000; Hubbell, 2001). A number of mechanisms have been proposed to explain how diversity and abundance are regulated in tropical and temperate systems (Wills et al., 1997; Wright, 2002). Niche differentiation among species, arising from resource partitioning across resource gradients, has been hypothesized to be a major mechanism maintaining tree diversity in both temperate and tropical forests. Many studies (Denslow, 1980; Kabakof and Chazdon, 1996; Chazdon et al., 1999; Capers et al., 2005; iarte and Chazdon, 2005; Gouvenain et al., 2007) have focused only on light gradients as driving force or mechanism in resource partitioning among tree species, however, little or no attention has been given to other resources such as soil variables.
Climatic variables might determine plant community distribution at a landscape level. Latitude, longitude, precipitation, temperature, and moisture variables have an effect in plant composition and distribution (Rzedowski, 1978, Gentry, 1982). However, at small–scale gradients (< 200 m in altitude), topography is considered a more relevant variable that affects tree community spatial variation rather than it is altitude or any other climatic variable (Basnet, 1992; Vázquez–García, 1995). Topography is also related to patterns in moisture regimes and soil chemical properties (Oliveira–Filho et al. 1999, 1994; Ratter, 1980; Bourgeron, 1983; Johnston, 1992). For instance, soil leaching patterns along an elevational gradient are affected by topography (Smith, 1990).
Soil characteristics and biotic interactions, at a small scale (within a hectare), might be more important determining plant community patterns (Fowler, 1981). The soil organic horizon is related to chemical composition, biological activity, fertility, and soil development. Soil nutrients distributions vary at a small–scale, affecting plant development and species distributions. For instance, nitrogen availability commonly limits plant growth in forested ecosystems (Aber et al., 1989). In upland forests, N mineralization rates have been related to tree species composition (Pastor et al. 1982; Zak et al., 1986; Garten et al., 1994) and productivity (Pastor et al., 1984; Liu and Muller, 1993).
Habitat specialization for soil resources in a tropical dry forest is likely to be more important at early stages in tree life histories than it is in later life history (Vargas–Rodríguez et al., 2005). This fact is very relevant for understanding tree establishment and regeneration and could help guide land managers in implementing restoration policies. However, environmental variables explaining main small–scale gradients in community composition of pine–oak forests in northern Jalisco have not been explored yet. Tree community variation needs to be evaluated at a much larger scale, however community variation of early stages in tree life histories (e.g. saplings) may be studied at smaller scales. Here we use 25 quadrats (0.04 ha) within 1 ha plot to focus on tree communities patterns at early stages in life tree histories (woody species > 2.5 cm in diam.) within a small area, with the advantage of low habitat and climatic heterogeneity.
In the present study, we aim to generate hypotheses, rather than to test them, about what environmental gradients are evident at a hectare plot and which environmental variables possibly explain small–scale community gradients at early stages of tree life histories of a pine–oak forest in Nueva Colonia, Mezquitic, Jalisco, Mexico.
METHODS
Study area
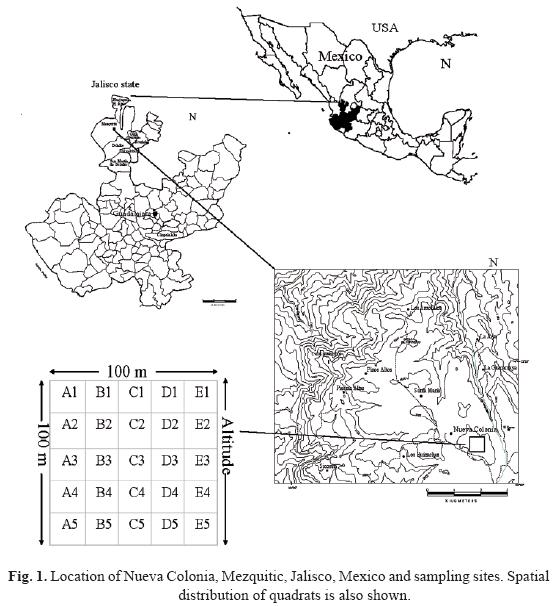
The study area was located at 6 km northeast of Nueva Colonia, southeast of Mezquitic, Mezquitic municipality, Jalisco, Mexico. We located a hectare plot within the tributary watershed Los Coyotes, along an 1800–2400 altitudinal gradient, at the southern portion of Sierra Madre Occidental (Fig. 1). The climate at Nueva Colonia was temperate with cold and moist winters (Vázquez–García et al., 2004). Mean annual rainfall was 600 mm with the wet season from July to August. Relief ranged from 5–20% slope, of southern aspect. Volcanic acidic extrusive rocks and igneous Toba rocks were prevalent in the area. Silt soils and ferric Luvisol soils were common in the area (INEGI 1970a, 1970b).
Floristic checklists for the municipality included over 373 vascular plant species (Vázquez–García et al. 2004). The dominant vegetation was pine and pine–oak forests. In accordance with vegetation class, the most common species in the area were Pinus oocarpa, P. devoniana, P. durangensis, P. duglasiana, Quercus castanea, Q. crassifolia, Q. splendens, Arbutus xalapensis, Befaria mexicana, Alnus jorullensis and Juniperus flacida (Vázquez et al., 1999; Vázquez and Vargas, 1999; Vázquez–García et al., 2004).
Field sampling
We selected the least disturbed (forest stands with larger dbh) pineoak forest in Nueva Colonia through field reconnaissance. The huichol community has designated this forest as sacred and no logging is allowed in this area. The total area sampled was 1 hectare (100 m x 100 m). We divided the hectare in 25 quadrats, each one was 400 m2. The altitudinal variation within the hectare ranged from 2353 to 2367 m (14 m difference in elevation or 14% slope degree). To prevent confusion, we assigned a number to each quadrat, following a west–east and north–south order (first row A1–A5, second row B1–B5, and so on) (Fig. 1). At each quadrat, we recorded plant species > 2.5 cm DBH and collected a voucher of each species. The sampling was conducted during the summer of 2002 (late June–early July). Species' identifications were determined by the authors. Nomenclature generally follows Vázquez–García et al. (2004). Herbarium specimens were deposited at the The University of Guadalajara's herbarium (IBUG) (Holmgren et al., 1990).
We collected a total of ten composite soil samples. Each of them represented five quadrats, either from a row within the elevational gradient or from a column along the elevational gradient. All of the samples were selected from one hectare's grid with twenty five 20 x 20 m. squares. The composite soil samples were obtained to increase soil representation in the area. Soil density, texture (sand, clay, silt), soil moisture, organic matter, cation exchange capacity (CEC), Mg, Ca, Ca+Mg, Na, K, pH, N, and P were analyzed (Table 1). Nutrients were measured following Mehlich III's extraction method, P was extracted with the Olsen method, pH with a potentiometer, soil moisture with the gravimetric method, organic matter with Walkey–Black's process, and texture with Bouyoucos' methods (AOAC International, 1990; APHA–AWWA–WPCF, 1992; Agricultural Experiment Stations, 1998). We recorded site conditions within the plot and determined position and altitude using a global positioning device (GPS 12 Garmin corporation) and aspect and slope using a compass and a clinometer, respectively. Rockiness was also noted visually as a percent of a quadrat covered and averaged for a row or a column of quadrats.
We measured the extent of natural and anthropogenic disturbances at each site. We assessed the degree of disturbance by recording the number of standing dead trees, fallen trees, woody stems, and tree stumps. Cattle grazing and fire evidence were also recorded (Olvera–Vargas et al., 1996). Average disturbance values were used to assign one of three disturbance classes (high, intermediate or low) for each column or row of quadrats.
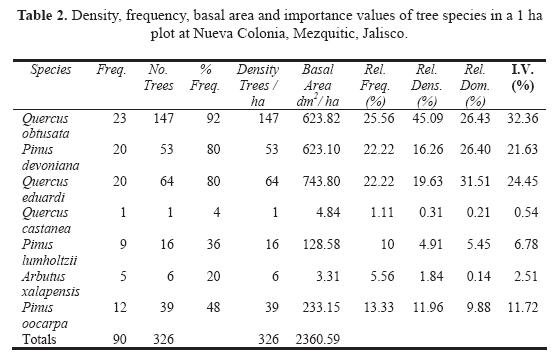
We obtained species basal area for each of the 25 quadrats and summarized them in one matrix, containing seven tree species and 25 quadrats (Table 2). The environmental variables were summarized in a matrix with 19 quantitative variables and 25 quadrats.
Environmental gradients
Using direct gradient analysis, we identified major environmental gradients and how they were related to altitude along the vertical gradient (from north to south) and to the horizontal gradient (from west to east), averaging data on variables for each row or column of quadrats.
Sociological ordination
Matrices were analyzed using Global Non–metric Multidimensional Scaling (NMS) ordination, available in PCORD v.4, a software that performs multivariate analyses for ecological data (McCune and Mefford 1999). NMS was performed following the method described by Kruskal (1964) and Mather (1976). NMS is an effective ordination method for ecological community data because it does not assume linear relationships among variables (Minchin, 1987). The method searches for the best position on n entities on k axes with the objective of minimizing the stress on the k–dimensional configuration (Minchin, 1987; McCune and Grace, 2002).
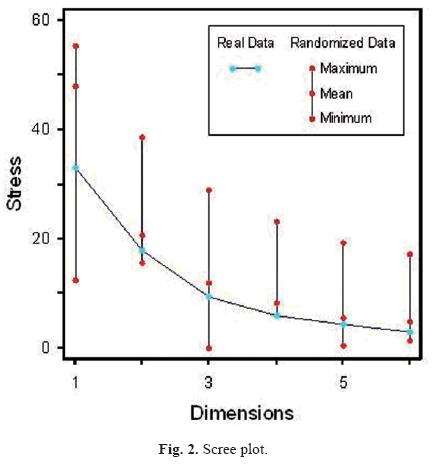
A general procedure for NMS was used as suggested by McCune & Grace (2002): a) data were adjusted, Quercus castanea occurred in less than 5% of sample units and was removed as a rare species; sample unit E2 was identified as a moderate outlier an thus removed; given the range of basal area values for each species, there was no need for their standardization; raw data were preferred over relativized ones for an appropriate representation of shifts in species structure (e.g. increases in basal area), b) a preliminary configuration with 100 runs was employed to asses how possible would the observed pattern be in the ordination if only a random process would be operating at the community configuration, this step suggested a three dimensional model; c) dimensionality of the data set was determined by plotting a scree plot, which shows stress as a function of dimensionality of the gradient model (Fig. 2); d) three dimensions were specified; e) a plot of stress vs. iteration assisted in checking for stability; f) the final configuration included the Sørensen distance measure, 3 axes, number five was used as a seed for the random number generator, with no step down dimensionality, one real run, and no Monte Carlo test (no randomized runs).
The Sørensen's distance measure was used, a robust measure for ecological distance (Beals, 1984; Faith et al., 1987). The relationship between tree species and environmental variables was evaluated using Pearson correlations between the calculated ordination axes and the environmental variables. Only variables from the secondary matrix with an r2 (with scores on either axis) larger than the cutoff value were plotted. The default cutoff is 0.2, but it was set to 0.3 to overlay few and relevant vectors.
Classification
We classified the studied forest plots using the grouping algorithm of Unweighted Pair Group Average (UPGMA). We used UPGMA in connection with the Sørensen distance S:
S = 1 – 2w/(a+b)
where a and b are the numbers of species in each of two samples and w is the number of species the samples share. UPGMA is a hierarchical polythetic agglomerative technique that groups samples into classes and classes into a hierarchy, using information simultaneously from all samples. Each sample is assigned to a cluster on a hierarchy of increasingly inclusive clusters until all samples are part of one cluster. Opposed to related classification techniques, UPGMA's dissimilarities between clusters are equal to the average dissimilarity rather than equal to the maximum dissimilarity (Orlóci, 1978). UPGMA maximizes the correlation between input and output compositional dissimilarities implied in the obtained dendrogram (Gauch 1982).
RESULTS
A total of 326 individual trees were recorded in the studied area (1 ha, 25 plots). We found seven tree species: Pinus devoniana, P. lumholtzii, P. oocarpa, Quercus obtusata, Q. eduardii, Q. castanea and Arbutus xalapensis. The tree diversity index value was H' = 1.483. Oak species were the dominant elements in the forest. Q. eduardii had the highest basal area (Table 2). Q. obtusata, Q. eduardii, and P. devoniana had the highest importance values (32%, 24%, and 22% respectively). Total basal area in the plot was 24 m2 ha (Table 2). Limited anthropogenic disturbance occurs in the study area by restricting visitors to the sacred forested area. We were told that the last wood selective extraction occurred about 30 years ago. Four percent of trees had fire scares. Evidence of past forest fires is attributable to lightening (local inhabitants, pers. comm.). There was no evidence of cattle grazing or insect predation, at least on tree saplings or adults.
Environmental ordination
Regression analysis showed that CEC, Ca + Mg, Mg and K increased with decreasing elevation, which represents an increase of nutrients from north to south (Fig. 3). Percent of total–nitrogen increased from west to east within elevation (Fig. 4).
Sociological ordination
The scree plot (Fig. 2) and the Monte Carlo test recommended a three dimensional solution. The proportion of randomized runs with stress less than or equal to the observed stress was 0.0495. The final stress for a three–dimensional solution of the NMS ordination was 9.718. Axes accounted for a reduction in stress of 0.0001 and instability of 0.06427 with 100 iterations. Most of the stress was reduced after 20 iterations. Axes 1 and 2 were not explained by any of the measured environmental variables. However, axis 3 was directly related to altitude along the vertical gradient, apparent density and the slope of the horizontal gradient (Fig. 5a); and inversely related to CEC, Mg, Ca + Mg, and altitude along the horizontal gradient (Fig. 5a). In relation to species patterns, axis 1 was directly related to basal area of Pinus oocarpa and inversely related to basal area of P. devoniana (Fig. 5b); axis 2 was directly related to basal area of Quercus obtusata (Fig. 5c); axis 3 was inversely related to basal area of Q. eduardii (Fig. 5d). Environmental variables explaining major community gradients in the sociological ordination were largely consistent with those identified through direct gradient analyses.
Classification
UPGMA recognized three groups containing 25% of the remaining information. A heterogeneous group is composed by A1, B1, B2, C3 y C4 quadrats, which are spatially correlated. A second heterogeneous group contains B3, C1, D1 and E2, which are spatially separated. The third group containing 16 quadrats was the largest, it was less heterogeneous, and spatially separated (Fig. 6). Classification results did not reproduced physical location of quadrats, and were weakly related to ordination results.
DISCUSSION
Community structure of pine–oak forest in Nueva Colonia may be explained by habitat specialization to altitude and soil gradients. Consistently, establishment of pine species has been related to soil and topographic conditions (Park 2001). In addition, species interactions allow differences in species dominance in the community, for instance, Pinus devoniana and P. oocarpa dominated different ends of axis 1; Quercus obtusata dominated axis 2; and Q. eduardii dominated axis 3. Differences in dominance along these axes was explained by CEC and by soil nutrients (Ca + Mg, Mg). Community patterns at the Sierra Huichola support the individualistic hypothesis of plant community organization, with no evidence for vegetational discontinuities.
The fact that five from seven species have an aggregated distribution pattern indicates habitat specialization or spatial autocorrelation (Harms et al., 2001). A correlation analysis with ecological equivalence among quadrants could help to explain species habitat preferences. A third of these species have random distribution, supporting the neutral theory, in which species are habitat independent.
There is a soil–species relationship in pine–oak forest at Nueva Colonia. Soil nutrients and soil moisture appear correlated with temperate tree species distributions (Cowell 1993). Other factors such as topography and moisture conditions might be important structuring forest composition, however, the role of nutrients and disturbance are primary forces controlling vegetation patterns in temperate forests (Peet and Christensen, 1980; Cowell, 1993; Park, 2001; Cavender–Bares et al., 2004). Variation in exchangeable magnesium, calcium, and potassium allow differences in growth and mortality rates of oaks and pine species, resulting in different patterns of species distributions (Breemen et al., 1997; Arii and Lechowicz, 2002; Bigelow and Canham 2002; Dijkstra and Smits, 2002; Horsley et al., 2002). Soil nutrients can change on small scale across topography gradients, thus producing gradients in soil fertility and affecting species dominance (Bailey et al., 2004). At a 1 hectare scale, species maintenance and basal area of oak and pine in Nueva Colonia can be explained by niche partitioning of soil gradients. This competitive interaction for soil resources is a key mechanism structuring the community (Muscolo et al., 2007). Consistently, 17 oak species in Florida show clear patterns of specialization along soil moisture and nutrient gradients and show a pattern of habitat differentiation (Cavender–Bares et al., 2004).
Habitat heterogeneity can be observed at a small scale, as showed in classification. Spatially contiguous sites with low climatic and topographic heterogeneity have small scale differences and have an effect in soil nutrients distribution (Muscolo et al. 2007). The increase in bases at lower elevations might be related to a leaching process. Constant soil leaching has a pronounced effect on Ca and Mg availability (Schier and McQueattie, 2000). Soil leaching occurring in Nueva Colonia resembles the cascade effect (low nutrients at higher elevations increasing at lower) observed in mountains of Sierra de Manantlán (Vázquez and Givnish, 1998; Ortiz–Arrona, 1999). The nitrogen relevance for this community might be the result of disturbance regime stage or habitat specialization. N availability is strongly correlated with vegetation patterns, linking below–ground processes and above–ground biodiversity (Hutchinson et al., 1999). Therefore, species are specialized and distributed in microsites with different nitrogen and other nutrients availability.
ACKNOWLEDGMENTS
The Centro de Ingeniería Ambiental, Unidad de Apoyo a Comunidades Indígenas, both from University of Guadalajara; SIMORE–LOS–CONACYT–1996, through Ordenamiento Ecológico Territorial de Jalisco; and Instituto Nacional Indigenista (SEDESOL) provided financial support for this research. Miguel de J. Cházaro B., Ramón Cuevas G., Mollie Harker, Jacqueline Reynoso D., Roberto González T. and María Eugenia Barba R. helped with species identification. Rafael López de la Torre, wirrarika, kindly helped to locate forest sites and gave us logistic support. Luzmila Herrera Pérez, Justo Murguía Castillo, María Eugenia Barba Robert, and Magdalena Alcázar García collected voucher plant specimens and assisted during field work. We thank Susana and Diana Parker for proofreading the English. Special thanks to anonimous reviewers of Polibotánica and to Ramón Cuevas, from the Manantlán Institute, Universidad de Guadalajara, for their valuable suggestions.
LITERATURE CITED
Aber, J.D., K.J. Nadelhoffer, P.Steudler & J. M. Melillo, 1989. "Nitrogen saturation in northern forest ecosystems". Bioscience, 39: 378–386. [ Links ]
Agricultural Experiment Stations, 1998. "Recommended chemical soil test procedures". North Central Regional Research Publications 221, Missouri Agricultural Experiment Station, Columbia, Missouri. [ Links ]
Allendorf, F.W. & N. Ryman, 2002. "The role of genetics in population viability analysis". In: Beissinger, S.R. & McCullough, D.R. (eds.) Population viability analysis, pp. 50–85. University of Chicago Press, Chicago, Illinois. [ Links ]
AOAC International, 1990. Official methods of analysis: volume I, Association of Official Analytical Chemists SB 1001. Missouri Agricultural Experiment Station, Maryland, Missouri. [ Links ]
APHA–AWWA–WPCF, 1992. Standard methods for the examination of water and wastewater, American Public Health Association, Washington, D.C. [ Links ]
Arii, K. & M.J. Lechowicz, 2002. "The influence of overstory trees and abiotic factors on the sapling community in an old–growth Fagus–Acer forest". Ecoscience, 9(3): 386–396. [ Links ]
Bailey, S.W., Horsley, S.B., Long, R.P. & Hallett, R.A., 2004. "Influence of edaphic factors on sugar maple nutrition and health on the allegheny plateau". Soil Science Society American Journal, 68: 243–252. [ Links ]
Basnet, K., 1992. "Effect of topography on the pattern of trees in tabonuco (Dacryodes excelsa) dominated rain forest in Puerto Rico". Biotropica, 4: 31–42. [ Links ]
Beals, E.W., 1969. "Vegetation changes along altitudinal gradients". Science, 165: 981–985. [ Links ]
––––––––––, 1984. "Bray Curtis ordination: an effective strategy for analysis of multivariate ecological data". Advances Ecol. Research, 14: 1–55. [ Links ]
Bigelow, S.W. & C.D. Canham, 2002. "Community organization of tree species along soil gradients in a north–eastern USA forest". Journal of Ecology, 90: 188–200. [ Links ]
Bourgeron, P.S., 1983. "Spatial aspects of vegetation structure". pp. 29–47. In: F.B. Golley (ed.). Ecosystems of the world: tropical rain forest ecosystems 14. Elsevier, Amsterdam. [ Links ]
Breemen, N. Van, A.C. Finzi & C.D. Canham, 1997. "Canopy tree–soil interactions within temperate forests: effects of soil elemental composition and texture on species distributions". Canadian Journal of Forest Research, 27: 1110–1116. [ Links ]
Brokaw, N. & R.T. Busing, 2000. "Niche versus chance and tree diversity in forest gaps". Tree, 15: 183–188. [ Links ]
Capers, R.S., R.L. Chazdon, Al. Redondo Brenes & B. Vilchez Alvarado, 2005. "Successional dynamics of woody seedling communities in wet tropical secondary forests". Journal of Ecology, 93: 1071–1084. [ Links ]
Cavender–Bares, J., K. Kitajima, & F. A. Bazzaz, 2004. "Multiple traits associations in relation to habitat differentiation among 17 floridian oak species". Ecological Monographs, 74: 635–662. [ Links ]
Chazdon, R.L., R.K. Colwell & J.S. Denslow, 1999. "Tropical tree richness and resource–based niches". Science, 285: 1459a [on–line journal only] [ Links ].
Cowell, C.M., 1993. "Environmental gradients in secondary forests of the Georgia Piedmont, U. S. A". Journal of Biogeography, 20: 199–207. [ Links ]
Cuevas G., R., 2002. Análisis de gradientes de la vegetación de la cañada El Tecolote en La Sierra de Manantlán, Jalisco, México. Tesis doctoral. Colegio de Posgraduados, Montecillo, México. 140 pp. [ Links ]
Curtis, J.T. & R.P. McIntosh, 1951. "An upland forest continuum of the prairie forest border region of Wisconsin". Ecology, 32: 476–496. [ Links ]
Denslow, J.S., 1980. "Gap partitioning among tropical rainforest trees". Biotropica, 12: 47–55. [ Links ]
Dijkstra, F.A. & M.M. Smits, 2002. "Tree species effects on calcium cycling: the role of calcium uptake in deep soils". Ecosystems, 5: 385–398. [ Links ]
Faith, P.D., P.R. Minchin & L. Belbin, 1987. "Compositional dissimilarity: a robust measure of ecological distance". Vegetation, 69: 57–68. [ Links ]
Fowler, C.W., 1981. "Density dependence as related to life history strategy". Ecology, 62: 602–610. [ Links ]
Garten, C.T., M.A. Huston & C.A. Thoms, 1994. "Topographic variation of soil nitrogen dynamics at Walker Branch Watershed, Tennessee". Forest Science, 40: 497–512. [ Links ]
Gauch, H.G., 1982. Multivariate analysis in community ecology. Cambridge University Press. [ Links ]
Gentry, A.H., 1982. "Patterns of neotropical plant species diversity". Annals of the Missouri Botanical Garden, 69: 557–593. [ Links ]
Glitzenstein, J.S., WJ. Platt & D. R. Streng, 1995. "Effects of fire regime and habitat on tree dynamics in North Florida longleaf pine savannas". Ecological Monographs, 65: 441–476. [ Links ]
Gouvenain, R. C., R. K. Kobe & J. A. Silander, 2007. "Partitioning of understorey light and dry–season soil moisture gradients among seedlings of four rain–forest tree species in Madagascar". Journal of Tropical Ecology, 23: 569–579. [ Links ]
Harms, K.E., R. Condit, S.P. Hubbell & R.B. Foster, 2001. "Habitat Associations of Trees and Shrubs in a 50–Ha Neotropical Forest Plot". Journal of Ecology, 89(6): 947–959. [ Links ]
Holmgren, P.K., Holmgren, N.H. & Barnett, L.C., 1990. "Index Herbariorum". Part I. Regnum Vegetation, 120: 1–693. [ Links ]
Horsley, S.B., R. P.Long, S.W. Bailey, R.A. Hallett & P.M. Wargo, 2002. "Health of eastern North America sugar maple forests and factors affecting decline". Northern J. Appl. For., 19: 34–44. [ Links ]
Hubbell, S.P., 2001. The Unified Theory of Biodiversity and Biogeography. Princeton University Press, Princeton, NJ. [ Links ]
Hutchinson, T.F., R.E. Boerner, L.R. Iverson, S. Sutherland & E.K. Sutherland, 1999. "Landscape patterns of understory composition and richness across moisture and nitrogen mineralization gradient in Ohio (USA) Quercus foresf". Plant Ecology, 144: 177–189. [ Links ]
INEGI, 1970a. Carta edafológica, escala 1: 50 000. Instituto Nacional de Estadística, Geografía e Informática, México, DF. [ Links ]
––––––––––, 1970b. Carta geológica, escala 1: 50 000. Instituto Nacional de Estadística, Geografía e Informática, México, DF. [ Links ]
Iriarte, S.B.B. & R.L. Chazdon, 2005. "Light–dependent seedling survival and growth of four tree species in Costa Rican second–growth rain forests". Journal of Tropical Ecology, 21: 383–395. [ Links ]
Johnston, M.H., 1992. "Soil–vegetation relationships in a Tabonuco forest community in the Luquillo mountains of Puerto Rico". Journal of Tropical Ecology, 8: 253–263. [ Links ]
Kitayama, K., 1992. "An altitudinal transect study of the vegetation of Mount Kinabalú, Borneo". Vegetatio, 102: 149–171. [ Links ]
Kabakoff, R.P. & R.L. Chazdon, 1996. "Effects of canopy species dominance on understory light availability in tropical secondary rain forest". Journal of Tropical Ecology, 12: 779–788. [ Links ]
Kruskal, J.B., 1964. "Nonmetric multidimensional scaling: a numerical method". Psychometrika, 29: 115–129. [ Links ]
Liu, Y. & R.N. Muller, 1993. "Above–ground net primary productivity and nitrogen mineralization in a mixed mesophytic forest of eastern Kentucky". Forest Ecology and Management, 59: 53–62. [ Links ]
Mather, P.M., 1976. Computational methods of multivariate analysis in physical geography. J. Wiley & Sons, London. [ Links ]
McCune, B. & J.B. Grace, 2002. Analysis of ecological communities. MjM Software Design, Gleneden Beach, Oregon. [ Links ]
McCune, B. & M.J. Mefford, 1999. PC–ORD. Multivariate analysis of ecological data: version 4.27. MjM Software, Gleneden Beach, Oregon, US. [ Links ]
Minchin, P.R., 1987. "An evaluation of the relative robustness of techniques for ecological ordination". Vegetatio, 69: 89–107. [ Links ]
Muscolo, A., M. Sidari & R. Mercurio, 2007. "Influence of gap size on organic matter decomposition, microbial biomass and nutrient cycle in Calabrian pine (Pinus laricio, Poiret) stands". Forest Ecology and Management, 242: 412–418. [ Links ]
Oliveira–Filho, A.T., J. A. Ratter & G. J. Shepperd, 1990. "Floristic composition and community structure of a central Brazilian gallery forest". Flora, 184: 103–117. [ Links ]
Oliveira–Filho, A.T., E.A. Vilela, D.A. Carvalho & M.L. Gavilanes, 1994. "Effects of soils and topography on the distribution of tree species in a tropical riverine forest in south–eastern Brazil". J. of Trop. Ecology, 10: 483–508. [ Links ]
Orlóci, L., 1978. Multivariate analysis in vegetation research. 2nd. Ed. Junk, The Hague. [ Links ]
Ortiz–Arrona, C., 1999. Environmental effects on cloud forest tree seedling establishment under a Pinus canopy in Mexico. M.Sc. thesis, Department of Forestry, University of Aberdeen, Aberdeen. [ Links ]
Park, A., 2001. "Environmental influences on post–harvest natural regeneration in Mexican pine–oak forests". Forest Ecology and Management, 144: 213–218. [ Links ]
Pastor, J., J.D. Aber & C.A. McClaugherty, 1982. "Geology, soils and vegetation of Blackhawk Island, Wisconsin". American Midland Naturalist, 108: 266–277. [ Links ]
––––––––––, 1984. "Aboveground production and N and P cycling along a nitrogen mineralization gradient on Blackhawk Island, Wisconsin". Ecology, 65: 256–268. [ Links ]
Peet, R.K., 1980. "Ordination as a tool for analyzing complex data sets". Vegetatio, 42: 171–174. [ Links ]
Peet, R.K.& N.L. Christensen, 1980. "Hardwood forest vegetation of the North Carolina piedmont. Veröff. Geobot". Inst. ETH Stiftung Rübel, 69: 14–39. [ Links ]
Ratter, J.A., 1980. Notes on the vegetation of Fazenda Água Limpa (Brasilia–DF, Brasil). Royal Botanical Garden, Edinburgh. [ Links ]
Rzedowski, J., 1978. La vegetación de México. Editorial Limusa, México, cuarta reimpresión, México, DF. 432 pp. [ Links ]
Schier, G.A. & C.J. McQuattie, 2000. "Effect of manganese on endomycorrhizal sugar maple seedlings". Journal of Plant Nutrition, 23: 1533–1545. [ Links ]
Smith, R.L., 1990. Ecology and Field Biology. Harper & Row Publishers, New York. [ Links ]
Vargas–Rodríguez, Y.L., J.A. Vázquez–García, & G.B. Williamson, 2005. "Environmental Correlates of Tree and Seedling–Sapling Distributions in a Mexican Tropical Dry Forest". Plant Ecology, 80(1): 117–134. [ Links ]
Vázquez–García, J.A., 1995. Gradient analyses of neotropical montane forests. Ph.D. Thesis. University of Wisconsin–Madison. 179 pp. [ Links ]
Vázquez–García, J.A. & T.J. Givnish, 1998. "Altitudinal gradients in tropical forest composition, structure and diversity in the Sierra de Manantlán". J. Ecology, 86: 999–1020. [ Links ]
Vázquez–García, J.A. & Y.L. Vargas–Rodríguez, 1999. "Diversidad de la flora vascular en la región huichola y municipios circunvecinos". Scientia–CUCB A (Universidad de Guadalajara) 1: 7–12. [ Links ]
Vázquez–García, J.A., E. Iracheta R. & Y.L. Vargas–Rodríguez. 1999. "Exploraciones botánicas en la región huichola de Jalisco". Mexicoa, 1(1): 86–89. [ Links ]
Vázquez–García, J.A., M. Cházaro, G. Nieves, Y.L. Vargas–Rodríguez, M. Vázquez & A. Flores, 2004. Flora del norte de Jalisco y etnobotánica huichola. Serie Fronteras de Biodiversidad 1. Universidad de Guadalajara, México. 181 pp. [ Links ]
Vázquez–García, J.A., Y.L. Vargas–Rodríguez & G. Nieves H., 2004. "Diversidad alfa y espectro de la flora vascular del norte de Jalisco y zonas adyacentes", pp 42–52. En: Vázquez–García et al. (eds.). Flora del norte de Jalisco y etnobotánica huichola. Serie Fronteras de Biodiversidad 1. Universidad de Guadalajara, México. 181 pp. [ Links ]
Whittaker, R.H., 1956. "Vegetation of the Great Smokey Mountains". Ecological Monographs, 26: 1–80. [ Links ]
––––––––––, 1960. "Vegetation of the Siskiyou Mountains". Oregon and California. Ecological Monographs, 30: 279–338. [ Links ]
Wills, C., R. Condit, R.B. Foster & S.P. Hubbell, 1997. "Strong density– and diversity–related effects help to maintain tree species diversity in a neotropical forest". Proceedings of the National Academy of Sciences of the United States of America, 94(4): 1252–1257. [ Links ]
Wright, S.J., 2002. "Plant diversity in tropical forests: a review of mechanisms of species coexistence". Oecologia, 130(1): 1–14. [ Links ]
Zak, D.R., K.S. Pregitzer & G.E. Host, 1986. "Landscape variation in nitrogen mineralization and nitrification". Canadian Journal of Forest Research, 16: 1258–1263. [ Links ]

