











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El análisis de las características de un ecosistema forestal como la estructura, la composición y la diversidad de especies a lo largo de un gradiente altitudinal permite comprender los cambios de las comunidades vegetales a diferentes altitudes (Ávila-Sánchez et al., 2018; Gong et al., 2019; Medrano-Meraz et al., 2017), ya que estos pueden ser evidentes en localidades con poca diferencia altitudinal y se prevé que con el cambio climático la distribución y la autoecología de las especies tengan cambios notables (Zacarías-Eslava y del Castillo, 2010).
Las modificaciones en los patrones de distribución de la diversidad de especies a escala regional o global están condicionados a factores como variación climática, latitud, origen geológico del suelo, historia evolutiva, efecto del área (McCain y Grytnes, 2010; Reich et al., 2010), características físicas y químicas del suelo (Sánchez-González y López-Mata, 2005), heterogeneidad de hábitats, complejidad estructural y heterogeneidad topográfica (Lee y Chun, 2016), entre otros.
De manera particular, la diversidad y la estructura son componentes y propiedades importantes que caracterizan a un ecosistema forestal, pues no solo se refieren a la riqueza de especies, sino a una variedad de complejas relaciones de funciones y procesos entre componentes bióticos y abióticos que determinan la heterogeneidad dentro del mismo (Gadow et al., 2012). Por lo tanto, la evaluación y monitoreo de los bosques de pino-encino mediante los índices de diversidad y estructura permiten conocer cómo está estructurada y compuesta la vegetación en diferentes condiciones (Monárrez-González et al., 2020; Ramírez-Santiago et al., 2019; Rendón-Pérez et al., 2021) y contar con elementos básicos para su caracterización y posterior conservación o manejo.
El manejo forestal requiere integrar componentes importantes como el sotobosque, mismo que contribuye a la dinámica de los ciclos biogeoquímicos e influye en el crecimiento y desarrollo de árboles, así como en la abundancia de fauna silvestre, microorganismos e insectos polinizadores (Landuyt et al., 2019); sin embargo, pocos estudios en bosques de pino-encino integran a este componente; de acuerdo con Luna-Bautista et al. (2015); Rendón-Pérez et al. (2021); y Vázquez-Cortez et al. (2018), considerar al sotobosque en estudios de caracterización del bosque de pino-encino, permitirá comprender mejor la dinámica de crecimiento y desarrollo de la vegetación como respuesta a los distintos factores de perturbación natural o antropogénica. Por lo anterior, el presente estudio comprende también al sotobosque.
La estructura, composición y diversidad de la vegetación del bosque de pino-encino de la comunidad de Miguel Hidalgo se desconoce, por lo que este estudio se convierte en una pieza importante para valorar la condición actual de la vegetación y caracterizar adecuadamente al ecosistema a través de un gradiente altitudinal; servirá como herramienta de apoyo para implementar estrategias y tomar decisiones sobre la conservación, manejo y uso de los recursos forestales de la comunidad.
Objetivos
Describir la estructura, composición y diversidad florística actual de las especies arbóreas, arbustivas y herbáceas de un bosque sin manejo forestal de pino-encino, en un gradiente altitudinal entre 1900 m y 2850 m s.n.m., en la comunidad de Miguel Hidalgo, Mpio. de San Antonio Huitepec, del estado de Oaxaca, México.
Materiales y métodos
Área de estudio
El estudio se realizó en la comunidad de Miguel Hidalgo, ubicada en el municipio de San Antonio Huitepec, distrito de Zaachila, Oaxaca (Fig. 1). La comunidad está ubicada dentro del sistema montañoso Sierras Centrales de Oaxaca. Se presenta una pendiente promedio de 40%; un gradiente altitudinal entre 1600 m y 2903 m s.n.m. De acuerdo con la clasificación climática de Köppen, modificada por García (1964), se presentan dos climas: templado subhúmedo y cálido subhúmedo, que dan formación al bosque de pino-encino y bosque de encino (Instituto Nacional de Estadística y Geografía [Inegi], 2010).

La comunidad de Miguel Hidalgo no cuenta ni ha contado con un programa de manejo forestal. En el intervalo altitudinal inferior se presentan afectaciones por plaga de descortezadores e incendios forestales, existe presencia de zonas de producción agrícola (maíz y frijol de temporal; cultivos frutícolas como la manzana, el aguacate y el durazno) y zonas de ganadería extensiva (vacuno, bovino y caprino). Además, la extracción de leña para consumo local es evidente por la cercanía a la zona urbana. En el intervalo altitudinal intermedio, la presencia de cultivos agrícolas y sistemas ganaderos son escasos, sin embargo, se presentan disturbios por incendios forestales de años anteriores. Y en el intervalo altitudinal superior las afectaciones por incendios, agricultura y ganadería son prácticamente nulas.
Diseño de muestreo y medición de variables
El área de estudio comprende una superficie de 1117 ha de bosque de pino-encino en un gradiente altitudinal de 1900 m a 2850 m s.n.m. La intensidad de muestreo fue de 0.2% y se determinaron tres intervalos altitudinales: inferior (1900 m a 2200 m s.n.m.), intermedio (2200 m a 2500 m s.n.m.) y superior (2500 m a 2850 m s.n.m.). De manera sistemática, se distribuyeron 19 sitios en el intervalo altitudinal inferior, 14 en el intervalo altitudinal intermedio y 12 en el intervalo altitudinal superior (Fig. 1), considerando para esto la metodología propuesta por Huerta-Martínez et al., (2014), Lee y Chun (2016) y Sánchez-González y López-Mata (2005).
Se establecieron sitios de muestreo en forma anidada, un sitio principal de 500 m2 para evaluar a las especies arbóreas (Huerta-Martínez et al., 2014), un sitio secundario de 200 m2 para arbustivas y cuatro subsitios de 1 m2 ubicados de manera sistemática a 7 m del centro de cada sitio principal, dirigidas en las cuatro orientaciones (N, S, E, O), para evaluar a las especies herbáceas (Luna-Bautista et al., 2015). Se midieron diámetro y altura total de todos los árboles con diámetro a la altura de 1.30 m mayor o igual a 2.5 cm; además se tomaron dos medidas de la proyección de la copa con orientación norte-sur y este-oeste.
Para las especies arbustivas se registraron la altura total y el diámetro de proyección de la copa en las orientaciones norte-sur y este-oeste. Se consideraron como especies arbustivas todas aquellas que ramificaban desde la base y que presentaron consistencia leñosa. Para las especies herbáceas se registró la altura, la frecuencia y se midió el diámetro (menor y mayor) de la proyección de copa de las diferentes especies, con la finalidad de obtener su cobertura.
Se realizó colecta de ejemplares (acículas/hojas, flor y fruto) en los periodos de diciembre de 2020 a junio de 2021. La identificación de las especies se realizó con base en claves taxonómicas (Calderón de Rzedowski y Rzedowski, 2005; García-Mendoza y Meave, 2011; Guízar, 2015; Rzedowski et al., 2016; Rzedowski y Calderón de Rzedowski, 2008; Zavala, 2003). Adicionalmente, se realizó un cotejo de los ejemplares en el herbario CHAPA, del Colegio de Postgraduados, y la nomenclatura científica se obtuvo mediante la página web Tropicos, del Missouri Botanical Garden (2021).
Análisis de la información
Para conocer la distribución de diámetros y alturas en cada intervalo altitudinal, se generaron histogramas de frecuencia con categorías diamétricas de 5 cm y categorías de altura de 5 m. El índice de valor de importancia relativa (IVI) se calculó de acuerdo con Müeller-Dombois y Ellenberg (1974; ecuación 1) y permitió jerarquizar el peso ecológico de las especies arbóreas, arbustivas y herbáceas en cada intervalo altitudinal.
donde:
DoR = dominancia relativa (área basal y/o cobertura de copa)
DeR = densidad relativa
FR = frecuencia relativa
Para las especies arbóreas, arbustivas y herbáceas, en cada intervalo altitudinal y de acuerdo con Moreno (2001), se calcularon el índice de riqueza de Margalef (ecuación 2) y el índice de diversidad de Simpson (ecuación 3).
donde:
S = número de especies presentes
n = número de individuos de la especie i
N = número total de individuos de todas las especies presentes
Además, con la finalidad de conocer la semejanza florística entre los intervalos altitudinales para las tres formas de vida en conjunto, se utilizó el índice de Sorensen, este índice se calcula con la ecuación 4 (Moreno, 2001) y presenta valores que van de 0 a 100, significando la menor y mayor semejanza florística, respectivamente.
donde:
A = número de especies encontradas en la comunidad A
B = número de especies encontradas en la comunidad B
C = número de especies encontradas en ambas comunidades
Para conocer el patrón de distribución y verificar posibles diferencias estadísticamente significativas en la riqueza, la diversidad y la estructura entre los tres intervalos altitudinales para las tres formas de vida, se realizaron análisis no paramétricos de Kruskal-Wallis seguidos de la prueba post hoc de comparaciones múltiples de la suma de rango de Dunn (P < 0.05), dado que las muestras son desiguales en cada intervalo altitudinal (Dunn, 1964). Previamente, los datos de cada variable se sometieron a la prueba de normalidad (Shapiro-Wilks) y a la prueba de homogeneidad de varianzas (Levene). Los análisis estadísticos, así como el cálculo de la diversidad y riqueza se ejecutaron en el programa SAS (SAS Institute, 2008) y PAST versión 4.03 (Hammer et al., 2001).
Resultados
Composición, diversidad y riqueza
Se evaluaron 3123 árboles, 1973 arbustos y 1643 hierbas, pertenecientes a 125 especies distribuidas en 84 géneros y 40 familias. Se registraron 27 especies arbóreas, 25 arbustivas y 73 herbáceas. Asteraceae y Fabaceae presentaron mayor número de especies y Ericaceae, Pinaceae y Fagaceae fueron las familias mejor representadas (Fig. 2)
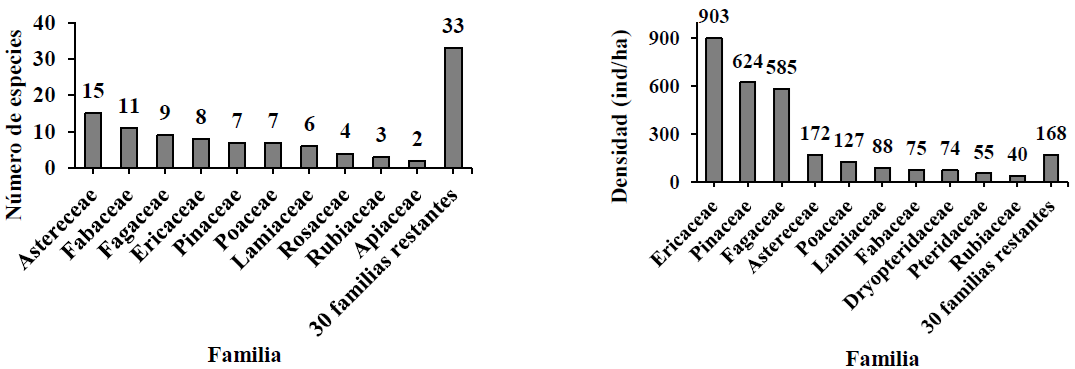
Figura 2 Número de especies y densidad de las familias más representativas en un gradiente altitudinal del bosque de pino-encino en Miguel Hidalgo, Oaxaca.
Se presentó mayor número de especies herbáceas respecto a las arbóreas y arbustivas; sin embargo, la abundancia total de especies fue mayor en el componente arbóreo (Tabla 1).
Tabla 1 Número de especies, abundancia total, índices de riqueza y diversidad de especies arbóreas, arbustivas y herbáceas por intervalo altitudinal del bosque de pino-encino en Miguel Hidalgo, Oaxaca.
| Intervalo altitudinal | Forma de vida | Número de especies | Abundancia total | Índice de Simpson | Índice de Margalef |
| Inferior | Arbórea | 21 ac | 1303 a | 0.64 a | 1.10 a |
| Arbustiva | 14 b | 800 b | 0.27 b | 0.55 bcd | |
| Herbácea | 47 cde | 665 c | 0.62 a | 1.63 ace | |
| Intermedio | Arbórea | 14 ad | 1195 a | 0.57 a | 0.90 ab |
| Arbustiva | 13 b | 907 b | 0.16 b | 0.29 d | |
| Herbácea | 37 cde | 390 c | 0.62 a | 1.71 ace | |
| Superior | Arbórea | 17 ae | 625 a | 0.55 a | 0.93 ac |
| Arbustiva | 16 b | 266 b | 0.26 b | 0.46 bcd | |
| Herbácea | 42 c | 588 c | 0.66 a | 1.94 e |
*Medias seguida por letras iguales indican que no hay diferencias significativas (Suma de rango de Dunn, P < 0.05).
El índice de diversidad de Simpson presentó valores promedio de 0.59 para arbóreas, 0.23 para arbustivas y 0.63 para herbáceas. El índice de Margalef presentó valores promedio de 0.98, 0.43 y 1.76, respectivamente, para las especies arbóreas, arbustivas y herbáceas.
Las comparaciones de medias de la suma de rango de Dunn no mostraron diferencias significativas en diversidad y riqueza para cada forma de vida entre intervalos altitudinales (Tabla 1). Sin embargo, el índice de Simpson de las especies arbustivas fue menor en los tres intervalos altitudinales respecto a las especies arbóreas y herbáceas. Además, los intervalos altitudinales inferior e intermedio presentaron el menor índice de Margalef para las especies arbustivas; y el intervalo altitudinal superior presentó la mayor riqueza de especies herbáceas.
La similitud de especies para arbóreas, arbustivas y herbáceas en conjunto entre el intervalo altitudinal intermedio y superior fue del 65%; el intervalo altitudinal inferior e intermedio presentó una similitud de 63%, y la menor similitud de especies lo presentó el intervalo altitudinal inferior y superior con 51%.
Distribución horizontal y vertical
Las distribuciones diamétricas en el intervalo altitudinal inferior e intermedio presentaron un patrón de J invertida y el intervalo altitudinal superior presentó un patrón unimodal, concentrando menor densidad de individuos en categorías de menor diámetro respecto al intervalo altitudinal inferior e intermedio (Fig. 3). Asimismo, las distribuciones por categorías de altura concentran una mayor densidad de individuos en las categorías de altura de 5 m y 10 m en los tres intervalos altitudinales (Fig. 4).
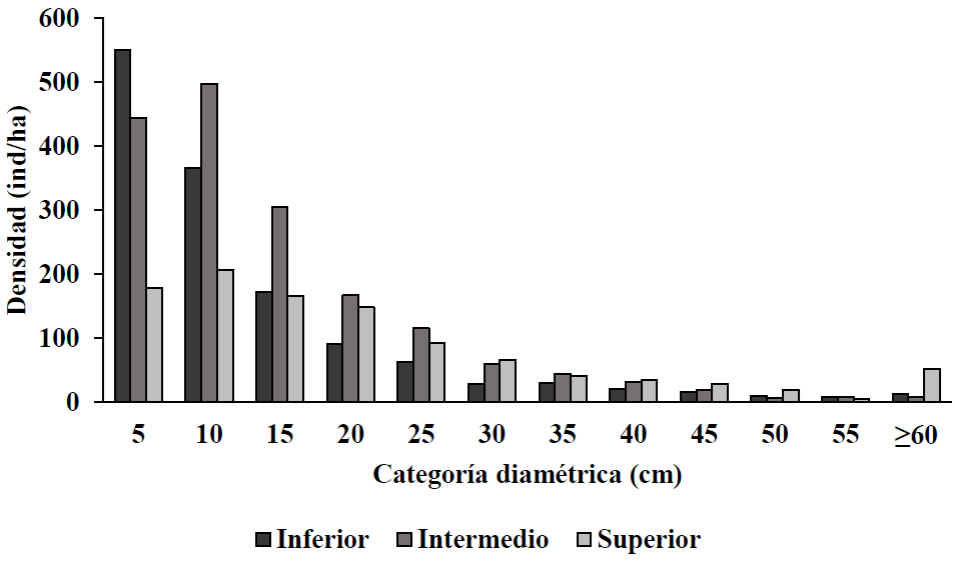
Figura 3 Distribución por categoría diamétrica de individuos ≥ 2.5 cm de diámetro normal, por intervalo altitudinal del bosque de pino-encino en Miguel Hidalgo, Oaxaca.
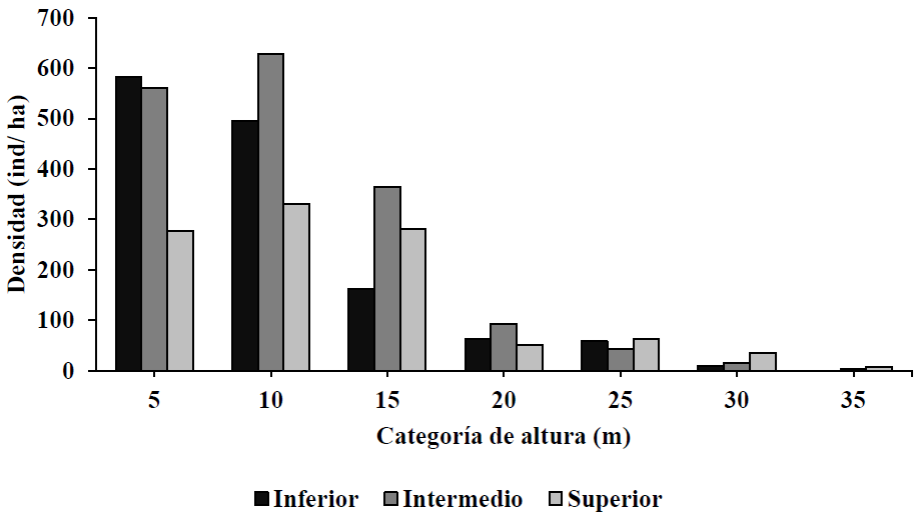
Figura 4 Distribución por categoría de alturas de individuos ≥ 2.5 cm de diámetro normal, por intervalo altitudinal del bosque de pino-encino en Miguel Hidalgo, Oaxaca.
El intervalo altitudinal superior presentó menor densidad de árboles respecto al intervalo altitudinal intermedio; asimismo, el intervalo altitudinal superior presentó mayores valores promedio de diámetro normal, altura y de cobertura de copa individual, valores que fueron diferentes (P < 0.05) a los intervalos altitudinales inferior e intermedio (Tabla 2). Así mismo, el intervalo altitudinal inferior presentó menor área basal respecto a los intervalos altitudinales intermedio y superior.
Tabla 2 Valores estructurales de las especies arbóreas, arbustivas y herbáceas, por intervalo altitudinal del bosque de pino-encino en Miguel Hidalgo, Oaxaca.
| Variables estructurales | Intervalo altitudinal | ||
| Inferior | Intermedio | Superior | |
| Arbórea | |||
| Densidad (ind/ha) | 1372 ab | 1707 a | 1042 b |
| Diámetro (cm) | 12.9 a | 14.6 b | 20.8 b |
| Área basal (m2/ha) | 31.4 a | 42.7 b | 55.5 b |
| Altura (m) | 9.5 a | 10.4 b | 12.2 c |
| Cobertura de copa individual (m2) | 10.5 a | 9.9 a | 16.7 b |
| Arbustiva | |||
| Densidad (ind/ha) | 2105 a | 3239 a | 1108 a |
| Altura (m) | 1.6 a | 1.5 b | 3.3 c |
| Cobertura de copa individual (m2) | 1.3 a | 1.0 b | 2.3 c |
| Herbácea | |||
| Densidad (ind/ha) | 8750 ab | 6964 a | 12 250 b |
| Altura (cm) | 23 a | 24.8 a | 40.9 c |
| Cobertura individual (cm2) | 302 a | 454.5 b | 362.6 b |
*Medias seguida por letras iguales indican que no hay diferencias significativas (Suma de rango de Dunn, P < 0.05). Las diferencias significativas solo aplican por forma de vida.
Por otro lado, las variables estructurales de las especies arbustivas y herbáceas indican que la altura y la cobertura de copa individual fueron mayor en el intervalo altitudinal superior, respecto al intervalo altitudinal inferior e intermedio (Tabla 2).
Índice de valor de importancia
Pinus oocarpa Schiede. presentó los valores más altos del IVI para los intervalos altitudinales inferior e intermedio y en el intervalo altitudinal superior Pinus pseudostrobus Lindl. presentó el valor más alto (Tabla 3). Pinus y Quercus representaron 44% y 38% respectivamente, mismas que suman un total de 82% del IVI para el total del área de estudio.
Tabla 3 Índice de valor de importancia (IVI) de las especies arbóreas por intervalo altitudinal del bosque de pino-encino en Miguel Hidalgo, Oaxaca.
| Intervalo altitudinal inferior | Intervalo altitudinal intermedio | Intervalo altitudinal superior | |||
| Especie | IVI (%) | Especie | IVI (%) | Especie | IVI (%) |
| Pinus oocarpa | 26.6 | Pinus oocarpa | 22.9 | Pinus pseudostrobus | 23.3 |
| Quercus scytophylla | 14.4 | Quercus scytophylla | 22.6 | Quercus rugosa | 14.3 |
| Pinus pseudostrobus | 12.5 | Arbutus xalapensis | 12.2 | Pinus leiophylla | 10.7 |
| Quercus elliptica | 8.8 | Pinus leiophylla | 11.2 | Arbutus xalapensis | 10.5 |
| Arbutus xalapensis | 6.4 | Quercus elliptica | 10 | Quercus obtusata | 9.1 |
| Quercus acutifolia | 6.2 | Pinus pseudostrobus var. apulcensis | 5.9 | Alnus acuminata subsp. arguta | 7.2 |
| Bejaria aestuans | 4.4 | Quercus obtusata | 4.5 | Pinus pseudostrobus var. apulcensis | 6.9 |
| Quercus magnoliifolia | 4.2 | Quercus rugosa | 3.8 | Quercus crassifolia | 3.8 |
| Pinus leiophylla | 3.4 | Pinus pseudostrobus | 2.8 | Quercus elliptica | 3.2 |
| Pinus teocote | 2.6 | Quercus acutifolia | 1.2 | Quercus scytophylla | 2.8 |
| Quercus obtusata | 2.5 | Quercus salicifolia | 1.1 | Pinus oocarpa | 2.4 |
| Pinus pseudostrobus var. apulcensis | 1.8 | Alnus acuminata subsp. arguta | 0.8 | Prunus serotina | 1.5 |
| Pinus devoniana | 1.3 | Buddleja cordata | 0.6 | Quercus acutifolia | 1.3 |
| Quercus glaucescens | 1 | Quercus glaucescens | 0.6 | Pinus patula | 0.9 |
| Juniperus poblana | 0.8 | TOTAL | 100 | Clethra mexicana | 0.7 |
| Diphysa americana | 0.8 | Buddleja cordata | 0.7 | ||
| Quercus salicifolia | 0.7 | Quercus salicifolia | 0.7 | ||
| Oreopanax xalapensis | 0.4 | TOTAL | 100 | ||
| Cornus disciflora | 0.4 | ||||
| Prunus serotina | 0.4 | ||||
| Vachellia pennatula | 0.4 | ||||
| TOTAL | 100 |
Del componente arbustivo y herbáceo, Gaultheria erecta Vent. y Pteridium aquilinum var. feei (W. Schaffn. ex Fée) Maxon presentaron el valor más alto de IVI en el intervalo altitudinal inferior e intermedio, y Salvia cinnabarina M. Martens & Galeotti en el intervalo altitudinal superior (Tablas 4 y 5).
Tabla 4 Índice de valor de importancia (IVI) de las especies arbustivas por intervalo altitudinal del bosque de pino-encino en Miguel Hidalgo, Oaxaca.
| Intervalo altitudinal inferior | Intervalo altitudinal intermedio | Intervalo altitudinal superior | |||
| Especie | IVI (%) | Especie | IVI (%) | Especie | IVI (%) |
| Gaultheria erecta | 51.1 | Gaultheria erecta | 58.6 | Gaultheria erecta | 46.4 |
| Lyonia squamulosa | 19.4 | Lyonia squamulosa | 29.9 | Baccharis heterophylla | 20.6 |
| Agarista mexicana | 11.5 | Baccharis heterophylla | 5.3 | ARB3 | 16.5 |
| ARB1 | 7.3 | Agarista mexicana | 4.0 | Comarostaphylis glaucescens | 5.3 |
| Baccharis heterophylla | 6.6 | Holodiscus argenteus | 0.6 | Lyonia squamulosa | 2.6 |
| *Otras especies | 4.1 | *Otras especies | 1.5 | * Otras especies | 8.6 |
| TOTAL | 100 | TOTAL | 100 | TOTAL | 100 |
Tabla 5 Índice de valor de importancia (IVI) de las especies herbáceas por intervalo altitudinal del bosque de pino-encino en Miguel Hidalgo, Oaxaca.
| Intervalo altitudinal inferior | Intervalo altitudinal intermedio | Intervalo altitudinal superior | |||
| Especie | IVI (%) | Especie | IVI (%) | Especie | IVI (%) |
| Pteridium aquilinum var. feei | 14.6 | Pteridium aquilinum var. feei | 20.8 | Salvia cinnabarina | 13.6 |
| Elaphoglossum latifolium | 14.4 | Adiantum andicola | 11.6 | Bromus dolichocarpus | 10.3 |
| Muhlenbergia emersleyi | 12.8 | Muhlenbergia emersleyi | 8.0 | Desmodium incanum | 9.8 |
| Chimaphila maculata | 7.1 | Elaphoglossum latifolium | 7.9 | Adiantum andicola | 7.4 |
| Melinis minutiflora | 6.8 | Bromus dolichocarpus | 5.1 | Alloispermum scabrum | 7.3 |
| Adiantum andicola | 3.4 | Salvia cinnabarina | 4.6 | Iresine diffusa | 6.3 |
| Cosmos crithmifolius | 3.4 | Chimaphila maculata | 4.4 | Piptothrix areolaris | 6.2 |
| Galium mexicanum subsp. Asperrimum | 2.9 | Stachys coccinea | 4.1 | Rumfordia floribunda | 5.8 |
| Bromus dolichocarpus | 2.8 | Eryngium monocephalum | 3.8 | Phenax hirtus | 5.0 |
| Desmodium incanum | 2.8 | Bidens pilosa | 3.5 | Crusea coccinea | 3.5 |
| *Otras especies | 29.0 | *Otras especies | 26.1 | *Otras especies | 24.8 |
| TOTAL | 100 | TOTAL | 100 | TOTAL | 100 |
Discusión
Composición, diversidad y riqueza
El número de especies arbóreas encontradas en el presente estudio está dentro del intervalo de 5 a 48 especies registradas en bosques de pino-encino sin manejo forestal (Méndez-Osorio et al., 2018; Ramírez-Santiago et al., 2019; Rendón-Pérez et al., 2021; Silva-García et al., 2021), y el número de especies arbustivas y herbáceas en conjunto es superior al intervalo de 12 a 68 especies encontradas para el mismo tipo de vegetación (Rendón-Pérez et al., 2021; Vázquez-Cortez et al., 2018; Zacarías-Eslava y del Castillo, 2010).
Se presentó un índice de Simpson para especies arbóreas catalogado por Moreno (2001) como diversidad media, superior a lo obtenido por Medrano-Meraz et al. (2017) en Durango y Manzanilla-Quijada et al. (2020) en Nuevo León; pero inferior a lo registrado por Luna-Bautista et al. (2015) y Vázquez-Cortez et al. (2018) en Oaxaca. Se encontró una diversidad baja para especies arbustivas y media para herbáceas, estos valores son inferiores a los encontrados por Luna-Bautista et al. (2015) en la Sierra Norte de Oaxaca. Sin embargo, la diversidad del sotobosque fue superior a lo encontrado por Rendón-Pérez et al. (2021) en Hidalgo.
Se registraron índices de Margalef inferiores a dos para las tres formas de vida, lo que se considera por Moreno (2001) como riqueza baja. Algunos estudios en bosques sin manejo forestal que han considerado especies arbóreas (López-Hernández et al., 2017; Manzanilla-Quijada et al., 2020; Méndez-Osorio et al., 2018; Vázquez-Cortez et al., 2018), o las tres formas de vida (Rendón-Pérez et al., 2021), han encontrado una riqueza similar. Sin embargo, Silva-García et al. (2021), en Durango, hallaron una riqueza media para especies arbóreas.
En bosques templados de pino-encino es común encontrar baja diversidad y riqueza de especies (Ávila-Sánchez et al., 2018), esto puede ser explicado en parte por la alta frecuencia y dominancia de los géneros Pinus y Quercus (Méndez-Osorio et al., 2018). Alfaro-Reyna et al. (2019); González-Tagle et al. (2008); y López-Hernández et al. (2017), los requerimientos de luz, la alteración natural y antropogénica -como los incendios forestales y las extracciones a pequeña escala, pero constantes- moldean la estructura y composición de especies de los bosques templados sin manejo, aumentando considerablemente la dominancia de estos dos géneros; lo anterior concuerda con lo observado en el presente estudio a través del gradiente altitudinal.
Además, en el gradiente altitudinal del presente estudio, los índices de riqueza y diversidad no presentaron cambios significativos. Sin embargo, como lo demuestra el índice de Sorensen, se presentó un cambio de especies de 49% entre el intervalo altitudinal inferior y el superior. Así mismo, el número de especies y la abundancia total sí presentaron variación entre intervalos altitudinales entre formas de vida, posiblemente debido a cambios en las condiciones bióticas y abióticas a lo largo del gradiente altitudinal.
Se ha observado que el tipo de patrón de distribución de la diversidad y riqueza de especies puede cambiar por formas de vida, ser similares o no mostrar un patrón definido (Gong et al., 2019; Lee y Chun, 2016). Diferentes factores contribuirán en la distribución de la diversidad y riqueza de especies (Reich et al., 2010) y su interacción con los diferentes procesos que existen en los ecosistemas forestales (Lee y Chun, 2016; McCain y Grytnes, 2010). Sin embargo, un patrón de distribución claramente definido pudiese estar relacionado con los diferentes tipos de vegetación que se contemplan a lo largo de un gradiente altitudinal (Ávila-Sánchez et al., 2018; Martin et al., 2021; Sánchez-González y López-Mata, 2005), similar a lo observado en el presente estudio.
Distribución horizontal y vertical
La disminución del número de individuos al incrementar el diámetro sugiere la existencia de masas irregulares en etapa de regeneración y crecimiento, como lo encontrado para bosques templados sin manejo por Manzanilla-Quijada et al. (2020) en Nuevo León, Ramírez-Santiago et al. (2019) en Oaxaca y Méndez-Osorio et al. (2018) en Guerrero. Así mismo, el comportamiento de la distribución en altura con respecto a la distribución diamétrica es muy similar, pues ambas variables están correlacionadas (Smith et al., 1997).
En bosques con alta densidad, como el que se presentó en el intervalo altitudinal intermedio, la eficiencia en el crecimiento estará limitada a la disponibilidad de espacio, agua, luz y nutrientes debido a una alta tasa de competencia (Binkley et al., 2013), generando mayor susceptibilidad del bosque al ataque de plagas, enfermedades o mayor riesgo de ocurrencia de incendios forestales; dichas perturbaciones fueron observadas en los intervalos altitudinales inferior e intermedio. Estos factores aceleran en mayor grado la dinámica de regeneración de las diferentes especies en los bosques templados mixtos (Alfaro-Reyna et al., 2019; Cortés-Montaño et al., 2012).
Un mayor crecimiento en altura, diámetro y cobertura de copa individual en especies arbóreas y una mayor altura en especies arbustivas y herbáceas en el intervalo altitudinal superior sugieren rodales en mejor estado de conservación y, probablemente, una mayor productividad forestal. Está comprobado que una alta productividad del rodal influye en un mejor crecimiento de los árboles en altitudes superiores en algunas regiones montañosas (Guerrero-Hernández et al., 2019; Zacarías-Eslava y del Castillo, 2010), apoyado en gran medida por una menor perturbación agrícola y ganadera debido a la elevada pendiente y a la lejanía de la zona urbana (Bonilla-Moheno et al., 2012; Pérez-Hernández et al., 2021), eso mismo se observó en este estudio.
Índice de valor de importancia
Está documentado que el manejo forestal modifica la estructura y composición de especies del bosque de pino-encino, incrementando el peso ecológico de especies de alto valor comercial como el género Pinus (López-Hernández et al., 2017; Monárrez-González et al., 2020; Rendón-Pérez et al., 2021). En el área de estudio no existe actualmente un manejo forestal; sin embargo, se presenta un nivel de perturbación antropogénica de acuerdo con cada intervalo altitudinal. Se observa que el género Pinus mantiene una mezcla proporcional con el género Quercus en porcentajes similares a lo encontrado por Monárrez-González et al. (2020) en Durango y Rendón-Pérez et al. (2021) en Hidalgo. No obstante, Méndez-Osorio et al. (2018) en Guerrero y Silva-García et al. (2021) en Durango, encontraron valores superiores del género Pinus en bosques de pino-encino sin manejo forestal. La diferencia en los pesos ecológicos en los bosques sin manejo forestal a través de un gradiente altitudinal está condicionada a factores climáticos, topográficos y a las alteraciones antropogénicas o naturales propias de cada región como lo documentan Bonilla-Moheno et al. (2012), Pérez-Hernández et al. (2021) y Salas y Williams (2019).
El comportamiento de la dominancia de las diferentes especies a través del gradiente altitudinal sugiere un cambio en las condiciones bióticas y abióticas pues en el intervalo altitudinal superior se presentaron especies como Quercus crassifolia Bonpl., Pinus patula Schiede ex Schltdl. & Cham. y Pinus pseudostrobus, estas especies podrían considerarse indicadoras de mayor humedad; y en el intervalo altitudinal inferior se encontraron especies como Pinus devoniana Lindl., Pinus teocote Schiede ex Schltdl. & Cham. y Pinus oocarpa, mismas que se distribuyen en condiciones de menor humedad (Martin et al., 2021; Ríos-Altamirano et al., 2016).
En especies del sotobosque de bosques templados, la dinámica de distribución, crecimiento y desarrollo pareciera ser más compleja, pues se ha encontrado que están sujetos en gran medida a la composición y estructura del rodal, a las características físicas y químicas del suelo, a los microclimas, a la microtopografía, a la capa de hojarasca y a la disponibilidad de luz (Barbier et al., 2008; Sercu et al., 2017; Tinya y Ódor, 2016). Además, como se observó en el presente estudio, los diferentes niveles de perturbación antropogénica en cada gradiente altitudinal también son un factor que modifica el peso ecológico de las especies.
Conclusiones
El bosque de pino-encino sin manejo forestal presentó una estructura característica de un bosque irregular; con altas densidades en los intervalos altitudinales intermedio e inferior, influenciado probablemente por una elevada tasa de perturbación debido a los incendios forestales y la plaga de descortezadores presentes en la zona. Así mismo, en el intervalo altitudinal superior se presentaron rodales en mejor estado de conservación, característico de un bosque maduro y probablemente con una mayor productividad forestal que en altitudes inferiores.
No se presentaron cambios significativos en diversidad y riqueza de las especies arbóreas, arbustivas y herbáceas a través del gradiente altitudinal, sin embargo, se presentó un cambio en el número de especies y su abundancia total entre formas de vida; por lo que, tal vez integrando variables climáticas se podría complementar la comprensión de la dinámica de distribución y crecimiento a través de este gradiente altitudinal del bosque de pino-encino.
El IVI presentó una similitud con otros bosques templados sin manejo forestal; sin embargo, a través del gradiente altitudinal se presentó un cambio en los pesos ecológicos de las especies arbóreas, arbustivas y herbáceas, misma que está condicionada a los factores climáticos, topográficos y a las alteraciones antropogénicas o naturales propias de cada región.

