











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
El estudio de la vegetación ribereña permite comprender sus interacciones con la dinámica hidrológica y geomor-fológica a lo largo de la red fluvial (Naiman et al., 2005); así como las decisiones y prácticas de los diferentes actores sociales en el territorio (Flores-Díaz et al., 2014). Wohl et al. (2021) describen a las zonas fluviales como altamente heterogéneas y con patrones de conectividad asociada a la historia regional, resultando en sistemas resilientes en constante reconfiguración. Alrededor del mundo las riberas han sido profundamente transformadas (Vázquez et al., 2015; Hughes y Vadas, 2021) y, dada la importancia que tienen para los ecosistemas dulceacuícolas y para la sociedad, resulta muy relevante generar conocimiento que permita guiar su recuperación (Naiman et al., 2002; Naiman et al., 2005; Tickner et al., 2020). Los ríos y sus riberas se encuentran bajo presión por factores como el uso del espacio ribereño para la producción agropecuaria, la construcción de viviendas, el acceso al agua, la construcción de caminos y la regulación del caudal (Dudgeon et al., 2006; Flores-Díaz, 2014; Postel y Richter, 2003; Singh et al., 2021). La presencia de árboles, arbustos y restos leñosos en el canal y en la ribera ha mostrado ofrecer beneficios como el incremento de la diversidad de hábitats y especies, además de la provisión de espacios más frescos en comparación con tramos desprovistos de vegetación (Garner et al., 2014; Bowler et al., 2012).
La vegetación ribereña está fuertemente influenciada por la variación estacional del caudal (Moore y Richardson, 2012; Naiman y Décamps, 1997; Poole, 2002), resultando en patrones locales que la distinguen de la vegetación adyacente (Naiman y Décamps, 1997). Las plantas propias de la ribera tienen características que favorecen la estabilización de los bancos del río, toleran las crecidas y los cambios en los patrones de flujo (Gurnell et al., 2012; Gurnell y Grabowski, 2016), e indican actividad hidrológica (Naiman, et al., 2005).
Estudios sobre la vegetación ribereña describen su asociación con rasgos geográficos como la elevación (Camacho-Rico et al., 2006); su relación con la tempo-ralidad de los arroyos a los que está asociada, los patrones de flujo de la red hídrica (Aguiar y Ferreira, 2005); los impactos derivados de la regulación del caudal (Postel y Richter, 2003); así como la importancia del conocimiento local sobre este sistema (Flores-Díaz, 2014). En diversos paisajes transformados, la vegetación ribereña constituye los únicos fragmentos de bosque, o bien de árboles aislados, que sostienen funciones críticas para la biodiversidad y los ecosistemas (Giraldo et al., 2022), ya que los árboles remanentes participan en procesos como la provisión de semillas y el mantenimiento de la conectividad (Von Thaden et al., 2022). La pérdida de bosques ribereños asociada a los cambios en el uso del suelo y al manejo del agua, cambia la dinámica de los ríos (Luque y González-Trujillo, 2021) y favorece la presencia de especies invasoras (Tolkkinen et al., 2020) y la capacidad de retención de sedimentos (Argueta Navarrete, 2019) por los cambios en el patrón de inundaciones.
La red hidrológica que cruza el territorio de ejidos y comunidades indígenas, ubicados en la zona de amortiguamiento de la Reserva de la Biósfera Mariposa Monarca (RBMM), cuyo programa de manejo carece de lineamientos para los ríos y riberas fluviales (Secretaría de Medio Ambiente y Recursos Naturales [Semarnat], 2001), ha mantenido procesos de cambio de uso del suelo de bosques a cultivos como la zarzamora y el aguacate (Hernández, 2018). Esta elevada demanda de agua para uso doméstico y agrícola ha dado pie a la existencia de múltiples formas de suministro como mangueras y canales de riego, siendo desconocido el impacto de estas prácticas sobre los ríos. Por lo anterior, esta zona representa una oportunidad para el estudio de la vegetación remanente y su relación con los patrones de los ríos y escurrimientos y, con ello, generar conocimiento situado, no solo por la unicidad de cada tramo fluvial (Gurnell y Grabowski, 2016; Hough-Snee, 2016; Moore y Richardson, 2003), sino por la importancia que tiene generar lineamientos acordes con el contexto local.
Objetivos
Caracterizar la estructura y composición de la vegetación remanente, su relación con la elevación, la precipitación, el orden hidrológico y la temporalidad del flujo de agua; así como la diversidad de especies principalmente arbóreas, y distinguiendo a aquellas que son potencialmente invasoras. Este trabajo busca contribuir a la conservación dulceacuícola en una zona importante para la biodiversidad global, que es la Reserva de la Biósfera de la Mariposa Monarca (RBMM), en México.
Materiales y métodos
La zona de estudio es una franja montañosa mexicana ubicada al noreste de la ciudad de Zitácuaro, Michoacán (Fig. 1); cabecera del río San Juan Zitácuaro, y forma parte de la cuenca del Río Balsas. Se encuentran inmersas en esta área las comunidades indígenas de Francisco Serrato, Car-pinteros y Curungueo.
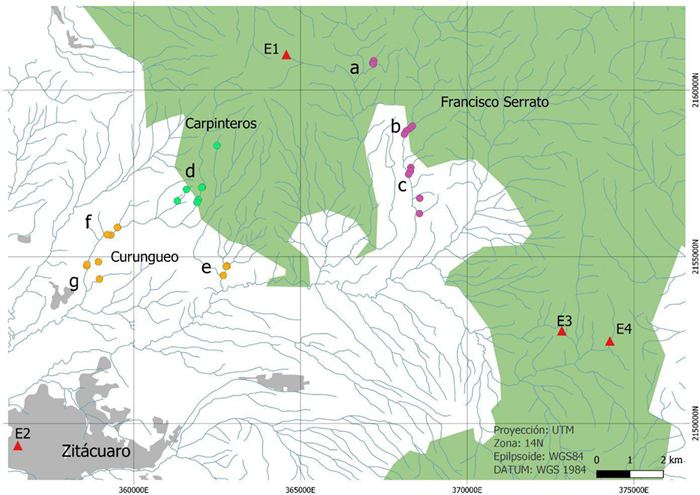
FIGURA 1 Ubicación de las redes (a - g) y sitios (círculos) en Francisco Serrato, Curungueo y Carpinteros.
La red hídrica está representada con la línea azul. Los triángulos indican estaciones meteorológicas (E1: Monarca II, E2: Zitácuaro, E3: Lindero, E4: Cuesta del Carmen). El polígono verde indica la zona de amortiguamiento de la RBMM, la Ciudad de Zitácuaro en gris.
Caracterización de los sitios. Los conjuntos vectoriales del Instituto Nacional de Estadística y Geografía [Inegi] (2010), escala 1:50 000 permitieron identificar los arroyos en la cabecera de la cuenca (de la red hidrológica) y asignarles orden hidrológico de1er a 3er orden, por ser cabecera de cuenca y de acuerdo con los criterios de Strahler (1957); además se consideró el factor de la temporalidad. En campo se registró la altitud y, para la definición de uso del suelo y vegetación, se utilizaron las categorías propuestas por Mas et al. (2017). La precipitación media anual fue interpolada utilizando la información de las 15 estaciones climáticas descritas en Hernández (2018) y analizadas en el sistema de información geográfica QGis (2020) con la herramienta Thin plate spline (Donato y Belongie, 2002). Los tramos de arroyos analizados fueron agrupados en función de la red hidrológica a la que pertenecieran y del ejido o comunidad en que estuvieran ubicados: Carpinteros y Curungueo, y los parajes La Junta, Xorejé y Tamejé de Francisco Serrato (Fig. 1).
Muestreo de vegetación de la ribera. En la vegetación ribereña arbórea remanente se realizó un muestreo siste-mático siguiendo los criterios de Sánchez-Velásquez & Pineda (2000) con transectos de 100 m paralelos al río por 3 m de ancho. En cada sitio se midieron con una cinta métrica solo los árboles con perímetro a la altura del pecho ≥ 10 cm; las alturas individuales fueron estimadas con un medidor láser DIS - 62 (±1 mm). Las morfoespecies fueron colectadas y enviadas al herbario del Instituto de Investigaciones en Ecosistemas y Sustentabilidad (IIES - UNAM) para su herborización, la determinación se hizo mediante la consulta de bases de datos florísticos como la de Jstor (s.f.). En sitios dominados por alguna especie arbustiva o herbácea, esta se determinó y se analizó su valor de importancia (descrito en la siguiente sección) por considerarse potencialmente invasoras (Richardson y Pyšek, 2004); entendidas como especies ajenas a la ribera y cuya presencia es dominante en un sitio.
Análisis. El valor de importancia (VI) de cada especie (área basal, altura y densidad de individuos) (Matteucci y Colma, 1982) fue estimado para cada sitio (n = 30) y para la red hidrológica (n = 7). Se realizó un análisis de componentes principales (ACP) para sintetizar las variables anteriormente señaladas y un agrupamiento jerárquico para evidenciar los posibles rasgos fisonómicos compartidos (descritos en la sección correspondiente), utilizando las funciones princomp y hclust de los programas R v.4.0.3 (R Core Team, 2020) y Rstudio v.1.1.1093 (RStudio Team, 2021).
La diversidad alfa fue calculada (Magurran, 2004) con Biodiversity Pro ® (McAleece et al., 1997), a partir del índice de Shannon (ecuación 1).
donde:
pi: proporción de individuos de la i-ésima especie
El valor de diversidad de las redes fue comparado con un análisis de rarefacción para S con Past®. El recambio de especies entre sitios (diversidad beta) se estimó con el índice de Simpson (ecuación 2).
donde:
A: cantidad de especies compartidas entre dos muestras B y C: número de especies registradas en un sitio
min: valor mínimo de especies restringidas en las muestras comparadas (B o C)
Este índice permite evidenciar las diferencias en la composición de especies entre sitios (Koleff et al., 2003; Koleff, 2005). La diversidad beta también fue estimada con el índice de Sørensen cuantitativo (SSOR), usando EstimateS (Colwell, 2005), con base en la ecuación 3, de Moreno (2001).
donde:
aN y bN: número total de individuos en los sitios A y B, respectivamente
pN: sumatoria de la densidad más baja registrada de especies compartidas
Se analizaron las relaciones entre la cubierta del suelo, la precipitación y el orden hidrológico, con los índices de diversidad y la presencia de especies potencialmente invasoras, utilizando modelos lineales generales y generalizados, con las funciones glm y lm del software R v.4.0.3 (R Core Team, 2020).
Resultados
Características del conjunto de tramos analizados. Los sitios analizados están ubicados en una zona donde la precipitación media anual (1020 mm - 1175 mm) varía de acuerdo con la elevación (1920 m - 2867 m) (n = 30, R2 = 0.83). En las partes más elevadas y lluviosas se encuentran bosques de pino-encino y oyamel, y hacia las medias se incrementan la agricultura de temporal y los cultivos perennes (Tabla 1). Los arroyos permanentes (n = 14) son de orden hidrológico más alto (X2 = 15.787, df = 2, p < 0.01); mientras los arroyos de orden menor son en su mayoría intermitentes y están en zonas más elevadas que los perennes.
TABLA 1 Características geográficas, riqueza e índice de Shannon-Weiner de los sitios analizados de la cuenca San Juan Zitácuaro.
| Comunidad indígena | Red | Sitio | OH* | Tempo- ralidad** | Cubierta del suelo*** | Elevación (m s.n.m.) | Precipitación anual (mm) | Familias | Géneros | Especies | Shannon H' |
| a: La Junta | Junta2 | 1 | I | BPE/O | 2867 | 1175 | 8 | 10 | 10 | 0.72 | |
| Junta3 | 1 | I | BPE/O | 2837 | 1174 | 6 | 7 | 7 | 0.54 | ||
| Junta1 | 1 | I | BPE/O | 2780 | 1174 | 4 | 6 | 7 | 0.67 | ||
| Xoreje1 | 3 | P | BPE | 2715 | 1129 | 5 | 6 | 7 | 0.59 | ||
| b: | Xoreje2 | 3 | P | BPE | 2656 | 1128 | 5 | 5 | 5 | 0.6 | |
| Francisco | Xorejé | Xoreje3 | 3 | P | BPE | 2640 | 1130 | 4 | 4 | 4 | 0.53 |
| Serrato | Xoreje4 | 3 | P | BPE | 2619 | 1129 | 3 | 3 | 3 | 0.47 | |
| Tam3 | 1 | I | AT | 2510 | 1108 | 4 | 5 | 5 | 0.64 | ||
| c: Tamejé | Tam5 | 1 | I | AT | 2497 | 1105 | 4 | 4 | 4 | 0.52 | |
| Tam4 | 1 | I | AT | 2488 | 1106 | 2 | 2 | 2 | 0.3 | ||
| Tam2 | 1 | I | AT | 2470 | 1078 | 5 | 6 | 7 | 0.71 | ||
| Tam1 | 1 | I | AT | 2051 | 1087 | 5 | 5 | 5 | 0.64 | ||
| Carp1 | 1 | I | BPE | 2587 | 1129 | 7 | 9 | 11 | 0.86 | ||
| Carp5 | 1 | I | BPE | 2333 | 1098 | 11 | 11 | 12 | 0.86 | ||
| d: Carpin | Carp4 | 1 | I | BPE | 2179 | 1089 | 9 | 9 | 11 | 0.97 | |
| Carpinteros | Carp2 | 2 | P | BPE | 2451 | 1123 | 8 | 11 | 12 | 0.96 | |
| Carp3 | 2 | P | BPE | 2365 | 1122 | 5 | 7 | 7 | 0.67 | ||
| Carp6 | 2 | P | BPE | 2265 | 1115 | 6 | 7 | 7 | 0.79 | ||
| Carp7 | 2 | P | BPE | 2262 | 1114 | 2 | 2 | 2 | 0.2 | ||
| Curu2 | 2 | I | BPE | 2145 | 1094 | 4 | 4 | 4 | 0.33 | ||
| e: Curun | Curu1 | 3 | P | AT | 2148 | 1089 | 5 | 5 | 5 | 0.54 | |
| Curu3 | 3 | I | AT | 2146 | 1092 | 2 | 2 | 2 | 0.24 | ||
| Curu11 | 2 | I | CP | 2047 | 1048 | 7 | 7 | 7 | 0.57 | ||
| Curungueo | f: Curun | Curu10 | 2 | I | CP | 2032 | 1048 | 5 | 5 | 5 | 0.47 |
| Curu9 | 3 | P | AT | 2003 | 1039 | 8 | 9 | 9 | 0.91 | ||
| Curu8 | 3 | P | BPE | 2000 | 1044 | 4 | 5 | 5 | 0.63 | ||
| Curu7 | 1 | I | AT | 1959 | 1028 | 2 | 2 | 2 | 0.3 | ||
| g: Curun | Curu4 | 1 | P | AT | 1938 | 1025 | 6 | 6 | 6 | 0.62 | |
| Curu6 | 3 | P | AT | 1942 | 1021 | 7 | 7 | 7 | 0.72 | ||
| Curu5 | 3 | P | AT | 1920 | 1021 | 8 | 9 | 9 | 0.82 |
*OH es el orden hidrológico del arroyo, según Strahler (1957). **La temporalidad de los arroyos se indica con I (intermitentes) y P (permanentes). ***BPE/O Bosque de Pino Encino / Oyamel; BPE Bosque de Pino Encino; AT Agricultura de Temporal; CP Cultivo Perenne.
La vegetación ribereña. En los 30 sitios analizados se midieron 805 individuos pertenecientes a 24 familias, 37 géneros y 56 especies (Fig. 2). Tres de las morfoespecies colectadas no se pudieron determinar, por esta razón fueron excluidas del análisis. Se identificaron 16 especies endémicas al territorio mexicano, así como tres con alguna categoría de riesgo dentro de la Norma Oficial Mexicana NOM-059-SEMARNAT-2010 [NOM-059] (Secretaría de Medio Ambiente y Recursos Naturales [Semarnat], 2010). Las 56 especies han sido incluidas en el repositorio de datos de Flores-Díaz y Martínez (2022).
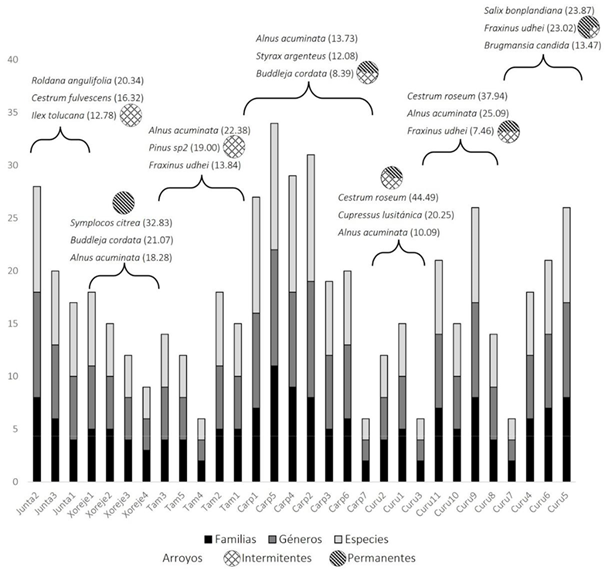
La composición y estructura de la vegetación remanente en las redes hidrológicas. Las redes Carpinteros y La Junta presentaron el mayor número de especies arbóreas (10 a 12 por sitio; Tabla 2). El ACP muestra que el número de especies, géneros y familias, junto con el número de ramas, y el número, altura y diámetro de los individuos, son las variables que más contribuyen a la primera dimensión; mientras la elevación y la precipitación son las que más contribuyen a la segunda, y en conjunto explican 71.1% de la variabilidad (Fig. 3). El agrupamiento jerárquico distinguió cuatro grupos en función de las variables mencionadas, con diferencias significativas para todas las variables excepto en la altura de los individuos (Kruskal-Wallis, p= 0.43). El grupo 1 (Fig. 3, color rosa) presenta el mayor número de familias, géneros y especies, y la mayor densidad de individuos. El valor más alto de área basal se encuentra en el grupo 4 (Fig. 3, color violeta) y los mayores diámetros en el grupo 3 (Fig. 3, color verde).
TABLA 2 Rasgos de la estructura de la vegetación ribereña remanente en el río San Juan Zitácuaro.
| Red | Sitio | Núm. Individuos | Altura promedio | Área Basal | Núm. Ramas | Diám. promedio |
| (ha) | (m) | (m2 ha-1) | (cm) | |||
| a: La Junta | Junta2 | 2,200.00 | 7.85 | 53.787 | 136 | 7.57 |
| Junta3 | 2,366.67 | 5.41 | 61.522 | 142 | 6.28 | |
| Junta1 | 1,200.00 | 6.25 | 14.136 | 60 | 6.43 | |
| b: Xorejé | Xoreje1 | 600 | 10.53 | 44.167 | 39 | 15.42 |
| Xoreje2 | 733.33 | 12.41 | 95.748 | 26 | 28.47 | |
| Xoreje3 | 300 | 7.78 | 22.901 | 14 | 15.63 | |
| Xoreje4 | 266.67 | 17.63 | 59.985 | 10 | 41.95 | |
| c: Tamejé | Tam3 | 233.33 | 10 | 46.726 | 7 | 38.65 |
| Tam5 | 333.33 | 9.6 | 36.426 | 12 | 26.82 | |
| Tam4 | 66.67 | 11.5 | 13.525 | 2 | 46.63 | |
| Tam2 | 466.67 | 10.07 | 36.076 | 23 | 14.53 | |
| Tam1 | 533.33 | 10.19 | 51.607 | 23 | 18.76 | |
| d: Carpin | Carp1 | 1,866.67 | 6.66 | 42.372 | 83 | 9.51 |
| Carp5 | 1,733.33 | 11.83 | 56.725 | 61 | 14.78 | |
| Carp4 | 1,366.67 | 11.5 | 68.127 | 45 | 18.58 | |
| Carp2 | 1,133.33 | 6.82 | 10.644 | 38 | 7.82 | |
| Carp3 | 766.67 | 5.35 | 5.081 | 34 | 6.51 | |
| Carp6 | 500 | 7.33 | 15.27 | 23 | 12.4 | |
| Carp7 | 400 | 17.08 | 17.335 | 16 | 19.17 | |
| e: Curun | Curu2 | 600 | 7.83 | 119.471 | 25 | 29.56 |
| Curu1 | 1,033.33 | 13.16 | 83.749 | 34 | 25.06 | |
| Curu3 | 133.33 | 10.28 | 9.867 | 4 | 30.56 | |
| f: Curun | Curu11 | 1,333.33 | 5.82 | 144.925 | 49 | 22.02 |
| Curu10 | 1,433.33 | 6.41 | 74.516 | 49 | 16.54 | |
| Curu9 | 566.67 | 8.5 | 19.475 | 24 | 11.64 | |
| Curu8 | 900 | 8.23 | 18.954 | 34 | 10.88 | |
| g: Curun | Curu7 | 66.67 | 7.84 | 9.713 | 3 | 29.07 |
| Curu4 | 800 | 12.83 | 77.228 | 33 | 18 | |
| Curu6 | 1,033.33 | 7.96 | 58.211 | 60 | 12.03 | |
| Curu5 | 1,433.33 | 13.75 | 62.245 | 63 | 13.58 |
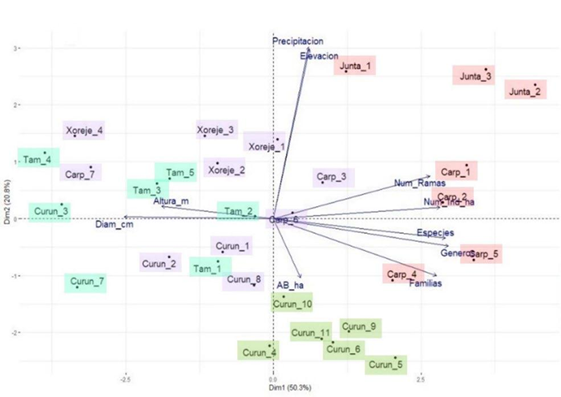
Las dos primeras dimensiones explican 71%. El agrupamiento jerárquico mostró cuatro grupos fisonómicos (colores).
FIGURA 3 Análisis de componentes principales.
En la red a): La Junta, están presentes Roldana angulifolia (DC.) H.Rob. & Brettell, Cestrum fulvescens Fernald, e Ilex discolor var. tolucana (Hemsl.) Edwin ex J.Linares, cuyo VI es de 20.37, 16.32 y 12.78, respectivamente. En el resto de las redes (b - g) es recurrente la presencia de Alnus acuminata Kunth, Buddleja cordata Kunth y Fraxinus udhei (Wenz.) Lingelsh, con altos VI (Fig. 2).
Diversidad de especies en las redes. Para las redes analizadas se obtuvo una representación de la riqueza de especies arbóreas (Tabla 1). La curva de rarefacción para las redes (Fig. 4) concuerda con los índices de diversidad. La red d: Carpinteros es la de mayor número de especies por individuos censados y, a pesar de que la red c: Tamejé tiene pocos individuos, estos están más uniformemente distribuidos entre las especies.
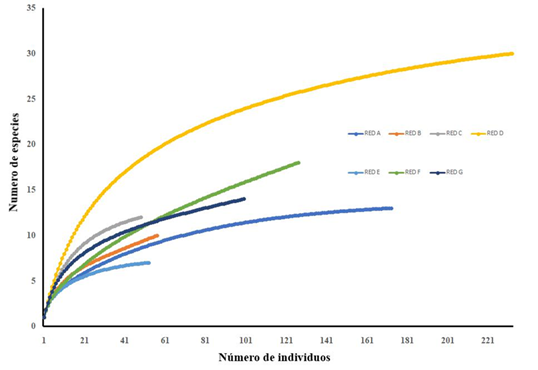
Los índices Sorensen Cuantitativo y β-Simpson, que muestran el recambio de especies, se presentan en la Tabla 3.
Tabla 3 Diversidad β en las redes analizadas.
| Red:a | Red:b | Red:c | Red:d | Red:e | Red:f | Red:g | |
| Red:a | 0.287 | 0.099 | 0.167 | 0.035 | 0.020 | 0.015 | |
| Red:b | 0.300 | 0.358 | 0.159 | 0.109 | 0.076 | 0.025 | |
| Red:c | 0.750 | 0.700 | 0.206 | 0.137 | 0.250 | 0.094 | |
| Red:d | 0.308 | 0.200 | 0.500 | 0.098 | 0.244 | 0.060 | |
| Red:e | 0.714 | 0.714 | 0.571 | 0.571 | 0.322 | 0.105 | |
| Red:f | 0.846 | 0.800 | 0.667 | 0.444 | 0.286 | 0.256 | |
| Red:g | 0.923 | 0.900 | 0.750 | 0.714 | 0.571 | 0.000 |
Sorensen Cuantitativo se ubica en la diagonal superior y β-Simpson en la inferior. En negritas se indican los valores más altos y en itálicas los más bajos.
La presencia de especies potencialmente invasoras y con alto VI en los sitios, mostró estar asociada con la densidad de individuos y con la interacción entre la elevación y el orden hidrológico (familia= binomial; Dev. Nula=41.45, Dev. Residual=28.38, AIC= 38.38). En este estudio las especies potencialmente invasoras son R. angulifolia, C. fulvescens y C. rostreum. Cupressus lusitanica Mill. no es invasora sino introducida en estas riberas.
El índice de Shannon-Weiner (H´) mostró una relación significativa (R2= 0.67, p < 0.05) con presencia de invasoras y densidad y altura de individuos, así como con las variables geográficas elevación y orden hidrológico (H´= 0.284 * Invasoras [0] - 0.000232 * Elevación - 0.0655 *Orden [2] + 0.0934 * Orden [3] + 0.000182 * Densidad -0.0256 * Altura - 1.19). Los sitios con densidad intermedia son los más diversos (R2 = 0.40, p < 0.05): H´ = 0.542D- 0.45D2 + 0.613.
Síntesis de características geográficas y rasgos de la vegetación en cada red hidrológica
Red a: La Junta. Se ubica a 2800 m de elevación, tiene arroyos temporales rodeados de bosque de pino encino / oyamel y precipitación anual de 1200 mm. Registró una alta densidad de individuos, cuya altura promedio es de 8 m. La riqueza está alrededor de H´ = 0.72 con alta diversidad beta con el resto de los sitios principalmente con la red g: Curungueo. Las especies más representativas (Fig. 2) fueron R. angulifolia, I. tolucana y C. fulvescens; ésta última es endémica.
Red fluvial b: Xorejé. Se ubica a 2600 m de elevación, tiene arroyos rodeados por bosque de pino encino, lluvia promedio anual de 1130 mm. Los individuos estudiados no mostraron una alta densidad y alcanzaron alturas de hasta 18 m. Los valores de diversidad estuvieron alrededor de H´= 0.6 y la mayor diversidad beta se presentó con la red fluvial c: Tamejé. Las especies con mayor VI fueron Symplocos citrea Lex. ex La Llave & Lex. (endémica, dominante y con estatus de protección por la NOM-059 (Semarnat, 2010)), B. cordata y A. acuminata.
Red fluvial c: Tamejé. Se ubica a 2400 m de elevación, con lluvias anuales de 1100 mm y arroyos temporales en una matriz de agricultura de temporal. Presentó baja densidad de individuos, que alcanzaban alturas de 12 m con los mayores diámetros (47 cm). Presentó valores de diversidad alrededor de H´= 0.71 y la diversidad beta más elevada se presentó con la red a: La Junta. Las especies con mayor VI fueron A. acuminata, Pinus sp.2. y F. udhei.
Red fluvial d: Carpinteros. Se ubica a 2300 m de elevación, en zonas de 1100 mm de precipitación anual. Los arroyos de esta red atraviesan un bosque de pino encino, con árboles de 17 m de altura promedio y sitios que varían en cuanto a densidad de individuos. La riqueza se encontró alrededor de H´= 0.97 y la mayor diversidad beta se presentó con la red f: Curungueo. Las especies con mayor VI fueron A. acuminata, Styrax argenteus C. Presl. y B. cordata.
Red fluvial e: Curungueo. Se ubica a 2100 m de elevación, con una precipitación anual de 1000 mm y arroyos que cruzan una matriz de agricultura de temporal. La densidad de individuos fue baja, alcanzando hasta 13 m de altura y diámetros de 33 cm. La riqueza de especies estuvo alrededor de H´= 0.54 y la diversidad beta fue elevada con las redes a: La Junta y b: Xorejé. Las especies con mayor VI fueron Cestrum roseum Kunth, C. lusitanica Mill. y A. acuminata.
Red fluvial f: Curungueo. Se ubica a 2000 m de elevación, en una región que presenta 1000 mm de lluvia anual. Los arroyos de esta red cruzan una matriz de agricultura de temporal y cultivos perennes (huertas de aguacate). La densidad de individuos fue intermedia, con alturas de hasta 9 m. y las áreas basales más elevadas, con valores de 43 000 cm2 ha-1. La riqueza de especies estuvo alrededor de H´= 0.91, la mayor diversidad beta con la red fluvial a: La Junta. Las especies con mayor VI fueron C. roseum, A. acuminata y F. undhei.
Red fluvial g: Curungueo. Se ubica a 2000 m de elevación, en una región que presenta 1000 mm de lluvia anual. Los arroyos de esta red atraviesan una matriz de agricultura de temporal. La densidad de individuos es intermedia, con alturas de hasta 14 m. La riqueza de especies fue muy variable alcanzando alrededor de H´= 0.82 en uno de los tramos, aunque en los demás fue baja. La diversidad beta fue elevada en comparación con la red a: La Junta. Las especies con mayor VI fueron S. bonplandiana Kunth, F. undhei y Brugmansia candida Pers.
Discusión
Las especies ribereñas en sus contextos geográficos
En la cabecera de la cuenca del río San Juan Zitácuaro analizada en el presente estudio, tres especies nativas tuvieron una mayor presencia en las riberas; de ellas, A. acuminata y F. udhei han sido referidas como especies asociadas a corrientes de agua en bosques templados (Conafor, s.f.), su madera es usada con fines culturales, medicinales y forestales (madera preciosa) se recomienda su introducción para la recuperación de suelos degradados, dada su fácil reproducción (Conafor, s.f.). La tercera especie es C. roseum, de porte arbustivo, esta mostró gran importancia fisonómica y estructural en estos sitios; aunque se conoce poco de su ecología, por su pertenencia a la familia Solanaceae y sus frutos carnosos, es posible suponer que su dispersión es zoocoria.
Lot et al. (s.f.) identificaron a Alnus jorullensis Kunth, Buddleja sp., A. acuminata, F. uhdei, Carpinus caroliniana Walt., Alnus sp. y Salix bonplandiana Kunth como ribereñas de la zona de Zitácuaro, siendo las dos últimas registradas en un río permanente; estos autores recomiendan el uso de A. jorullensis para la restauración ribereña. A. acuminata estuvo presente de manera importante en tramos fluviales ubicados debajo de los 2800 m de altitud, al igual que B. cordata y F. udhei. S. bonplandiana presentó un bajo VI y fue encontrada únicamente en los tramos ubicados alrededor de 1900 m de altitud. Algunas de estas especies ribereñas como A. acuminata, está presente en otras regiones como el río Xaltatempa, Puebla (Aguilar Luna, 2018) en elevaciones de 1451 m a 1680 m; y en el bosque templado del río Quila, Jalisco (Santiago-Pérez et al., 2014) entre 1876 m y 2200 m de elevación. Symplocos citrea tiene gran importancia en Xorejé y ha sido registrada también en riberas de bosque mesófilo (López-Pérez et al., 2011). En Tamejé se encontraron A. acuminata, Pinus sp2 y F. udhei, propias de sitios sombreados, algunos de los cuales pueden ser arroyos que corren en valles profundos, alrededor de 2400 m de altitud. En Carpinteros, tuvo una presencia importante S. argenteus, también identificada como hidrófila en bosque mesófilo (López-Pérez et al., 2011), cuyo crecimiento se ve favorecido alrededor de los 2300 m de altitud. En Curungueo, C. lusitanica parece haber sido incorporada por los habitantes de los terrenos adyacentes a la ribera, ya que se encontraron individuos coetáneos y distribuidos de manera uniforme, además de que no es una especie propia de esos sitios; es una especie incluida en la NOM-059 (Semarnat, 2010) con categoría de riesgo. La presencia de Cestrum roseum y Brugmansia candida indica un fuerte disturbio en esta red y su cercanía a centros de población (Conabio, 2006), estas especies se presentan también en zonas de cultivos inmersas en bosques de pino encino del centro de México (Martínez et al., 2020).
Tanto la densidad de individuos como la riqueza mostraron variar con la cubierta del suelo que es atravesada por el arroyo, como se registra también en regiones como la Sierra Madre de Chiapas (Díaz-Pascacio et al., 2018; Escalona-Domenech et al., 2021; Fernandes et al., 2016). Sin embargo, es necesario incluir en el análisis los cambios en el caudal, ya que en zonas como la red a: La Junta, donde los arroyos atraviesan el bosque de oyamel, se presentan especies como R. angulifolia que podría indicar alteraciones en el patrón del caudal (Greet et al., 2015). Esta red se encuentra en la interfaz entre la zona núcleo y la de amortiguamiento de la RBMM; el manejo forestal se detuvo hace 50 años, pero es creciente la extracción de agua de los arroyos para riego. El estudio de las especies potencialmente invasoras de las riberas puede alertar sobre dichos cambios (Catford et al., 2011; Hood y Naiman, 2000).
El reconocimiento de las especies ribereñas para guiar mejores prácticas de manejo
Los ejemplares de especies arbóreas remanentes en las zonas de ribera han sido dejados en pie por los pobladores locales. Su permanencia puede indicar que han resistido fenómenos naturales e impactos antrópicos de la zona, esto las haría potencialmente útiles en iniciativas de recu-peración fluvial. Los resultados aquí presentados incluyen especies listadas en la NOM-059 (Semarnat, 2010), que tienen gran relevancia por tratarse de un área natural protegida. Por su posición en la cabecera de la cuenca, los arroyos analizados están asociados a procesos como el intercambio de materiales entre la ribera y la columna de agua (Barmuta et al., 2009; Flores-Díaz et al., 2017), el balance entre caudal y sedimentos (Gurnell et al., 2006; Finlay, 2011; Masese et al., 2017; Wohl, 2017) y la integridad de la red hídrica (Castillo et al., 2012; Sánchez, 2007). Las funciones de las riberas fluviales están fuertemente asociadas a la interacción entre la dinámica de la vegetación y la temporalidad del flujo, su velocidad y el tamaño de la cuenca vertiente (Gurnell et al., 2006; Gomes et al., 2021).
Conclusiones
Este estudio muestra la riqueza arbórea de especies remanentes en las riberas de arroyos de cabecera y el recambio de especies entre las redes analizadas en el gradiente altitudinal. La presencia de especies potencialmente invasoras mostró estar asociada con la densidad de individuos, el índice de diversidad, el orden hidrológico y la elevación, lo cual sugiere la importancia de incorporar los rasgos de la red hídrica en los estudios de flora ribereña. El patrón del caudal y la vegetación son interdependientes, y forman parches heterogéneos a lo largo de la red hídrica. La elevación, la precipitación y el orden hidrológico establecen condiciones particulares para la vegetación de las riberas, por lo que es necesario analizarlos con los aspectos de manejo del territorio, ya sean los cambios de cubierta del suelo o de caudal, distinguiendo el impacto de cada uno de ellos de forma individual o acumulativa. Las especies indicadas como nativas y propias de la ribera pueden formar parte de la paleta vegetal en iniciativas de recuperación en arroyos de cabecera, la cual es una zona crítica para alcanzar las metas de conservación y de suministro de agua. Los estudios que busquen orientar las decisiones de manejo ribereño (conservación, restauración o aprovechamiento) requieren reconocer la unicidad de los tramos en cada arroyo y red. La interdependencia entre el caudal y la vegetación es uno de los patrones que las iniciativas de restauración podrían emular. En la RBMM es necesario evaluar las presiones que el cambio de uso del suelo de bosques a cultivos está generando sobre los ecosistemas dulceacuícolas, manifiesto en la pérdida de cubierta boscosa en las zonas adyacentes y en la propia ribera, y en los cambios en el caudal que podrían favorecer la presencia de invasoras.

