











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
A la distribución en el espacio y por clases de tamaños de las diferentes especies en una comunidad vegetal se le conoce como estructura (Gadow et al., 2007) y está determinada por factores bióticos y abióticos. Su importancia radica en que, desde el punto de vista ecológico, permite conocer la dinámica del bosque y, junto con la diversidad y densidad, son los principales atributos para la descripción de las comunidades vegetales forestales. Los bosques de Quercus (BQ) o bosques de encino, también llamados encinares, se establecen siguiendo patrones determinados por factores de heterogeneidad ambiental como el tipo de suelo, humedad, orientación de las laderas, variación de la precipitación y temperatura (Block y Meave, 2015; Aguilar-Romero et al., 2016), entre otros.
Las especies dominantes de los bosques de encino son, en general, árboles del género Quercus, y que, debido de la variedad de climas y condiciones fisiográficas del país (Suárez-Mota et al., 2015), han generado una de las comunidades vegetales más diversas (Villaseñor y Ortiz, 2014; Sabas-Rosales et al., 2015). Los encinares predominan principalmente en zonas montañosas de México, como en la Sierra Madre Occidental, Sierra Madre Oriental, Sierra Madre del Sur y Faja Volcánica Transmexicana (Rzedowski, 2006), en consecuencia, se han identificado más de 160 especies de encinos en el país, de las cuales 109 son endémicas; así, México es considerado como uno de los centros de origen y diversidad de este género (Valencia, 2004).
No obstante, son diversos los factores que intervienen en el deterioro y fragmentación de estas comunidades, de las cuales destacan las actividades antropogénicas (Encina-Domínguez et al., 2011) como el crecimiento demográfico, la agricultura y la ganadería (Pineda-Jaimes et al., 2009; Rosete-Vergés et al., 2014). Además, se ha sugerido que el cambio climático podría estar afectando su distribución, dadas las alteraciones en los patrones de temperatura y precipitación (Gutiérrez y Trejo, 2014).
Se ha abordado la descripción y análisis estructural y de diversidad de los BQ en diversas regiones de México. Investigaciones como la de Bravo-Bolaños et al. (2020) relacionan la alta heterogeneidad ambiental y el gradiente altitudinal con la composición, estructura y distribución de los BQ en el estado de Nayarit, México. Mientras que Block y Meave (2015) atribuyen la estructura y composición a la complejidad del pasado geológico en el cerro el Tepozteco, Morelos. Además, Endara-Agramont et al. (2012) sugieren que actividades como: tala, pastoreo e incendios, así como los asentamientos humanos podrían estar comprometiendo el reclutamiento y modificando la estructura de encinares de Quercus laurina en el centro de México. En tanto Olvera-Vargas y Figueroa-Rangel (2012) relacionan la diferencia estructural de las especies de Quercus a respuestas estratégicas para compartir los recursos y coexistir.
En otras regiones de México con BQ como Ixtlán de Juárez, Oaxaca, al sur del país, Ruiz-Aquino et al. (2015) consideran que la estructura y el patrón de distribución horizontal es atribuido a las estrategias de regeneración e influido por las prácticas silvícolas. Mientras que, al norte de México, en Coahuila, Encina-Domínguez et al. (2011) sugieren que el impacto antropogénico ocasiona bajas en la regeneración y densidades de árboles juveniles, lo que reduce y fragmenta estos bosques. En la Sierra de Santa Rosa, Guanajuato, en la Meseta Central, Martínez-Cruz et al. (2009) abordan la diversidad de estas comunidades. Otros trabajos de caracterización estructural en encinares de la FVT son los realizados por Rubio-Licona et al. (2011) en el Estado de México y Zacarias-Eslava et al. (2011) en el Cerro del Águila, Michoacán. La generación de conocimiento en los últimos 20 años sobre la estructura y diversidad de BQ en México ha tenido avances importantes. No obstante, dada la heterogeneidad de estas comunidades, es importante ampliar la información que permita entender la dinámica, las características y las condiciones básicas estructurales y de diversidad.
Objetivos
El presente trabajo tuvo como objetivo caracterizar la estructura y diversidad arbórea de los bosques de encino en dos microcuencas de la Faja Volcánica Transmexicana en el centro de México.
Materiales y métodos
Sitio de estudio
El área de estudio comprende dos sitios con BQ en dos microcuencas: cuenca Presa de Guadalupe (CPG) y cuenca Río Prieto (CRP), en el Estado de México (Fig. 1), ubicadas dentro de la FVT (Cervantes-Zamora et al., 2001), en la provincia Lagos y Volcanes del Anáhuac. Esta provincia fisiográfica es una región montañosa que atraviesa la República mexicana de oriente a occidente en el centro de México y es reconocida como centro de diversificación, endemismo y transición biogeográfica para una gran variedad de taxa incluyendo la familia Fagaceae a la que pertenecen las especies del género Quercus (Rzedowski, 1991; Suárez-Mota et al., 2013)
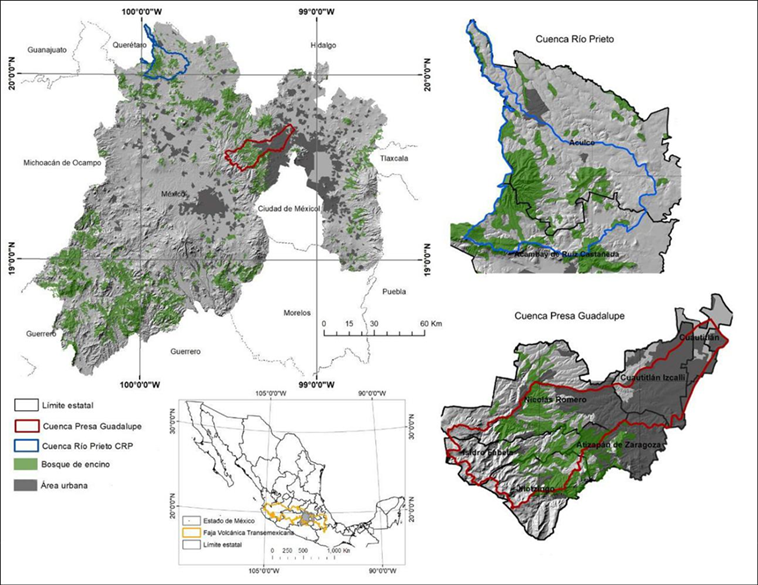
La CPG se localiza en la parte noroccidental de la Zona Metropolitana del Valle de México, y es en la parte media alta de la cuenca la zona donde se distribuyen los encinares y sitios de muestreo, correspondiendo con los municipios de Nicolás Romero, Isidro Fabela y Jilotzingo. La superficie aproximada es de 38 000 ha y en alrededor de 5500 predominan los BQ; además, otros tipos de vegetación presentes son los bosques de Abies religiosa y de Pinus hartwegii (Secretaría de Medio Ambiente y Recursos Naturales [Semarnat], 2014) en las partes altas > 3000 m. En cuanto a los tipos de suelo, predominan Andosol (70.6% en la zona boscosa) y Luvisol (27.5%) (Sotelo et al., 2006). Respecto al clima, predominan el templado subhúmedo con lluvias en verano C(w1) y el semifrío con lluvias en verano C(w2) (García, 1998), con un intervalo de temperatura de 6 °C a 14 °C y una precipitación entre 800 mm y 1300 mm.
La CRP se localiza al noroeste del Estado de México y abarca los municipios de Acambay y Aculco en una superficie aproximada de 58 000 ha, en las que 7000 ha corresponden al BQ y en menor medida existe bosque de pino (Semarnat, 2014). En esta región predomina el clima templado con lluvias en verano C(w1) con precipitación media anual de 700 mm a 1000 mm; los tipos de suelo en su mayoría son del tipo Pheaozem (47%), Luvisol (20.8%), Planosol (16.12%) y Vertisol (10.96%) (Sotelo et al., 2006).
Muestreo
Se realizó un muestreo subjetivo denominado preferencial estratificado, también llamado muestreo dirigido (Matteucci y Colma, 1982; Ferro-Díaz, 2015), que consistió en parcelas circulares de 1000 m2. La estratificación se hizo con base en los criterios de fisonomía característicos de los encinares de la región, cobertura del dosel mayor a 75% y un intervalo altitudinal entre 2600 m y 3100 m s.n.m., además de no contar con aprovechamiento forestal regulado. Se muestrearon 42 parcelas en CPG y 28 en CRP entre febrero y septiembre de 2017. Se midieron todos los individuos arbóreos con una dimensión mayor o igual a 5 cm de diámetro a la altura de pecho (DAP) o diámetro normalizado (DN), para lo cual se utilizó una forcípula. En tanto que, para la altura, al inicio de cada muestreo se utilizó un clinómetro, para tener una referencia base; posteriormente, para el resto de los ejemplares la altura se obtuvo visualmente a través de una medida aproximada; aunado a esto, se midió la altura y el diámetro de los tocones. Los ejemplares botánicos se identificaron en campo, cuando fue posible, y se colectaron muestras para su posterior identificación y verificación en herbario con base en Romero et al. (2002) y Rzedowski y Rzedowski (2005).
Estructura
La caracterización estructural se hizo con base en lo establecido por Curtis y McIntosh (1951); Mueller-Dombois y Ellenberg (1974) y Chai y Wang (2016), considerando las siguientes variables: dominancia, con base en el diámetro del tronco; abundancia, obtenida a través de la densidad (individuos por hectárea); área basal (m2 ha-1); y valor de importancia relativa (VIR), que se obtiene con la sumatoria de los valores relativos de densidad, área basal y frecuencia para todos los individuos censados.
La estructura horizontal se obtuvo mediante la clasificación en categorías de clases diamétricas en intervalos de 5 cm (5.0 cm - 9.9 cm, 10.0 cm - 14.9 cm, etc.). Para la representación gráfica de la distribución diamétrica, se eligieron las seis especies que se presentan en ambos sitios y que presentan altas y medias densidades. Aunado a esto, para determinar si existen diferencias entre los tamaños diamétricos de estas especies entre ambos sitios y debido a que los datos no cumplieron con los criterios de normalidad (Field, 2009), se aplicó la prueba no paramétrica U de Mann Whitney en el paquete estadístico SPSS.
Para el análisis de la estructura vertical se empleó el índice A de Pretzsch de distribución vertical de especies (Ecuación 1).
donde:
A : índice A de Pretzsch de distribución vertical de especies
S : número de especies presentes
Z : Número de estratos de altura
Pij : porcentaje de especies (i) en cada sitio (j)
donde:
Nij= número de individuos de la misma especie (i) en la zona (j)
N = número total de individuos
El índice A es una modificación del índice de Shannon, que se representa en tres estratos, con base en la altura máxima, lo que permite analizar la dinámica de las especies arbóreas a través de las proporciones y dimensiones de estas (Rubio-Camacho et al., 2014). Del índice A se derivan las ecuaciones 3 y 4.
donde:
Amax: valor máximo de A
Arel: porcentaje estandarizado del índice A
El resto de las variables se definieron con anterioridad
Los estratos se establecen con base en el árbol de mayor altura (100%): estrato I (80% - 100%), estrato II (50% - 80%) y estrato III (0% - 50%).
Diversidad
Para el cálculo de la diversidad se utilizó el software Past 4.3, partiendo de los datos del número de especies o riqueza específica (S) y la abundancia de estas en cada sitio. Se calcularon los índices Alfa (α) y de Shannon de acuerdo con Magurran (2004), al cual se aplicó la prueba de t propuesta por Hutcheson (Moreno, 2001) para determinar diferencias. Además, se estimó la equidad con base en Shannon (E) y Simpson (E1/D) (Monero, 2001; Magurran 2004).
La diversidad Beta (β) se define como el grado de reemplazo de especies a través de gradientes ambientales (Whittaker, 1972) y puede evaluarse con base en índices o coeficientes de similitud o disimilitud. Se empleó para ello el índice de Jaccard que puede calcularse con datos cualitativos (Moreno, 2001). Además, se utilizó el coeficiente de similitud de Sorensen, el cual se emplea para datos cuantitativos (Magurran 2004), lo cual fue corroborado con Bray-Curtis en PAST 4.3.
Resultados
La superficie de muestreo fue de 4.2 ha y 2.8 ha en CPG y CRP, respectivamente, en ese orden se midieron 3184 y 3856 individuos (total: 7040), con una densidad de 918 y 1138 árboles por hectárea. Las especies más abundantes en CPG fueron: Quercus laurina, Quercus rugosa, Quercus crassipes y Garrya laurifolia, las cuales en conjunto suman 738.57 individuos por hectárea, 80.44% de la densidad relativa. Las 18 especies restantes presentan una densidad de menos de 21 individuos por hectárea (Tabla 1). En esta misma tabla, se muestra la dominancia medida en área basal en la que estas cuatro especies suman 29.29 metros cuadrados por hectárea (m2/ha), el resto de las especies presentaron un área basal menor a 0.5 m2 ha-1. En consecuencia, la sumatoria del valor de importancia de estas cuatro especies es de 68.58.
TABLA 1 Atributos estructurales del bosque de la Cuenca Presa Guadalupe.
| Especie | Densidad (ind. ha-1) | Densidad Relativa (%) | Frecuencia | Frecuencia Relativa (%) | Área basal (m2 ha-1) | Área basal relativa (%) | Valor de importancia (VIR) |
| Quercus laurina | 250.48 | 27.28 | 17.00 | 11.18 | 12.35 | 36.48 | 24.98 |
| Quercus rugosa | 185.71 | 20.23 | 15.00 | 9.87 | 6.24 | 18.42 | 16.17 |
| Quercus crassipes | 171.43 | 18.67 | 12.00 | 7.89 | 8.57 | 25.30 | 17.29 |
| Garrya laurifolia | 130.95 | 14.26 | 15.00 | 9.87 | 2.13 | 6.29 | 10.14 |
| Prunus serotina | 20.48 | 2.23 | 12.00 | 7.89 | 0.39 | 1.14 | 3.75 |
| Arbutus xalapensis | 12.62 | 1.37 | 12.00 | 7.89 | 0.35 | 1.02 | 3.43 |
| Quercus obtusata | 30.24 | 3.29 | 7.00 | 4.61 | 0.75 | 2.21 | 3.37 |
| Abies religiosa | 18.57 | 2.02 | 7.00 | 4.61 | 1.03 | 3.03 | 3.22 |
| Salix paradoxa | 20.00 | 2.18 | 9.00 | 5.92 | 0.20 | 0.60 | 2.90 |
| Quercus castanea | 20.00 | 2.18 | 5.00 | 3.29 | 0.32 | 0.93 | 2.13 |
| Rhamnus sp. | 17.14 | 1.87 | 6.00 | 3.95 | 0.11 | 0.34 | 2.05 |
| Pinus patula | 2.62 | 0.29 | 7.00 | 4.61 | 0.27 | 0.80 | 1.90 |
| Buddleja cordata | 4.76 | 0.52 | 6.00 | 3.95 | 0.16 | 0.48 | 1.65 |
| Alnus jorullensis | 8.57 | 0.93 | 5.00 | 3.29 | 0.16 | 0.47 | 1.56 |
| Pinus montezumae | 2.86 | 0.31 | 4.00 | 2.63 | 0.25 | 0.75 | 1.23 |
| Quercus laeta | 8.57 | 0.93 | 3.00 | 1.97 | 0.17 | 0.49 | 1.13 |
| Clethra mexicana | 6.19 | 0.67 | 3.00 | 1.97 | 0.16 | 0.48 | 1.04 |
| Crataegus mexicana | 3.81 | 0.41 | 2.00 | 1.32 | 0.09 | 0.27 | 0.67 |
| Pinus leiophylla | 0.95 | 0.10 | 1.00 | 0.66 | 0.05 | 0.15 | 0.30 |
| Quercus crassifolia | 0.95 | 0.10 | 1.00 | 0.66 | 0.05 | 0.14 | 0.30 |
| Cupressus lusitanica | 0.71 | 0.08 | 1.00 | 0.66 | 0.00 | 0.01 | 0.25 |
| Pinus hartwegii | 0.24 | 0.03 | 1.00 | 0.66 | 0.00 | 0.01 | 0.23 |
| 918.10 | 100.00 | 152.00 | 100.00 | 33.86 | 100.00 | 100.00 |
En la CRP, las abundancias más altas corresponden a Q. laurina y Q rugosa, que entre ambas suman 86.4% de la densidad relativa; seguidas de A. xalapensis y Q. crassipes. Las ocho especies restantes presentan menos de 11 ha-1 individuos (Tabla 2). En cuanto a dominancia, estas cuatro especies suman 33.56% m2 ha-1, 95.13% de la densidad relativa y por consiguiente un VIR de 86.04. Mientras que los tocones representan 16% y 12% para CPG y CRP de la densidad del arbolado.
TABLA 2 Atributos estructurales del bosque de la Cuenca Río Prieto.
| Especie | Densidad (ind. ha-1) | Densidad Relativa (%) | Frecuencia | Frecuencia Relativa (%) | Área basal (m2 ha-1) | Área basal relativa (%) | Valor de importancia (VIR) |
| Quercus laurina | 500.71 | 44.03 | 26.00 | 22.22 | 16.42 | 46.55 | 37.60 |
| Quercus rugosa | 481.79 | 42.37 | 27.00 | 23.08 | 13.79 | 39.08 | 34.84 |
| Arbutus xalapensis | 55.71 | 4.90 | 19.00 | 16.24 | 1.25 | 3.53 | 8.22 |
| Quercus crassipes | 47.50 | 4.18 | 7.00 | 5.98 | 2.10 | 5.97 | 5.38 |
| Prunus serotina | 10.71 | 0.94 | 10.00 | 8.55 | 0.13 | 0.37 | 3.29 |
| Pinus teocote | 6.07 | 0.53 | 7.00 | 5.98 | 0.74 | 2.09 | 2.87 |
| Quercus crassifolia | 11.79 | 1.04 | 6.00 | 5.13 | 0.37 | 1.06 | 2.41 |
| Garrya laurifolia | 7.14 | 0.63 | 5.00 | 4.27 | 0.04 | 0.10 | 1.67 |
| Pinus montezumae | 5.36 | 0.47 | 3.00 | 2.56 | 0.36 | 1.03 | 1.36 |
| Crataegus mexicana | 3.93 | 0.35 | 4.00 | 3.42 | 0.03 | 0.09 | 1.28 |
| Alnus jorullensis | 0.71 | 0.06 | 2.00 | 1.71 | 0.00 | 0.01 | 0.59 |
| Clethra mexicana | 5.71 | 0.50 | 1.00 | 0.85 | 0.04 | 0.11 | 0.49 |
| Total | 1137.14 | 100.00 | 117 | 100 | 35.28 | 100 | 100 |
Estructura horizontal
En CPG, 70% de los diámetros se ubican en el intervalo de categorías de 5 cm a 22.5 cm, mientras que para CRP en 80%, lo que indica que hay un predominio de individuos jóvenes. En los dos sitios, Q. laurina y Q. rugosa son las dominantes, en tanto, A. xalapensis y Prunus serotina, presentan porcentajes bajos pero similares entre ellos (Fig. 2). En cuanto a Q. crassipes y G. laurifolia difieren ampliamente, la segunda contribuye con 17% de la densidad y categoría de 5 cm - 48 cm en los diámetros en CPG. En tanto que en CRP, G. laurifolia tiene 0.6% y categoría DN de 5 cm - 16 cm. El comportamiento de la distribución de los diámetros se asemeja a una J invertida en Q. laurina, Q. rugosa y A. xalapensis (Fig. 2, a, b y d), así como en G. laurifolia y P. serotina (Fig. 2, e y f); en tanto, para Q. crassipes esta tendencia se aleja de dicho patrón en ambos sitios en categorías menores a 10 cm y a 15 cm (Fig. 2. c).

FIGURA 2 Comportamiento de las distribuciones diamétricas de seis de las especies dominantes y codominantes presentes en ambos sitios.
La prueba no paramétrica de Mann-Whitney no mostró diferencias significativas en los diámetros entre CPG y CRP, para las especies A. xalapensis, G. laurifolia, Q. crassipes, Q. laurina y Q. rugosa (P = 0.032, 0,001, 0.042, 0.001, 0.001) correspondientemente, el valor de significancia fue de 0.05. Sin embargo, esta prueba determinó que P. serotina sí mostró diferencias en los diámetros entre los dos sitios (P = 0.897).
Estructura vertical
En CPG, la mayor altura identificada fue 30 m, medida a partir de la cual se definieron los estratos en: I (24 m - 30 m), II (15 m - 23 m), III (< 15 m). En el estrato III se presenta 89.16% de las alturas con presencia de 21 especies de las 22 que se registran. Para los estratos I y II tienen 8.27% y 2.57% y con 16 y 7 especies presentes, respectivamente (Tabla 3). El resultado de índice A fue de 2.07, Amax 4.23 y un Arel de 49.0%. En CRP los estratos se definieron en los siguientes: estrato I (22 m - 28 m), estrato II (14 m - 22 m) y estrato III (< 14 m). Del total de las alturas de los árboles, 66% se ubica en este último estrato, en donde las 12 especies tienen representatividad, no así en los estratos II y I en los que solo tienen presencia ocho y siete especies con 23.27% y 10.33% de estas en ese orden (Tabla 4).
TABLA 3 Valores del índice de distribución vertical (Pretzsch) de Cuenca Presa Guadalupe.
| Especie | Estrato I | Estrato II | Estrato III | |||||||||
| N | N ha-1 | Del total | En la zona | N | N ha-1 | Del total | En la zona | N | N ha-1 | Del total | En la zona | |
| Q. laurina | 78 | 18.5 | 78.0 | 2.0 | 171 | 40.7 | 53.6 | 4.4 | 803 | 191.1 | 23.3 | 20.8 |
| Q. rugosa | 14 | 3.3 | 14.1 | 0.3 | 26 | 6.2 | 8.1 | 0.7 | 740 | 176.1 | 21.5 | 19.1 |
| Q. crassipes | 3 | 0.7 | 3.0 | 0.0 | 46 | 10.9 | 14.4 | 1.1 | 671 | 159.7 | 19.5 | 17.4 |
| G. laurifolia | 1 | 0.2 | 1.0 | 0.0 | 14 | 3.3 | 4.4 | 0.3 | 535 | 127.3 | 15.5 | 13.8 |
| Q. obtusata | - | - | - | - | 5 | 1.2 | 1.5 | 0.1 | 122 | 29.0 | 3.5 | 3.1 |
| P. serotina | 1 | 0.2 | 1.0 | 0.0 | 4 | 0.9 | 1.2 | 0.1 | 81 | 19.3 | 2.3 | 2.1 |
| S. paradoxa | - | - | - | - | 5 | 1.2 | 1.5 | 0.1 | 79 | 18.8 | 2.3 | 2.0 |
| R. serrata | - | - | - | - | 0 | - | - | - | 72 | 17.1 | 2.0 | 1.8 |
| Q. castanea | - | - | - | 15 | 3.6 | 4.7 | 0.3 | 69 | 16.4 | 2.0 | 1.8 | |
| A. religiosa | 1 | 0.2 | 1.0 | 0.0 | 10 | 2.3 | 3.1 | 0.2 | 67 | 15.9 | 1.9 | 1.7 |
| A. xalapensis | 0 | - | - | - | 4 | 0.9 | 1.2 | 0.1 | 49 | 11.6 | 1.4 | 1.2 |
| A. jorullensis | 0 | - | - | - | 3 | 0.5 | 0.9 | 0.0 | 33 | 7.9 | 0.9 | 0.8 |
| Cl. mexicana | 0 | - | - | - | - | - | - | - | 26 | 6.4 | 0.7 | 0.6 |
| Q. laeta | 0 | - | - | - | 10 | 2.5 | 3.1 | 0.2 | 26 | 6.4 | 0.7 | 0.6 |
| B. cordata | 0 | - | - | - | 1 | 0.5 | 0.3 | 0.0 | 19 | 4.5 | 0.5 | 0.5 |
| Cr. mexicana | 0 | - | - | - | 0 | - | - | - | 16 | 3.8 | 0.4 | 0.4 |
| P. montezumae | 0 | - | - | - | 2 | 0.5 | 0.6 | 0.0 | 10 | 2.4 | 0.3 | 0.2 |
| P. patula | 0 | - | - | - | 2 | 0.5 | 0.6 | 0.0 | 10 | 2.4 | 0.3 | 0.2 |
| P. leiophylla | 0 | - | - | - | 0 | - | - | - | 4 | 0.9 | 0.1 | 0.1 |
| C. lussitanica | 0 | - | - | - | 0 | - | - | - | 3 | 0.7 | 0.0 | 0.0 |
| Q. crassifolia | 0 | - | - | - | 1 | 0.2 | 0.3 | 0.0 | 3 | 0.7 | 0.0 | 0.0 |
| P. hartwegii | 1 | 0.2 | 1.0 | 0.0 | - | - | - | - | - | - | - | - |
| Total | 99 | 23.6 | 100 | 2.6 | 319 | 75.9 | 100 | 8.2 | 3438 | 818.5 | 100 | 89.1 |
TABLA 4 Valores del índice de distribución vertical (Pretzsch) de Cuenca Río Prieto.
| Especie | Estrato I | Estrato II | Estrato III | |||||||||
| N | N ha-1 | Del total | En la zona | N | N ha-1 | Del total | En la zona | N | N ha-1 | Del total | En la zona | |
| Q. laurina | 249 | 88.9 | 75.6 | 7.8 | 353 | 126. | 47.6 | 11.0 | 800 | 285.7 | 37.8 | 25.1 |
| Q. rugosa | 55 | 19.6 | 16.7 | 1.7 | 320 | 114. | 43.1 | 10.0 | 974 | 347.8 | 46.0 | 30.5 |
| Q. crassipes | 12 | 4.2 | 3.6 | 0.3 | 38 | 13. | 5.1 | 1.1 | 83 | 29.6 | 3.9 | 2.6 |
| P. teocote | 6 | 2.1 | 1.8 | 0.1 | 4 | 1.4 | 0.5 | 0.1 | 7 | 2.5 | 0.3 | 0.2 |
| P. montezumae | 3 | 1.0 | 0.9 | 0.0 | 1 | 0.3 | 0.1 | 0.0 | 11 | 3.9 | 0.5 | 0.3 |
| Q. crassifolia | 3 | 1.0 | 0.9 | 0.0 | 17 | 6.0 | 2.2 | 0.5 | 13 | 4.6 | 0.6 | 0.4 |
| A. xalapensis | 1 | 0.3 | 0.3 | 0.0 | 7 | 2.5 | 0.9 | 0.2 | 148 | 52.8 | 7.0 | 4.6 |
| P. serotina | - | - | - | - | 1 | 0.3 | 0.1 | 0.0 | 29 | 10.3 | 1.3 | 0.9 |
| G. laurifolia | - | - | - | - | - | - | - | - | 20 | 7.1 | 0.9 | 0.6 |
| Cl. mexicana | - | - | - | - | - | - | - | - | 16 | 5.7 | 0.7 | 0.5 |
| Cr. mexicana | - | - | - | - | - | - | - | - | 11 | 3.9 | 0.5 | 0.3 |
| A. jorullensis | - | - | - | - | - | - | - | - | 2 | 0.7 | 0.0 | 0.0 |
| Total | 329 | 117.5 | 100 | 10.3 | 741 | 264.6 | 100 | 23.2 | 2114 | 755 | 100 | 66.3 |
Diversidad
Se obtuvo riqueza específica S = 22 para CPG y S = 12 para CRP, agrupada en 13 géneros y 11 familias; Fagaceae fue la más representativa con 7 especies, seguida de Pinaceae (6), Rosaceae (2) y el resto de las familias con un solo género y una especie (Tabla 5). En cuanto a diversidad, el índice α de Fisher señaló mayor diversidad en CPG, y una situación similar mostró el de Shannon 2.074 y 1.347. La equidad con base en Shannon (E) y Simpson (E1/D) mostró valores más altos para CPG (Tabla 6). Sin embargo, la prueba de t indicó que no hay diferencias significativas en la diversidad; aunado a esto la prueba de varianzas de Fisher señaló igualdad de varianzas. En diversidad beta se obtuvieron los siguientes valores: Jaccard, 0.479; Sorensen, 0.60, lo que indica alrededor de 60% de similitud, corroborado con Bray-Curtis 0.598.
TABLA 5 Familias, géneros y especies en los sitios de estudio.
| Familia | Género | Núm. de especies | Especies |
| Quercus laurina Bonpl., Quercus obtusata Bonpl, Quercus crassipes | |||
| Fagaceae | Quercus | 7 | Bonpl, Quercus crassifolia Bonpl. Quercus castanea Née, Quercus |
| obtusata Bonpl. Quercus laeta Liebm. | |||
| Pinus montezumae Lamb., Pinus patula Schltdl. & Cham., Pinus | |||
| Pinaceae | Pinus | 5 | teocote Schltdl. & Cham., Pinus leiophylla Schl. & Cham., Pinus |
| hartwegii Lindl. | |||
| Abies | 1 | Abies religiosa (Kunth) Schltdl. & Cham. | |
| Cupressaceae Rosaceae | Cupressus | 1 | Cupressus lusitanica Mill. |
| Prunus | 1 | Prunus serotina Ehrh. | |
| Crataegus | 1 | Crataegus mexicana DC. | |
| Garryaceae | Garrya | 1 | Garrya laurifolia Hartw. Ex Benth. |
| Ericaceae | Arbutus | 1 | Arbutus xalapensis Kunth |
| Clethraceae | Cletrhra | 1 | Clethra mexicana DC. |
| Scrophulariaceae | Buddleja | 1 | Buddleja cordata Kunth. |
| Rhamnaceae | Rhamnus | 1 | Rhamnus sp. |
| Saliceae | Salix | 1 | Salix paradoxa Kunth. |
| Betulaceae | Alnus | 1 | Alnus jorullensis Kunth. |
Discusión
Abundancia y dominancia
Las especies más abundantes y dominantes en ambos sitios de estudio fueron Q. laurina, Q rugosa y Q. crassipes. Por consiguiente, la densidad en conjunto de ambos sitios de estudio es de 1027 individuos por hectárea, situación que se ha descrito en diversos trabajos (Zacarias-Eslava et al., 2011; Sánchez-González y López-Mata 2003) realizados en la FVT y en intervalos de 2500 m a 3000 m. En este sentido, Rzedowski (2006) refiere que, en altitudes superiores a los 2400 m de esta provincia fisiográfica, son comunes los bosques en los que dominan estas tres especies. Zacarias-Eslava et al. (2011) registraron para el bosque de Q. rugosa en Michoacán una densidad de 940 individuos por hectárea, mientras que Sánchez-González y López-Mata (2003), en La Sierra Nevada, documentaron que Q. rugosa, Q. laurina y A. xalapensis tienen los VIR más altos. Por su parte, Block y Meave (2015) determinaron como especie dominante a Q. rugosa y en menor medida a A. xalapensis y Q. laurina en encinares del estado de Morelos.
En otras provincias fisiográficas, como la Meseta Central y la Sierra Madre del Sur, se han analizado asociaciones de Quercus, con condiciones similares a lo aquí expuesto. En la primera, Martínez-Cruz et al. (2009) en bosques de encinos en Guanajuato (2600-2750 m) encontraron una densidad de la asociación Q. laurina-Q. rugosa de 1133 individuos por hectárea. De modo similar, en Oaxaca (2700 m), Ruiz-Aquino et al. (2015) observaron la densidad de 1200 ha-1 y Q. laurina comparte dominancia con Q. crassifolia, a diferencia de los sitios de esta investigación donde la especie en mención comparte dominancia con Q. rugosa y Q. crassipes.
Lo anterior sugiere que las condiciones de densidad para la asociación vegetal de Q. rugosa-Q. laurina oscilan entre 882 in ha-1 y 1208 in ha-1, y con cobertura del dosel en más de 50%. Los factores fisiográficos y ambientales implicados serían la altitud mayor a 2500 m y hasta los 3100 m y precipitaciones mínimas de 790 mm. Aunado a esto, de acuerdo con Aguilar-Romero et al. (2016), la humedad y temperatura estarían interviniendo en la relación y distribución de las especies de encino. Además, esta asociación estaría acompañada generalmente de otras especies arbóreas de dominancia intermedia como Q. crassipes, A. xalapensis y Q. crassifolia.
Las especies A. xalapensis, P. serotina y G. laurifolia obtuvieron valores intermedios de VIR. En particular, G. laurifolia en CPG obtuvo alta densidad, ocupando el cuarto sitio. Rzedowski (2006) menciona que este género se asocia a bosques dominados por Quercus, es más característico en fondos de cañadas, arroyos o hábitats similares; pero también se le encuentra en bosques mesófilos de montaña en la FVT y es de amplia distribución (Carranza González, 1996; López-Pérez et al., 2011). Los factores de mayor humedad y tala podrían favorecer el incremento de la abundancia de esta especie en estos bosques, ya que CPG tiene una precipitación de hasta 1300 mm, en contraste con CRP con 1000 mm como precipitación máxima y dada su densidad podría estar interviniendo en la disminución del reclutamiento de algunas especies como Q. crassipes. Por su parte, A. xalapensis en CRP ocupa el tercer sitio en cuanto a VIR, a su vez en CPG se ubica hasta en la sexta posición, por debajo incluso de P. serotina; de acuerdo con Granados et al. (2014), esta especie se establece preferentemente en los bordes del bosque y zonas de transición entre el borde y el núcleo. Además, es de amplia distribución en montañas del norte y Centroamérica (Rzedowski, 2006; González y González, 2014).
Estructura horizontal
Las poblaciones de las especies dominantes y codominantes no difieren entre ambos sitios y solo lo hace P. serotina. Bajo este contexto, la estructura de un bosque dominado por árboles con diámetros menores o categorías diamétricas inferiores (5 cm -10 cm) tendrá un alto número de individuos, mientras que conforme se incrementa la categoría diamétrica la cantidad de individuos disminuirá (> 50 cm), esto sugiere que está ocurriendo un reclutamiento permanente de individuos, lo cual gráficamente se configura como una forma de J invertida (Lamprecht, 1990; Newton, 2007; Restrepo et al., 2012) y es característico de especies tolerantes o semitolerantes a la sombra y su codominancia podría deberse a una etapa de sucesión intermedia equilibrando la estratégicamente la composición y estructura (Olvera-Vargas y Figueroa-Rangel, 2012).
Quercus laurina, y Q. rugosa muestran el patrón de J invertida en ambos sitios (Fig. 2 a y b); sin embargo, la segunda en CPG presenta una ligera disminución en la categoría menor a 10 cm, lo que indica que se podría afectar el reclutamiento de esta especie, y posiblemente estaría siendo desplazada por Q. laurina, ya que esta tiene un alto porcentaje de individuos en el estrato alto, lo que disminuye la cantidad de radiación solar para Q. rugosa, que se considera de poca tolerancia a la sombra (Granados et al., 2014).
En los dos sitios de estudio, Q. crassipes es la única que difiere del comportamiento de la distribución diamétrica, distribución que se conoce como bimodal asimétrica (Prodan et al., 1997), ya que presenta menor cantidad de individuos en las primeras categorías (< 15 cm). En este sentido, Olvera-Vargas y Figueroa Rangel (2012) plantean que, aunque esta especie es tolerante a la sombra, tiene un mejor desarrollo en sitios abiertos, como en claros, bordes del bosque y orillas de caminos (Granados et al., 2014). En esta investigación se trabajó solo al interior del bosque, de modo que se puede compartir el planteamiento de la tolerancia a la sombra. Sin embargo, se sugiere que la disminución de individuos jóvenes podría deberse a presiones de tipo natural como la competencia por los recursos como la disponibilidad de radiación solar, en particular con Q. rugosa y Q. laurina debido a su altura en ambas microcuencas; además se cree que estarían interviniendo otros factores como los nutrientes en el suelo y las diferentes estrategias de regeneración (Ruíz-Aquino et al., 2014). Otro aspecto para considerar es que, de acuerdo con Hernández (2002) las personas que talan árboles para leña y carbón en encinares del centro de México suelen preferir diámetros de entre 20 cm y 50 cm; de manera que, esta disminución no se debería a esta actividad, porque además los tocones medidos tienen un diámetro promedio de 20.6 cm.
La presencia de tocones indica que persiste la tala no regulada; aun cuando en estos sitios es menor en comparación, por ejemplo, con lo que sucede en encinares del Área de Protección de Flora y Fauna del Nevado de Toluca, en donde, Endara et al. (2012) registran que, la extracción maderable por tala asciende a 29% de la densidad, y como consecuencia hay regeneración abundante proveniente de rebrotes. Por consiguiente, se considera que los cambios en la estructura del dosel ocasionados por actividades humanas influyen en la pérdida de biodiversidad y pueden favorecer el establecimiento o invasión por especies exóticas en estos bosques (Encina-Domínguez, 2011; Del-Val et al., 2015).
Estructura vertical
La distribución vertical muestra para CPG que las alturas se concentran en el estrato de III (< 15 m; Fig. 3), mientras que en CRP se distribuyen preferentemente tanto en los estratos II como en III, es decir, media y baja altura; de acuerdo con Rzedowski (2006) en los encinares se pueden distinguir hasta tres estratos. Esta distribución vertical es similar a lo observado en bosques templados en el norte de México (Rubio-Camacho et al., 2014; Dávila-Lara et al., 2019; Silva-García et al., 2021). No obstante, Martínez-Cruz et al. (2009) definen, en términos de estructura vertical, la asociación Q. laurina-Q. rugosa como de estrato alto, debido a que comparan con otras asociaciones de Quercus de menor altura. Las alturas promedio identificadas en este trabajo coinciden con lo obtenido por el índice de Pretzsch, en ambos sitios la altura se ubica en estrato bajo.

FIGURA 3 Estructura vertical de las especies representativas de los sitios de estudio. Pinus sp, Psp; Quercus laurina, Ql; Quercus rugosa, Qr; Quercus crassipes, Qc; Arbutus xalapensis, Ax; Garrya laurifolia, Gl; Prunus serotina, Ps. Elaboración propia, estratos con base en Pretzsch (2009).
Diversidad
En la CPG se obtuvo una diversidad específica mayor que CRP, ambas con amplia representatividad de especies de Quercus, los índices muestran que en general los valores de diversidad fueron mayores en la primera; sin embargo, estos no muestran diferencias significativas. Esta situación coincide con lo que sostienen Martínez-Cruz et al. (2009) para la asociación Q. rugosa-Q laurina, quienes registraron menor diversidad en comparación con otras asociaciones de este género y lo atribuyen a variables estructurales como la densidad y el área basal, aunado a las diferencias de composición y abundancia de especies. Por su parte, Ruiz-Aquino et al. (2015) consideran que una menor diversidad en bosques de Quercus también se debe a la alta dominancia de algunas especies del género. En este sentido, en la CRP solamente tres especies por sí mismas alcanzan más de 80% de VIR, mientras que para CPG se requieren ocho especies para alcanzar este valor.
Otra variable que influye en la diversidad en estas comunidades vegetales es la altitud. Al respecto, Medrano et al. (2017), en alturas superiores a 2700 m en bosques de Durango obtuvieron valores bajos de diversidad en comparación con intervalos altitudinales inferiores. Mientras que, Sánchez-González y López-Mata et al. (2003) refieren que esta, junto con variables ambientales y condiciones del suelo, influyen en los patrones de composición, estructura y distribución de encinares de la FVT. La altitud considerada en este trabajo fue por encima de los 2600 m y Martínez-Cruz et al. (2009) en intervalo similar, de 2600 m - 2750 m, identificaron una asociación de bosque de encino como la aquí descrita.
Una de las características de los bosques templados en México es que concentran alta diversidad de plantas vasculares, incluyendo niveles de endemismos más altos para el país (Villaseñor y Ortiz, 2014) aunque los que más destacan son los bosques mixtos. No así los encinares, como los descritos en este estudio y los ya citados, realizados en otras regiones y que muestran niveles bajos de diversidad arbórea. A pesar de esto, la FVT es considerada una de las regiones más biodiversas (Suárez-Mota et al., 2013), incluso, de las más ricas en especies de Quercus en México (Valencia, 2004).
Los valores de riqueza, diversidad y recambio indican que las microcuencas de este estudio, en términos de la asociación florística de Q. laurina-Q. rugosa, tienen similitud. Sin embargo, la CPG es una zona más heterogénea en cuanto a las especies arbóreas que la componen, lo cual podría ser atribuido a condiciones de humedad y climáticas, así como a la transición con otras asociaciones y tipos de vegetación como los bosques mixtos de pino-encino y de Abies religiosa; incluso a una mayor interacción con actividades antrópicas como la tala ya que esta cuenca registró más tocones.
Conclusiones
Los resultados de este trabajo muestran que las especies Quercus laurina y Quercus rugosa son dominantes en estos sitios y conforman una asociación florística característica del centro de México y de la Faja Volcánica Transmexicana. Sin embargo, Q. crassipes presenta una disminución de individuos jóvenes en las poblaciones de ambas microcuencas; se presume que esto es debido a un posible desplazamiento por la competencia de acceso a recursos con otras especies. Sin embargo, los objetivos y alcances de esta investigación no profundizan al respecto, por lo que es necesario dar seguimiento al comportamiento poblacional de las especies arbóreas y los cambios en la diversidad que componen esta comunidad vegetal.
Hay otras especies que pueden ir formando parte de la comunidad por las etapas naturales de sucesión ecológica. No obstante, es posible que además de lo anterior, factores antrópicos como la tala y los incendios forestales generen cambios en la estructura y la diversidad de los encinares. Por lo anterior, es importante analizar los procesos ecológicos de estos bosques, ya que son ecosistemas frágiles y propensos a la transformación por actividades humanas.

