











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Los bosques templados brindan una serie de servicios ecosistémicos como la regulación del clima, captura de carbono y contribuyen a la recarga de mantos acuíferos, entre otros (Solís, Blanco, Sáenz, Méndez y López, 2018). Sin embargo, algunos bosques presentan distintas alteraciosnes (Mápula, López, Vargas y Hernández., 2007; Velasco-García et al., 2007). Debido a ello, es necesario generar información sobre la estructura de estos bosques a escala de rodal, que brinde soporte para la toma de decisiones sobre su manejo, conservación y restauración.
La estructura del bosque puede ser caracterizada a nivel de diversidad de especies, dimensiones y a nivel espacial (Chávez-Flores, Corral-Rivas, Vega-Nieva, López-Serrano y Rubio-Camacho, 2020; Gadow, Sánchez y Álvarez, 2007; Kulik, Molas y Rodríguez, 2019; Lin, Lam, Gadow y Kershaw, 2020; Mena-Mosquera, Andrade y Torres-Torres, 2019; Pommerening y Stoyan, 2020). Sin embargo, la investigación sobre estructura forestal en México ha sido enfocada a caracterizar la diversidad de especies y de dimensiones, siendo la distribución espacial una de las menos utilizadas, aunque brindan un mayor nivel de detalle. Estos estudios son importantes para determinar los cambios que originan los patrones y la variación espacial de la estructura del rodal, así como los efectos en la subsecuente sucesión de los ecosistemas forestales a escala de rodal (Yu, Wiegand y Yang 2009; Gu, Li, Qi y Wang, 2019).
Los análisis estructurales se han realizado a través de distintos índices y métodos que brindan información para la descripción de los ecosistemas; por ejemplo la estructura horizontal y vertical ha sido evaluada por medio del índice de valor de importancia (IVI), diagramas de perfil (Ogawa, Yoda, Ogino y Kira, 1965; Organización de las Naciones Unidas para la Educación, la Ciencia y la Cultura [Unesco], 1980), determinando pisos de altura (Lamprecht, 1990) o por medio del índice de A (Pretzsch, 2009); mientras que las relaciones de vecindad se han descrito usando el método de muestreo de los cinco árboles, desarrollado para evaluar los atributos estructurales (dimensiones, especies y la regularidad de sus posiciones) de los árboles (Aguirre, Hui, Gadow y Jiménez, 2003; Gadow, Real y Álvarez, 2001).
En algunos estudios llevados a cabo tanto dentro de México como a escala internacional se ha evaluado la estructura y distribución espacial de comunidades forestales, tratando de responder a distintas preguntas de investigación. En bosques templados, por ejemplo, se han utilizado para caracterizar la estructura de rodales con algún grado de perturbación (Ávila-Flores, González-Tagle, Jiménez-Pérez, Aguirre-Calderón, Treviño-Garza, Vargas-Larreta y Alanís, 2014; Rubio-Camacho, González-Tagle, Himmelsbach, Ávila-Flores, Alanís-Rodríguez y Jiménez-Pérez, 2017), también para comprender la distribución espacial de comunidades arbóreas como bosques de pino - encino en Oaxaca y bosques subtropicales caducifolios en China (Castellanos-Bolaños, Treviño-Garza, Aguirre-Calderón, Jiménez-Pérez y Velázquez-Martínez, 2010; Du, Hu, Zeng, Wang, Peng, Zhang y Song, 2017), conocer relaciones interespecíficas entre especies (Wédjangnon, Kuiga, Houêtchégnon y Ouinsavi, 2020). Asimismo, se han utilizado para contrastar la estructura de bosques con y sin manejo forestal (García, Olivares y Ramos, 2013; García, Narváez, Olivas y Hernández, 2019; Hernández, Aguirre, Alanís, Jiménez, Treviño, González, Lujan, Olivas y Domínguez, 2013) y para diseñar patrones de plantación que aumenten la competencia asimétrica y la resistencia de los ecosistemas (Zhang, Hui, Hu, Zhao, Guan, Gadow y Zhang, 2019).
Sin embargo, existen escasos estudios que evalúen los patrones de distribución espacial en bosques de alto valor de conservación, que presentan comunidades vegetales con especies vulnerables. Un ejemplo de estos ecosistemas es el bosque de Pseudotsuga menziesii, que tiene escasa distribución natural en México, encontrándose en la Sierra Madre Occidental y Oriental, así como en el centro - sur del país. Las poblaciones P. menziesii se encuentran fragmentadas en rodales o manchones aislados, normalmente dominados por otras especies (Debreczy y Racz, 1995; Domínguez-Calleros, Chávez-Flores, Rodríguez-Téllez, Corral-Rivas, Goche-Telles y Díaz-Vásquez, 2014; Rzedowski, 1978).
Diversos factores amenazan los bosques de Pseudotsuga en México, ya que tiene problemas de regeneración y desarrollo (Torres, 2006; Mápula et al., 2007). Entre los factores más importantes se encuentran el cambio de uso de suelo, incendios, sobrepastoreo, tala clandestina y plagas (Mápula et al., 2007; Ventura, López, Vargas y Guerra, 2010; Velasco-García, López-Upton, Angeles-Pérez y Guerra-de la Cruz, 2007). Sin embargo, uno de los factores que más efectos podría tener es el cambio climático, ya que las poblaciones de Pseudotsuga meziesii emigraron hacia el sur durante las glaciaciones, por lo que son sensibles a los cambios de temperatura a escala global ocasionando, por un lado que las áreas con condiciones adecuadas para su desarrollo y establecimiento se reduzcan y contraigan hacia las partes más altas de los sistemas montañosos y hacia el hemisferio norte (Hermann, 1985; Mápula-Larreta, López-Upton, Vargas-Hernández y Hernández-Livera, 2008). De ahí que el hábitat de Pseudotsuga menziesii esté compuesto de especies altamente especializadas, con alta biodiversidad y alto número de especies amenazadas (Schwendtner, Recalde, Alcalde, Gómez y Cárcamo, 2005).
Objetivos
El objetivo de este estudio fue caracterizar los patrones de distribución espacial de un bosque maduro en el Estado de Chihuahua, México. Para ello, se plantearon las siguientes preguntas de investigación: 1) ¿La distribución del arbolado es aleatoria? 2) ¿La mezcla de especies es uniforme? 3) ¿Existe homogeneidad dimensional a nivel espacial? 4) ¿Cuáles son las principales diferencias y/o similitudes con otros estudios? Dar respuesta a estas preguntas proporcionará información básica para la toma de decisiones en cuanto al manejo, conservación y restauración de estos ecosistemas de alta importancia ecológica, económica y social.
Materiales y métodos
Área de estudio
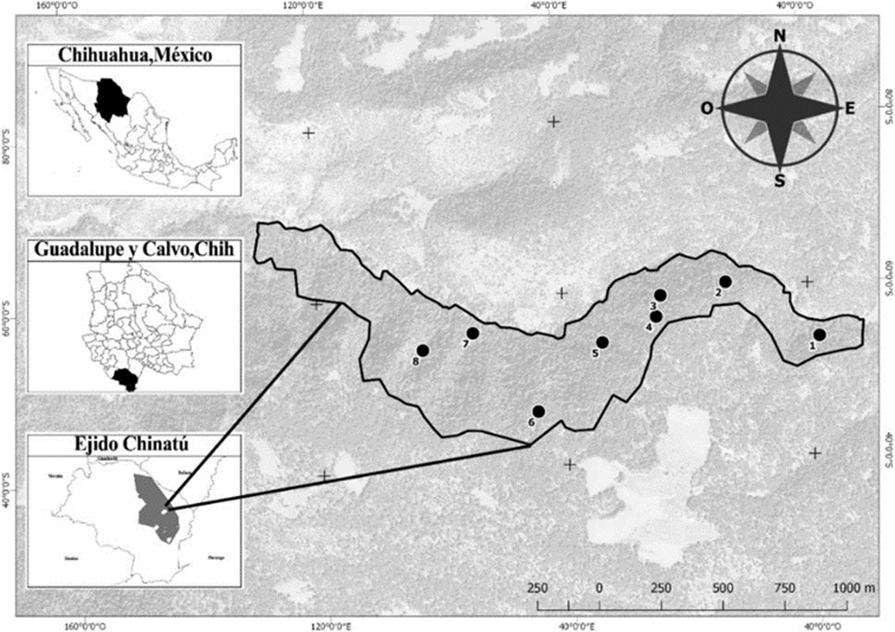
El estudio se llevó a cabo en el paraje “El Triste”, ubicado en el ejido Chinatú, municipio de Guadalupe y Calvo, al suroeste del estado de Chihuahua. Geográficamente se encuentra entre las coordenadas 26°14’ 43” y 106°38’ 37” (Fig. 1). Dicho bosque tiene una superficie de 80.46 ha-1 y altitudes entre los 2530 m y 2830 m s.n.m. Pertenece a la Región hidrológica Río Fuerte, a su vez a la provincia fisiográfica Sierra Tarahumara y a la subprovincia Gran Meseta y Cañones Chihuahuenses. El tipo de suelo predominante en el área es litosol. El clima de la región está clasificado como templado subhúmedo (Cw), con temperatura media anual de 13.7 °C y una precipitación pluvial media anual de 1126.8 mm (Chávez, 2009). El bosque está conformado principalmente por P. menziesii, P. arizonica y Quercus tuberculata, con presencia de especies importantes para la conservación como Abies durangensis y Picea chihuahuana. Dicha área se encuentra catalogada como “Relicto de bosque viejo o sobremaduro” de acuerdo con Galván-Moreno (2016) y se encuentra excluida del aprovechamiento forestal en el programa de manejo.
Toma de datos
Se establecieron ocho sitios aleatorios circulares de 1000 m2 para el levantamiento de información. Dicho número de sitios se determinó de acuerdo con un premuestreo realizado y de acuerdo con una intensidad de muestreo de 1%. Se consideraron individuos con diámetro normal (DN) (d1.30) ≥ 7.5 cm, además se registró la altura total (h, m), distancias de los árboles al centro del sitio, azimut y especie de cada individuo.
Análisis de la información
Se realizó el cálculo de la densidad (número de árboles por hectárea, ha-1), área basal (m2 ha-1), diámetro normal promedio (d1.30 (cm)) y altura total (m) promedio para conocer de manera general las características dasométricas de cada uno de los géneros presentes en el bosque. La caracterización de los patrones de distribución espacial del bosque se obtuvo utilizando técnicas de análisis de patrón de puntos y relaciones de vecindad basados en el método de los cinco árboles (Albert, 1999; Hui y Hu, 2001) el cual toma un árbol de referencia (i) y sus cuatro vecinos más cercanos
Índice de uniformidad de ángulos (Wi)
Se basa en la medición de los ángulos entre los vecinos a un árbol de referencia i y su comparación con un ángulo estándar α, de tal manera que al considerar cuatro vecinos al árbol de referencia Wi puede tomar valores de 0 hasta 1, donde un valor cercano a cero representa condiciones de regularidad, valores cercanos a 0.5 muestran tendencia a la aleatoriedad y los próximos a 1 presentan condiciones de agrupamiento (Gadow, Hui y Albert, 1998):
Donde:
Wi = valor del índice para el j-ésimo árbol referencia
n = es el número de árboles vecinos considerados
Vij = variable 1 cuando el j-ésimo ángulo α entre dos árboles vecinos próximos es menor o igual al ángulo estándar α, en caso contrario toma un valor de 0.
Grado de mezcla en la distribución espacial de especies (M i )
Se calculó con base en el índice de mezcla de especies (M i ) el cual es una medida de la diversidad en la distribución espacial de especies mostrando el grado de mezcla entre estas, Füldner (1995) define este índice como la proporción de n vecinos más cercanos que no pertenecen a la misma especie que el árbol de referencia, en este caso se toman los cuatro vecinos más cercanos.1
Donde 0 ≤ M i ≤ 1, vj es igual a 0 cuando el árbol j es de la misma especie que el árbol de referencia i y será 1 en caso contrario (Gadow et al., 2007).
Dominancia dimensional (U i )
Se define como la dominancia del árbol referencia i con respecto a sus vecinos más cercanos, es decir, la proporción de árboles n más pequeños (en diámetro o altura) que el árbol de referencia.
Donde 0 ≤ U i ≤ 1 y vj será igual a 1 si el árbol j es menor al árbol referencia i, y 0 en caso contrario. Con cuatro vecinos el índice de dominancia Ui puede tomar cinco valores y es muy útil cuando se desea ver la dominancia relativa de una especie en particular (Aguirre, Hui, von Gadow y Jiménez, 2003; Gadow et al., 2007).
Diferenciación dimensional (T i )
Se puede aplicar a cualquier variable representativa de las dimensiones del arbolado, en este caso para diámetros y alturas. El grado de diferenciación dimensional establece una manera de representación de la proximidad espacial que existe entre árboles de diferentes dimensiones en el bosque. Toma en cuenta los n árboles más cercanos al árbol de referencia i y se define de la siguiente manera:
Donde:
Ti = grado de diferenciación en diámetros o/y alturas
j = 1. . . n árboles vecinos
i = árbol de referencia
D imi = diámetro o altura del árbol i
D imj = diámetro o altura del árbol j
El valor del T i se incrementa al aumentar la diferencia media del tamaño de los árboles vecinos (0 ≤ T i ≤ 1). Entre más bajo sea el valor de T i las diferencias son menores, un valor de 0 indica que todos los árboles son del mismo tamaño (Gadow et al., 2007). Siguiendo el método de Aguirre, Kramer y Jiménez (1998), el grado de diferenciación en diámetros y alturas se clasificó en cinco categorías, 0-0.2 = débil; 0.2-0.4 = moderada; 0.4-0.6 clara; 0.6-0.8 fuerte; 0.8-1.0 muy fuerte.
El cálculo de los índices se ejecuta dentro de una ventana de observación (W), que esta vez son los sitios circulares de muestreo. En dichos sitios se realiza cada uno de los análisis de la información con las fórmulas previamente descritas. Considerándose el efecto de borde de la ventana (Pommerening y Stoyan, 2006), este efecto desempeña un papel importante para la correcta caracterización de la estructura espacial.
Para una correcta interpretación de las características estructurales en el espacio se debe tomar en cuenta el efecto de borde, ya que hay árboles dentro de los sitios de muestreo que se encuentran cerca del borde W y normalmente tienen a sus vecinos más cercanos del otro lado. Lo anterior implica errores en la estimación de los valores de vecindad de los árboles que se encuentran dentro. Con la finalidad de evitar errores se utilizó el estimador NN1 el cual consiste en omitir como árbol de referencia (i) a aquellos cuya distancia a su j-ésimo vecino más cercano sea menor a la distancia entre este y el borde de W, pese a esto pueden formar parte de otros grupos estructurales como vecino más cercano (Pommerening y Stoyan, 2006).
Con el objetivo de comparar estadísticamente los índices antes mencionados con bosques perturbados o bajo manejo, se amplió la base de datos a 1000 submuestras utilizando la paquetería “WRS2” en el programa “R” (R Core Team, 2019), mediante la cual se usaron simulaciones “Bootstrap” para calcular la diferencia entre medias con un IC de 95% (Mair, Wilcox y Schoenbrodt, 2015; Wilcox, 2011).
Resultados
Los resultados de distribución espacial se muestran para el bosque en general y por géneros (Pseudotsuga, Quercus y Pinus), por ser los más abundantes, de mayores dimensiones y más investigados en este aspecto. La densidad total fue de ha-1 con un área basal de 10.14 m2 ha-1 siendo Pseudotsuga el género más importante (Tabla 1).
Tabla 1 Características dasométricas del área de estudio.
| Género | Densidad (ha -1 ) | G (m 2 ha -1 ) | d1.30 (cm) | h (m) |
| Pseudotsuga | 105 | 3.4780 | 21.24 | 11.04 |
| Quercus | 28 | 1.7196 | 21.45 | 10.94 |
| Pinus | 53 | 1.6963 | 21.30 | 10.99 |
| Arbutus | 34 | 2.2618 | 21.29 | 10.96 |
| Juniperus | 16 | 0.5176 | 21.27 | 10.82 |
| Abies | 13 | 0.2504 | 21.71 | 10.92 |
| Picea | 6 | 0.2194 | 21.87 | 10.72 |
| Total | 254 | 10.143 |
G = área basal, d1.30 = diámetro normal promedio, h = altura promedio.
Distribución espacial del arbolado
En la Figura 2 se muestra la distribución espacial del arbolado en cada uno de los sitios después de usar el estimador NN1, es decir, únicamente se muestran los árboles que pueden ser el árbol i del grupo estructural.

Como resultado, en el análisis de los índices de vecindad se utilizaron un total de 203 grupos estructurales de los cinco árboles para el bosque en general, siendo los sitios 2 y 6 donde se conformó una mayor cantidad de grupos, 41 y 54 respectivamente; Pseudotsuga generó 84 grupos estructurales, Quercus 22 y Pinus 42.
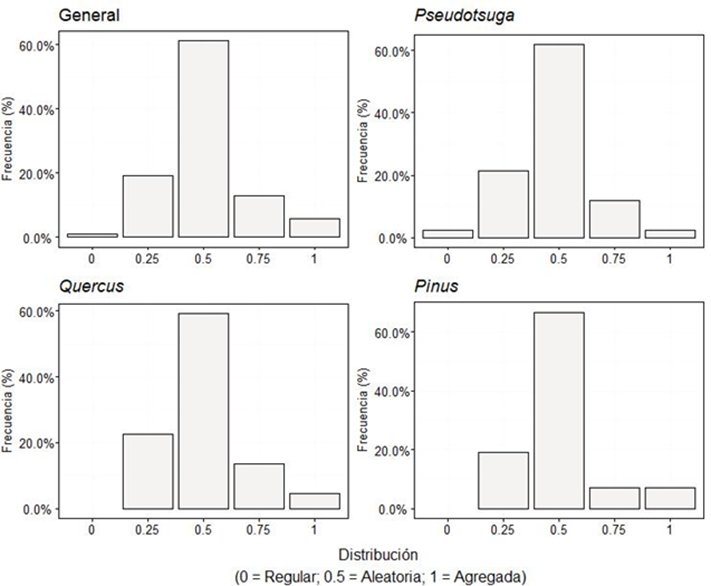
Uniformidad de ángulos (W i )
En el análisis general, los resultados muestran una media (
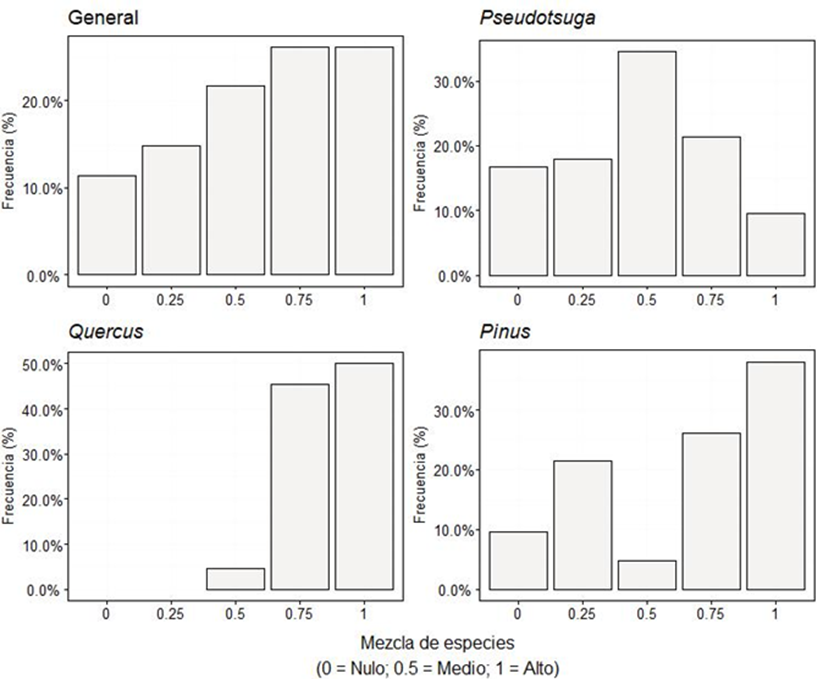
Índice de mezcla de especies (M i )
Se obtuvo (
Índice de dominancia dimensional diamétrica (UD i )
Las distribuciones de los valores de dominancia relativa no muestran ninguna tendencia en el análisis general del bosque (Fig. 5) (
Índice de dominancia dimensional de altura (UH i )
De manera general (

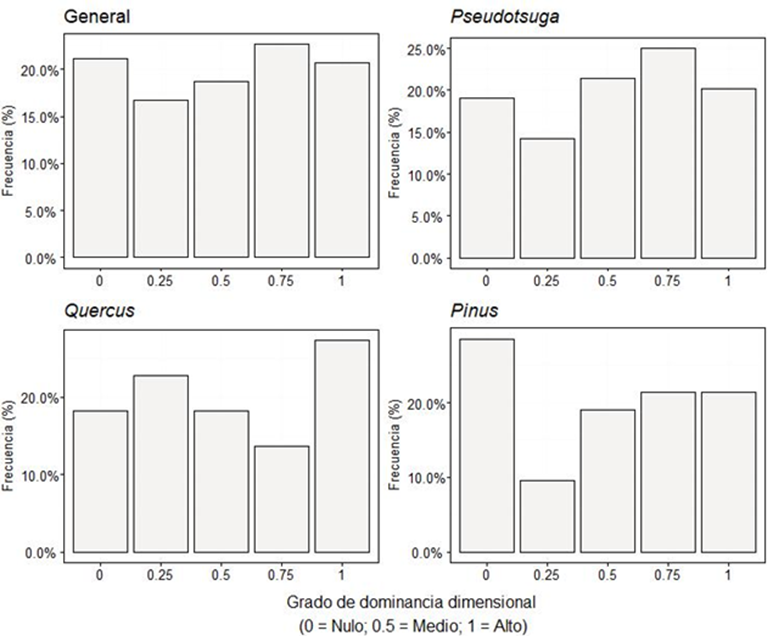
Índice de diferenciación dimensional diamétrica (TD i )
El bosque muestra una diferenciación diamétrica moderada, teniendo alrededor de 60% de frecuencia en esta categoría (Fig. 7), seguida de la categoría clara. El análisis por género mostró la misma tendencia (Moderada). Por otra parte, los valores medios obtenidos mediante la prueba Bootstrap quedaron dentro del intervalo de confianza para todos los grupos.

Índice de diferenciación dimensional de altura (TH i )
Al igual que en la diferenciación diamétrica la distribución de alturas se muestra similar presentando la mayoría de sus frecuencias en la categoría moderada, seguida de la clara (Fig. 8). El análisis por género mostró la misma tendencia y los valores medios obtenidos mediante la prueba Bootstrap quedaron dentro del intervalo de confianza para todos los grupos.

Discusión
Respondiendo a las preguntas de investigación planteadas la distribución y mezcla del arbolado fue aleatoria, asimismo el bosque presentó una tendencia a la homogeneidad ya que la diferenciación dimensional fue de moderada a clara. Una de las principales similitudes con otros estudios se reflejó en el índice de Uniformidad (Wi) aplicado también en bosques naturales, en bosques bajo manejo y en bosques con antecedentes de incendios (Graciano-Ávila, Alanís-Rodríguez, Rubio-Camacho, Valdecantos-Dema, Aguirre-Calderón, González-Tagle y Mora-Olivo, 2020; Rubio-Camacho et al., 2017; Zhang et al., 2019). Mientras que una de las diferencias se presentó en el índice de mezcla de especies (Mi) el cual fue distinto a lo encontrado en bosques de pino - encino y a otras poblaciones de Pseudotsuga (Domínguez-Calleros et al., 2014; Rubio-Camacho et al., 2017).
La estructura espacial del bosque es el resultado de dinámicas pasadas e interacciones complejas entre muchos procesos (Marie-Agnès y Audrey, 2003). Esta dinámica incluye la heterogeneidad ambiental y la competencia entre individuos y poblaciones (Duncan, 1991; Valverde y Silvertown, 1997). Aguirre et al. (2003) mencionan que este tipo de descripciones son más completas que los enfoques tradicionales.
Distribución espacial del arbolado
Los grupos estructurales generados (254 ha-1) para este tipo de bosque fueron superiores a los creados por Graciano-Ávila, Alanís-Rodríguez, Aguirre-Calderón, González-Tagle, Treviño-Garza y Mora-Olivo, (2019) para una parcela con tratamiento de regeneración (96 ha-1) y otra con tratamiento de selección (134 ha-1) para un bosque de pino-encino, mientras que Rubio-Camacho et al. (2017) generaron 243 ha-1 grupos para una zona con evidencias de incendio y 281 ha-1 para una sin evidencias de incendios en un bosque pino-encino. La mayoría de las especies arbóreas en los bosques de todo el mundo se agregan espacialmente a diferentes escalas y los patrones espaciales de los árboles cambian con la etapa de desarrollo del bosque, particularmente en los bosques naturales no muy perturbados por acción antropogénica (Condit, Ashton, Baker, Bunyavejchewin, Gunatilleke, Gunatilleke y Yamakura, 2000; Getzin et al., 2008; Yang et al., 2018).
Índice de uniformidad de ángulos (W i )
Dicho índice evidenció una distribución aleatoria del bosque. Los resultados fueron similares a los calculados por Zhang et al. (2019), quienes evaluaron bosques naturales de Picea schrenkiana, Pinus koraiensis y un bosque mixto de pino - roble en diferentes regiones de China. Rubio-Camacho et al. (2017) registraron también esta distribución en bosques regenerados después de los incendios forestales. Mientras que Graciano-Ávila et al., (2020) lo documentaron para bosques bajo aprovechamiento forestal.
Los resultados de Quercus y Pinus no presentaron diferencias significativas (p ≥ 0.05) contra estudios en bosques similares (Rubio-Camacho et al., 2017), ya que sus valores están entre 0.47 y 0.50 quedando dentro del intervalo de confianza (IC) de 95%, Pseudotsuga por su parte no presentó diferencias con los valores registrados (S1: 0,509 y S2: 0,532) por Domínguez-Calleros et al. (2014).
Índice de mezcla de especies (M i )
De manera general indica que los árboles referencia (i) están rodeados por individuos de especies diferentes a él. El análisis por género revela que Pseudotsuga no muestra preferencia por estar o no mezclado, contrario a lo publicado por Domínguez-Calleros et al. (2014), quienes observaron una mezcla alta para dos poblaciones de Pseudotsuga menziesii en la reserva de la biosfera “La Michilía” en los estados de Durango y Zacatecas. Quercus y Pinus muestran un mayor grado de mezcla en este estudio que en bosques de pino - encino (Rubio-Camacho et al., 2017).
Los resultados generales para mezcla de especies presentaron diferencias significativas comparado con lo mencionado por Rubio-Camacho et al. (2017), ya que sus valores medios fueron P1: 0.57 y P2: 0.58. Lo anterior se corrobora en el análisis por género ya que los mismos autores mencionan valores de P1: 0.49 y P2: 0.56 para Pinus y Quercus respectivamente quedando fuera del IC calculado (Pinus: 0.83-0.97, Quercus: 0.80-0.98). Pseudotsuga de igual manera presentó diferencias estadísticamente significativas (p ≥ 0.05) con lo observado (S1: 0,732 y S2: 0,665) por Domínguez-Calleros et al. (2014).
Índice de dominancia dimensional diamétrica (UD i )
No mostró ninguna tendencia para el bosque en general. Estos resultados al igual que los de Graciano-Ávila et al., (2019) y Rubio-Camacho et al. (2017) determinan que el bosque es heterogéneo en este aspecto, sin embargo, el análisis por género muestra que Pseudotsuga es dominante sobre las demás especies ya que este género es de los más abundantes y de mayores dimensiones (Diámetro normal, cobertura de copa) en el área de estudio (García et al., 2019). Quercus mostró una alta dominancia sobre las demás especies, aunque tiene un gran porcentaje en las categorías menores, Pinus por su parte se comportó como especie dominada contrario a lo observado por Rubio-Camacho et al. (2017).
Índice de dominancia dimensional de altura (UH i )
Manifestó que Pseudotsuga es el género dominante en el área de estudio lo cual se entiende, ya que, de acuerdo con García et al., (2019) sus individuos son abundantes y de alturas máximas de hasta 26.30 m. Al igual que en el estudio de Rubio-Camacho et al. (2017)Quercus es dominado por los demás géneros, mientras que Pinus es codominante según la clasificación de Gadow et al. (2007). Los resultados generales mostraron diferencias significativas (p ≤ 0.05) contra la parcela 1 estudiada por Rubio-Camacho et al. (2017), quienes obtuvieron una media de 0.35, mientras que para la parcela 2 no existieron diferencias donde el valor fue de 0.45 encontrándose dentro del IC (0.43-0.59).
Índice de diferenciación dimensional diamétrica y en altura (TD i y TH i )
El grado de diferenciación dimensional tanto en diámetros como en alturas fue moderado, similar a lo encontrado por Rubio-Camacho et al. (2017) y Graciano-Ávila et al. (2019) en bosques de pino - encino. Esto indica que existe escasa variabilidad en las dimensiones del arbolado y que la estructura, tanto horizontal como vertical es poco heterogénea.
Conclusiones
Los resultados de esta investigación indican que la uniformidad (Wi), en este caso aleatoria, no es exclusiva de bosques sin disturbio o sin manejo, además se observa que la mezcla de especies (Mi) no necesariamente es mayor en bosques maduros y sin intervención como en este caso. Por su parte la dominancia diamétrica (UDi) muestra que géneros como Pinus pueden llegar a ser dominados por otros, así como la dominancia en altura (UHi) corrobora que Quercus es un género dominado. Por su parte la diferenciación dimensional, en otros tipos de bosques es homogénea. Lo obtenido en este estudio deja en claro que existe necesidad de generar información de la estructura y dinámica espacial, de este tipo de ecosistemas ya que es escasa. La aplicación de los índices aquí empleados contribuye de manera importante al entendimiento de los procesos que se llevan a cabo en un bosque para llegar a generar una estructura. Lo anterior da bases para el manejo y conservación de comunidades vulnerables como la aquí estudiada, teniendo conocimiento de manera más individualizada de la dinámica a nivel especie o género.

