











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
El incremento de temperatura global a consecuencia de las emisiones de gases de efecto invernadero hacia la atmós- fera, es cada vez más alto, se espera que para el siglo XXI sea de entre 1.7 °C y 4.8 °C (IPCC, 2014). Algunos autores predicen que para el 2030 la temperatura será de 1.5 °C más (Sáenz-Romero et al., 2010) y se estima que para que no se rebase este umbral, las emisiones deberían reducirse en al menos 25% para ese año (Allen et al., 2018).
Con el aumento de temperatura, se modifica el sistema climático afectando tanto a la población humana como a la biodiversidad de flora y la fauna de todo el mundo; entre los problemas más notables están la migración de especies vegetales hacia nuevas áreas (Sáenz-Romero et al., 2010; Allen, 2018), otras especies modificarán su intervalo altitu- dinal (Lenoir, Gégout, Marquet, De Ruffray y Brisse, 2008) mientras que otras, aumentarán su área de distribución (Cruz-Cárdenas, López, Silva, Bernal, Estrada y López, 2016).
Se ha demostrado a través el diagrama «BAM» que la distribución de las especies depende de varios factores (Soberón y Nakamura, 2009): i) deben existir las condi- ciones abióticas necesarias («A»), ii) condiciones bióticas favorables («B») y iii) hayan estado accesibles a la dispersión de la especie desde la región de su origen histórico («M»). Actualmente existen diferentes algoritmos para modelar la distribución de especies destacando BIOCLIM (Busby, 1991), GARP (Stockwell y Peters, 1999) y Maxent (Phillips, Dudík y Schapire, 2004), etc., considerado este último como uno de los mejores predictores de la idoneidad de las especies (Elith et al., 2006; Phillips, Anderson y Schapire, 2006).
Los bosques contribuyen significativamente a la mitigación del cambio climático al almacenar el CO2 atmosférico a través del proceso de la fotosíntesis (Alberto, 2010), en México estos cubren aproximadamente 17% del territorio nacional, equivalente a 10 millones de hectáreas (Comisión Nacional para el Conocimiento y Uso de la Biodiversidad [Conabio], 2008), sin embargo, las especies de estos ecosistemas también son susceptibles al cambio climático (Cruz-Cárdenas et al., 2016). Pinus engelmannii Carriére, una especie de gran importancia comercial en el país (Bustamante-García, Prieto-Ruíz, Merlín-Bermudes, Álvarez-Zagoya, Carrillo-Parra y Hernández-Díaz, 2012), es característica de las zonas montañosas de México y se distribuye en la Sierra Madre Occidental (SMOc) (Martínez, 1948) y en algunas poblaciones aisladas al sureste de Arizona y Nuevo México (Farjón, Pérez y Styles, 1997).
Los modelos de nicho ecológico son una herramienta importante para evaluar la respuesta de las especies al cambio climático (Soberón y Nakamura, 2009; Sáenz- Romero et al., 2010). Así mismo, se han optimizado los parámetros de los algoritmos de modelación y con ello, las predicciones son cada vez mejor (Cobos, Townsend, Barve y Osorio, 2019), se están actualizando los modelos de circulación general (MCGs) (Karger et al., 2017) permitiendo predecir la distribución futura de las especies bajo diferentes niveles de concentración de CO2 (Cruz- Cárdenas et al., 2016), no obstante, la distribución futura de las especies a causa del incremento de temperatura por el cambio en el clima, es incierta y debe ser cuantificada a nivel especie.
Objetivos
i) Modelar la distribución actual y potencial de P. engelmannii Carriére, usando variables bioclimáticas, cuatro MCGs: HadGEM2-AO, CSIRO-Mk3-6-0, CCSM4 y CNRM-CM5, en dos trayectorias de concentración representativas (RCPs) 4.5 y 8.5 al año 2050; ii) conocer las variables climáticas que mayor influencia poseen sobre la distribu- ción de la especie; y iii) determinar nuevas áreas idóneas y áreas de alto impacto (áreas idóneas más vulnerables al cambio climático) de P. engelmannii. La hipótesis es que las áreas idóneas actuales de P. engelmannii podrían ser afectadas en los próximos años, debido al cambio climático.
Materiales y métodos
Área de estudio y especie
El estudio consideró el área de distribución de P. engelmannii, la cual se sitúa en la Sierra Madre Occidental; ahí, las temperaturas promedio varían entre 13 °C y 28 °C (García, 1998) y la precipitación varía entre 500 mm y 800 mm anuales (Descroix, González y Estrada, 2004). En México esta especie se encuentra en altitudes que van desde 1200 m a 3000 m s.n.m (Martínez, 1948, Farjón et al., 1997) y puede desarrollarse en suelos profundos (Perry, 1991; González-Elizondo, González-Elizondo, Tena-Flores, Ruacho-González y López- Enríquez, 2012). Se le conoce como ‘pino real’ (Martínez, 1948) y es la única especie en la SMOc con fascículos muy largos y hojas muy gruesas (Farjón et al., 1997).
Variables bioclimáticas y modelos de circulación general
Para modelar la distribución actual de la especie se utilizaron 15 variables bioclimáticas (Tabla 1), obtenidas de: http://chelsa-climate.org/downloads (Climatologies at High resolution for the Earth´s Land Surface Areas), son cubiertas geográficas del periodo actual (1979-2013) en formato raster de 1 km2 de resolución (Karger et al., 2017) y re muestreadas a 5 km2 para este análisis; estos autores han demostrado que con estas variables se predice mejor que modelar con variables de WorldClim. Para modelar la distribución futura (2050), se utilizaron las mismas variables bioclimáticas, de cuatro modelos de circulación general: HadGEM2-AO (Martin et al., 2011), CSIRO-MK3-6-0 (Collier et al., 2011), CCSM4 (Gent et al., 2011) y CNRM- CM5 (Voldoire et al., 2013), utilizando los RCPs 4.5 y 8.5, para el periodo 2041-2061. Aquí, se omitieron las variables Bio 8, Bio 9, Bio 18 y Bio 19 por combinar información de precipitación y temperatura en la misma capa (Escobar, Lira-Noriega, Medina-Vogel y Townsend, 2014), por lo que su uso en modelos de nicho trae consigo predicciones erróneas.
TABLA 1 Variables bioclimáticas empleadas para generar modelos de nicho ecológico para modelar la distribución actual y potencial de Pinus engelmannii Carriére.
| Variable | Descripción |
| Bio 1 | Temperatura media anual (°C) |
| Bio 2 | Intervalo medio diurno (Media por mes [Temp. Max -Temp. Min]) |
| Bio 3 | Isotermalidad (Bio2/Bio7) (* 100) |
| Bio 4 | Temporalidad de la temperatura (desviación) |
| Bio 5 | Temperatura máxima del mes más cálido (°C) |
| Bio 6 | Temperatura mínima del mes más frío (°C) |
| Bio 7 | Intervalo de temperatura anual (Bio5-Bio6) |
| Bio 10 | Temperatura media del 1/4 más cálido (°C) |
| Bio 11 | Temperatura media del 1/4 más frío (°C) |
| Bio 12 | Precipitación anual (mm) |
| Bio 13 | Precipitación del mes más húmedo (mm) |
| Bio 14 | Precipitación del mes más seco (mm) |
| Bio 15 | Temporalidad de la precipitación (Coeficiente de variación, %) |
| Bio 16 | Precipitación del 1/4 anual más húmedo (mm) |
| Bio 17 | Precipitación del 1/4 anual más seco (mm) |
Obtención de registros y limpieza
Los registros de presencia de la especie fueron obtenidos de Global Biodiversity Information Facility (GBIF), Inventario Nacional Forestal y de Suelos (INFyS, 2009), Naturalista, Artículos Científicos y Tesis y de la Red Mundial de Información Sobre Biodiversidad (REMIB), Conabio, estos fueron depurados, eliminando duplicados, fuera del área de distribución en altitud, latitud y longitud, siguiendo la propuesta por algunos autores (Méndez, Méndez, Mendieta, López y Nájera, 2021). Registros atípicos o ‘outliers’ fueron identificados y eliminados realizando el espacio ambiental con Bio 1 y Bio 12 (Owens et al., 2013) y a través de Análisis de Componentes Principales (ACP), usando 15 variables bioclimáticas (estandarizadas), descartando aquellos fuera de la elipse de 99%, para después eliminar otros registros por autocorre- lación a través del paquete SpThin aplicando una distancia de 5 km (Aiello-Lammens, Boria, Radosavljevic, Vilela y Anderson, 2015).
Área de calibración (M)
La delimitación del área de estudio, «M» del diagrama BAM es importante para la calibración del modelo y en lo suce- sivo, para transferirlo a nuevas áreas y tiempos (Soberón, Osorio-Olvera y Peterson, 2017), esta se generó aplicando un búfer de 70 km de radio a cada registro y su delineación final fue después de pasar el proceso de depuración de los registros y eliminación de ‘outliers’, incluyendo todos los registros, antes del proceso de autocorrelación para captar toda la variabilidad climática de la especie.
Selección de variables bioclimáticas
La selección de variables predictivas, se basó en los siguientes criterios: 1) las de mayor contribución acorde a la prueba de jackknife, haciendo una modelación preliminar (Ávila, Villavicencio y Ruiz, 2014; García-Aranda, Méndez- González y Hernández-Arizmendi, 2018; Aceves-Rangel, Méndez, García y Nájera et al., 2018), 2) las de mayor contribución relativa obtenida de un ACP realizado según lo propuesto por Méndez et al. (2021), 3) de acuerdo con la biología de la especie, 4) no correlacionadas (r ≤ 0.8) (Elith et al., 2006; Cruz-Cárdenas, Villaseñor, López-Mata, Martínez-Meyer y Ortiz, 2014; Martínez-Méndez Aguirre- Planter, Eguiarte y Jaramillo-Correa, 2016), 5) que tuvieran una distribución normal o sesgada (Méndez et al., 2021) y 6) por el tipo de respuesta de la variable bioclimática. De esta selección preliminar, se organizaron tres grupos (sets) con diferentes variables bioclimáticas, utilizados para calibrar el modelo para predecir la distribución de P. engelmannii (Cobos et al., 2019).
Calibración, generación, validación y transferencia La calibración, creación, evaluación y transferencia del modelo se realizó en R, utilizando el paquete ‘kuenm’ (Cobos et al., 2019). Se evaluaron 21 multiplicadores de regularización, 29 tipos de respuesta y tres grupos de variables ambientales, en tres modalidades: extrapolación libre (EL), extrapolación clamping (EC) y no extrapolación (NE). El formato de salida fue de tipo logístico, donde los modelos resultantes (mapas en formato raster) representan valores de idoneidad (0-1) de la especie (Phillips et al., 2006). Los modelos generados fueron evaluados a través de su significancia estadística, ROC parcial (curvas operadas por el receptor), tasa de omisión, criterio de información Aikaike o complejidad del modelo (AICc) (Phillips et al., 2006) y pruebas jackknife (Cobos et al., 2019). Se empleó ROC parcial para determinar el ajuste del modelo, descartando el estadístico AUC (Area Under Curve) ya que no predice en todo el espectro de área predicha presente de la curva ROC (Peterson, Papeş y Soberón, 2008). El riesgo de extrapolación (transferencia) se evaluó a través de MOP (Mobility- Oriented Parity) que identifica zonas de extrapolación estricta (Owens et al., 2013).
Los mapas de idoneidad actual se estratificaron en alta, media y baja, utilizando cuantiles relativos (Instituto Nacional de Estadística Geografía en Informática [Inegi], 2010). Los modelos al futuro se promediaron y binarizaron al umbral de 10 percentil y a través de álgebra de rasters se identificaron: i) nuevas áreas idóneas, NA (áreas idóneas en el futuro, pero que no existen actualmente), ii) áreas de alto impacto, AAI (áreas idóneas que existen actualmente pero que podrían no serlo en el futuro), y iii) áreas de bajo impacto, ABI (áreas idóneas que existen actualmente y en el futuro). Todo este proceso metodológico, desde la descarga de los registros hasta la validación de la transferencia del modelo, se ilustra en la figura 1.

Resultados
El modelo de nicho de Pinus engelmannii Carriére De un total de 507 registros de P. engelmannii, resultaron útiles solo 346 (68.2%), 241 para modelación y 105 (independientes) para evaluación. Se generaron 1827 modelos candidatos, el modelo seleccionado fue de un regularizador de 3.5, quadratic + threshold (qt), con ROC parcial de 1.3, tasa de omisión de 0.05% y AICc de 3107. Las variables que mejor predicen la distribución actual de P. engelmannii son: Bio 1 [temperatura media anual (ºC)] y Bio 17 [precipitación de 1/4 anual más seco (mm)], contribuyendo al modelo con 75.6% y 6.3%. El resto, Bio 16, 15 y 5 representaron solo 18.1% de la contribución.
Según el espacio ambiental construido con Bio 5, 6 y 12 (Fig. 2), los registros de P. engelmannii se distribuyen de 20 °C a 37 °C (Bio 5, temperatura máxima), de -3.6 °C a 15 °C (Bio 6, temperatura mínima) y de 313 mm a 1682 mm (Bio 12, precipitación); el elipsoide (nicho fundamental) se inclina ligeramente sobre Bio 6, revelando que la especie prefiere climas semisecos y fríos (Fig. 2), ocupando una pequeña porción del espacio ambiental y pocos de los múltiples climas existentes en el país (círculos verdes).
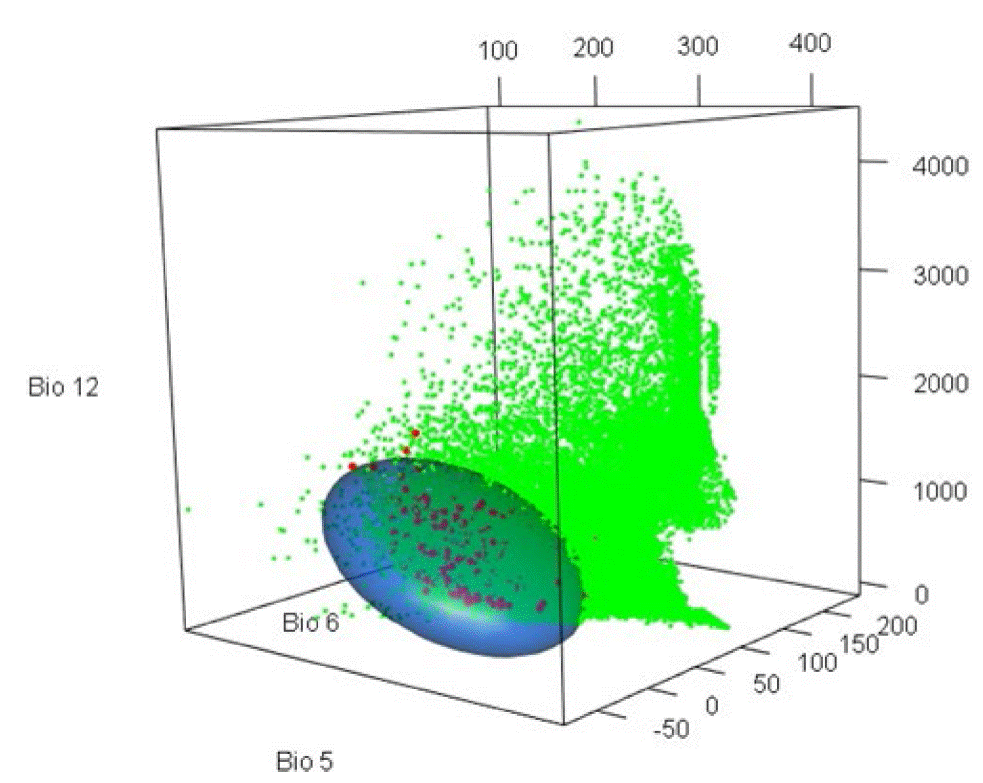
FIGURA 2 Espacio ambiental indicando el nicho fundamental (elipsoide al 99%.) de Pinus engelmannii Carriére, elaborado con Bio 5, Bio 6 y Bio 12, utilizando 346 registros y 50 000 puntos de background distribuidos en todo México. Puntos rojos representan los registros de la especie y puntos verdes indican valores de las Bios; es decir, los climas del país).
Distribución actual de Pinus engelmannii Carriére En el área de calibración «M», los tres tipos de modelación (EL, EC y NE) predicen idoneidad similar de P. engelmannii, solo se presenta la modalidad NE (Fig. 3a). La incertidumbre del modelo representada aquí por el Coeficiente de Variación, es baja en las áreas de alta idoneidad (Fig. 3b). La idoneidad alta de P. engelmannii ocurre de forma discontinua en Chihuahua (29 090 km2), Durango (9310 km2) y Sonora (2080 km2) y representa 12.82% total del «M», esta ‘discontinuidad’ podría indicar una fragmentación del hábitat más que una posible dispersión (migración) de la especie hacia nuevas áreas (Saltré, Duputié, Gaucherel y Chuine, 2014) por efectos del aprovechamiento inadecuado o del cambio climático. El estrato con idoneidad media (51 080 km2) constituye 15.75% de «M» y el bajo (231 570 km2) 71.42%.
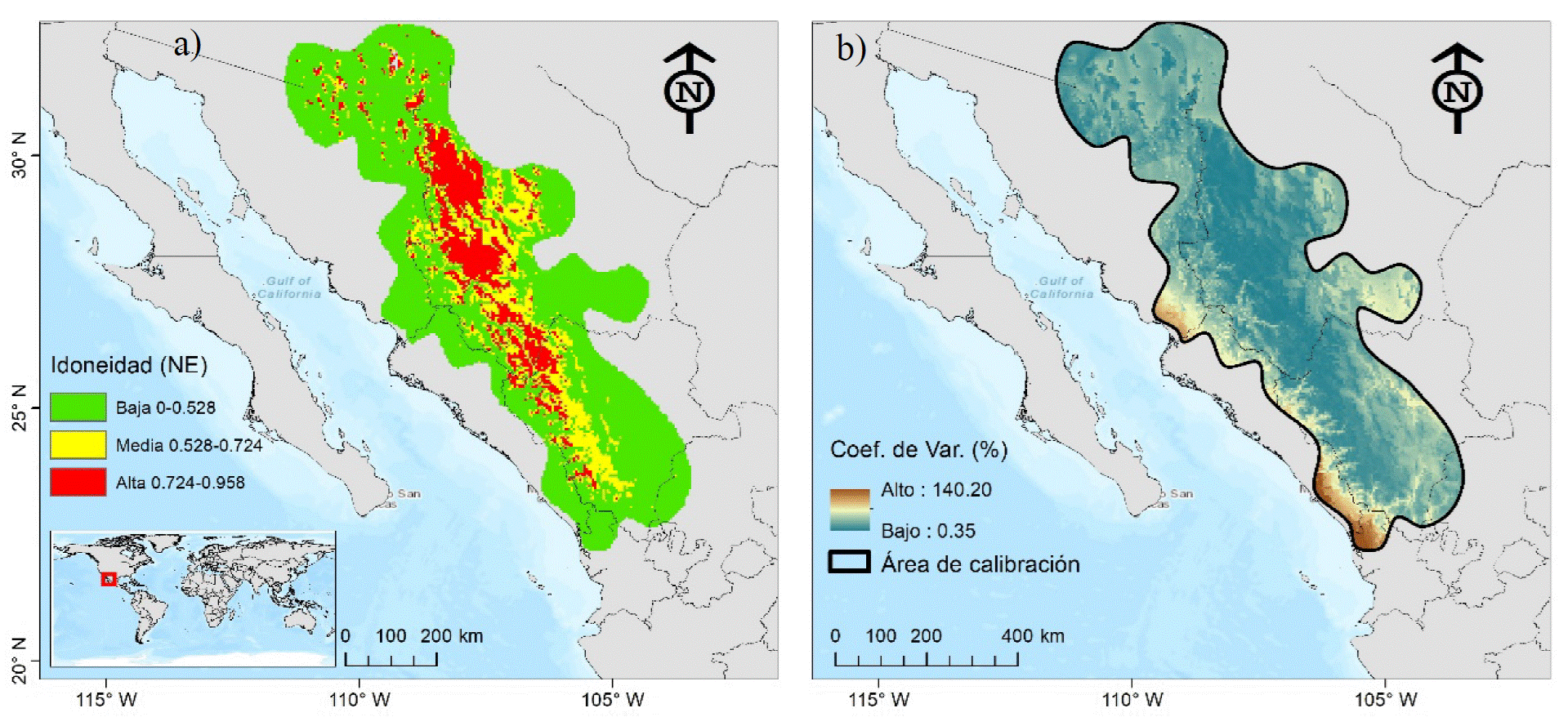
FIGURA 3 Modelo de idoneidad actual de Pinus engelmannii Carriére, modalidad sin extrapolación, estratificada en alta, media y baja (a), e incertidumbre del modelo, expresada a través del coeficiente de variación (b).
Transferencia del modelo y el riesgo de extrapolación
El análisis de resultados demostró que al trasferir el modelo de nicho de P. engelmannii al 2050, las áreas idóneas predichas para la especie (Fig. 4 b-4c), son similares en extensión a la distribución actual (Fig. 4a), independientemente, si el escenario de emisiones de CO2 es bajo, RCP 4.5 (Fig. 4b) o alto, RCP 8.5 (Fig. 4c). Sin embargo; no es posible sugerir las áreas predichas si el riesgo de trasferencia del modelo no es evaluado (Owens et al., 2013). Por ejemplo, el modelo (elegido de entre 1827 candidatos por su mejor desempeño) predice una extrapolación irrazonable de áreas de idoneidad al modelar con EL y EC (resultados no presentados) que al no extrapolar (Fig. 4a-4c), por lo que es crucial probar las diferentes modalidades de trasferencia de un modelo para elegir la adecuada.

FIGURA 4 Modelo de distribución potencial actual (a) y futuro al 2050 en el RCP 4.5 (b) y RCP 8.5 (c) para Pinus engelmannii Carriére, predicha en la modalidad no extrapolación. Riesgo de extrapolación (MOP) de la transferencia del modelo, estimado para el RCP 4.5 (d) y RCP 8.5 (e).
En este trabajo se eligió la modalidad no extrapolación (Fig. 4a-4c), por presentar predicciones razonables. El análisis del riesgo de transferencia del modelo indicó similaridad ambiental entre las áreas idóneas actuales predichas dentro del área de calibración y las áreas idóneas estimadas al 2050 (Fig. 4d-4f). Las grandes regiones idóneas predichas en las modalidades EL y EC (mapas no mostrados aquí), el MOP las clasificó a como extrapolación estricta; es decir, no todas las áreas idóneas predichas por un modelo de nicho son adecuadas para la especie.
Distribución potencial de Pinus engelmannii Carriére Una vez validada la transferencia del modelo, se observa que dentro del área de calibración (Fig. 5), independientemente de si las emisiones de CO2 al 2050 sean altas (RCP 8.5) o bajas (RCP 4.5) no se predicen nuevas áreas idóneas para la especie, pero tampoco se pierden las áreas idóneas existentes (Fig. 5a y 5b). Fuera de «M», (pero dentro del país) el modelo de nicho predice, pequeñas nuevas áreas idóneas en Nuevo León, Puebla, Tlaxcala y Ciudad de México (Fig. 5a y 5b).
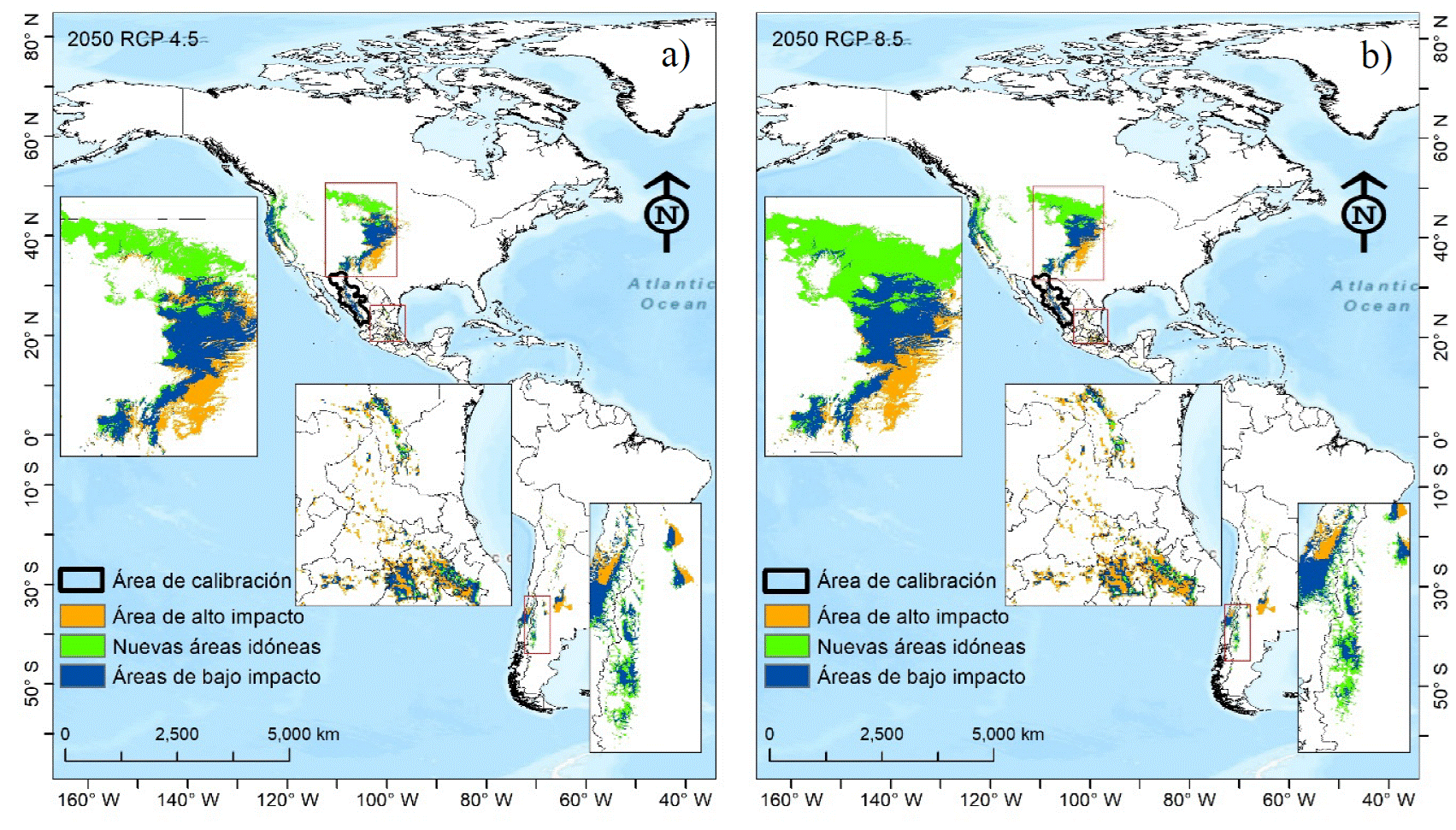
FIGURA 5 Área de idoneidad actual (color azul y naranja) y potencial (color verde) de Pinus engelmannii Carriére, promedio de cuatro modelos de circulación general (MCG´s) en dos trayectorias de concentración representativas RCP´s 4.5 (a) y 8.5 (b).
Es notable que las NAI más extensas se registran al centro-norte de Estados Unidos (Fig. 5a y 5b) y algunas otras en Sudamérica, en Argentina y Chile. Usando los valores de la climatología actual (1979-2013), según la prueba no paramétrica de Kruskal-Walis existe alta probabilidad (99%) de una tendencia clara de que las NAI (Fig. 5b y 5b) serán aquellas más frías, mientras que las AAI (áreas que dejarían de ser idóneas para la especie) es donde se espera un incremento significativo de temperatura al 2050 (Fig. 6).
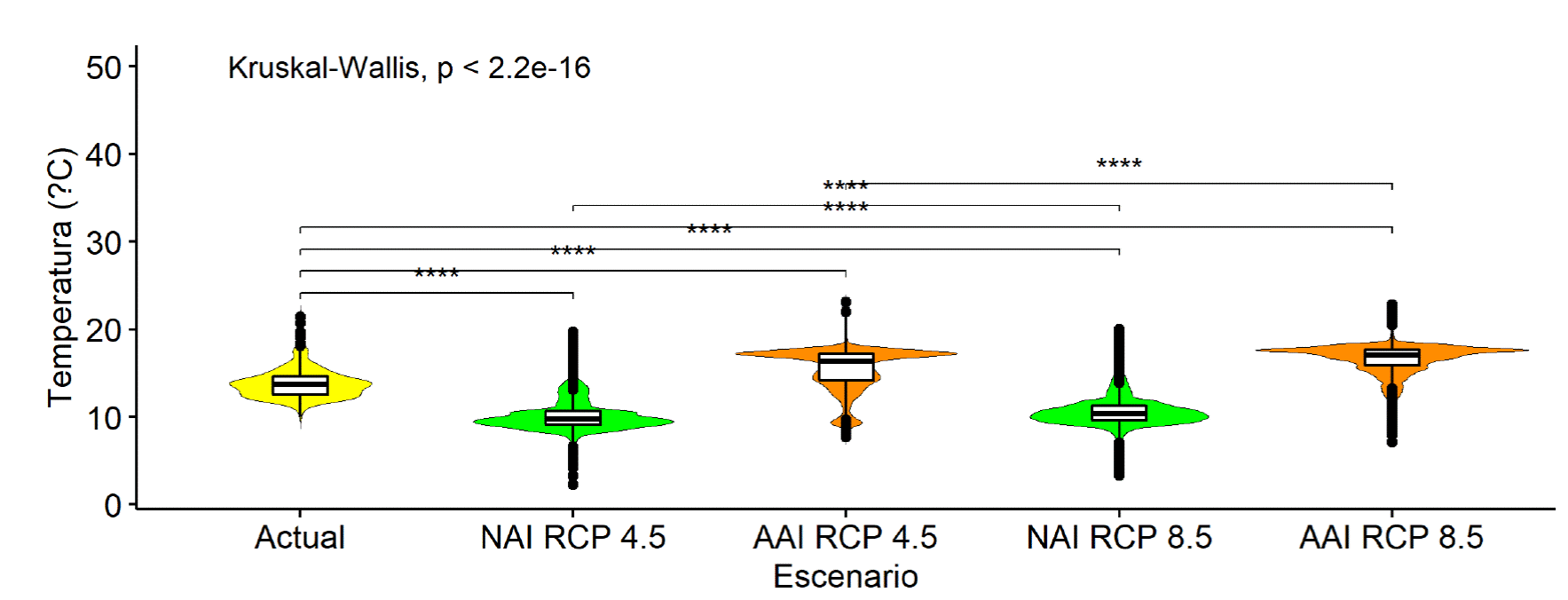
FIGURA 6 Temperatura media anual (Bio 1) actual, registrada en nuevas áreas idóneas (NAI) y en áreas de alto impacto (AAI), predichas al 2050, promedio de cuatro modelos de circulación general (MCG´s) en dos RCP´s (4.5 y 8.5) para Pinus engelmannii Carriére.
Perfil bioclimático de Pinus engelmannii Carriére Según los registros de ocurrencia, esta especie actualmente se registra entre 9.8 °C y 25.7 °C (Bio1, Tabla 2), preferentemente entre 12 °C y 16 °C (Fig. 7a); y desde 7 mm hasta 72 mm (Bio 17, Tabla 2, Fig. 7b). Según las proyecciones de los MCGs al 2050, donde hoy ocurre la especie, en general se espera que en promedio la temperatura sea 1.6 °C y 1.9 °C más (RCP´s 4.5 y 8.5), mientras que la precipitación disminuya entre 5 mm y 7 mm (Tabla 2). Aunque el intervalo altitudinal de la especie es muy amplio, sus preferencias son desde 2600 m a 1600 ms.n.m. (Fig. 7c). Bio 1 muestra un sesgo a la derecha (Fig. 7a), Beaumont, Hughes y Poulsen, (2005) determinaron que, una distribución normal o sesgada de la variable, es relevante para definir el perfil bioclimático de la especie, mientras que otros autores (Méndez et al., 2021), indican que es indicio de una adecuada elección del predictor.
TABLA 2 Perfil bioclimático actual y futuro de Pinus engelmannii Carriére elaborado con 346 registros, cuatro modelos de circulación general y dos RCP´s 4.5 y 8.5.
| CCSM4 | ¶ | CNRM-CM5 ¶ | CSIRO-Mk3-6-0 ¶ | HadGEM2-AO ¶ | ||||||||
| Actual | RCP 4.5 | RCP 8.5 | RCP 4.5 | RCP 8.5 | RCP 4.5 | RCP 8.5 | RCP 4.5 | RCP 8.5 | ||||
| Bio 1 | 9.8 - 25.7 | 11.4 - 27.0 | 11.8 - 27.8 | 11.4 - 27.0 | 11.7 - 27.3 | 11.9 - 27.5 | 12.1 - 27.7 | 11.8 - 27.4 | 11.9 - 27.5 | |||
| Bio 17 | 7 - 72 | 5 - 60 | 4 - 51 | 4 - 50 | 5 - 47 | 7 - 74 | 9 - 86 | 4 - 55 | 6 - 68 | |||
| Bio 16 | 205 - 1073 | 185 - 1050 | 216 - 1064 | 202 - 1135 | 216 - 1091 | 196 - 1069 | 211 - 1125 | 186 - 1123 | 179 - 1072 | |||
| Bio 15 | 64 - 120 | 65 - 119 | 71 - 127 | 68 - 123 | 70 - 124 | 64 - 120 | 62 - 119 | 68 - 125 | 60 - 121 | |||
| Bio 5 | 20.1 - 36.2 | 21.2 - 37.8 | 22.0 - 38.9 | 21.9 - 38.3 | 21.9 - 38.0 | 22.3 - 39.2 | 22.8 - 38.8 | 22.3 - 38.5 | 22.2 - 38.6 | |||
MCG´s = Modelos de circulación general; RCP = Trayectorias Representativas de Concentración de CO2.

FIGURA 7 Variables ambientales de mayor contribución al modelo de nicho de Pinus engelmannii Carriére: Bio 1(a) y Bio 17 (b) en el periodo actual (barras obscuras) y al 2050 (líneas), promedio de cuatro MCG´s en dos trayectorias de concentración RCP´s 4.5 y 8.5). La altitud (c) se presenta como complemento.
Discusión
El ajuste y las variables del modelo del género Pinus
Peterson et al. (2008) , indican que un modelo es mejor que lo aleatorio si el valor de ROC parcial está por encima de 1, por lo tanto, el modelo generado aquí es bueno, al obtener un ROC de 1.3. Al modelar la distribución actual de Pinus montezumae y Pinus pseudostrobus,Manzanilla et al. (2019) obtienen valores de ROC parcial de 1.312 y de 1.381; a pesar de que ROC parcial es una buena métrica del ajuste del modelo (Peterson et al., 2008), pocos son los autores que la utilizan hoy día.
En general, se observa que en los modelos de nicho de las especies del género Pinus, la temperatura media anual (Bio 1) es la variable que mejor predice la distribución de las especies. Por ejemplo, Aceves-Rangel et al. (2018) , modela- ron la distribución actual de esta misma especie, encontrando que Bio 1, es variable que más contribuye al modelo con 24.9%. En un trabajo similar realizado por Ramos-Dorantes, Villaseñor, Ortiz y Gernandt (2017) encuentran que Bio 1, es la más importante en 10 de 16 especies de coníferas. Manzanilla et al. (2019) indican que la variable de mayor contribución al modelo de nicho de P. pseudostrobus es Bio1. Eguiluz, (1982) al igual que este estudio, demuestra que la distribución del género Pinus, depende en su gran mayoría de la temperatura. Sin embargo, la contribución de cada variable además de la especie depende del número de registros y su distribución, número y tipo de variables incluidas en la modelación, tipo de análisis y algoritmo (Elith et al., 2006), por lo que en cada estudio podría diferir, incluso si se trata de la misma especie.
En contraparte, en Pinus pinceana Gordon, Bio 5 es la que mejor explica su distribución actual (Cruz-Cárdenas et al., 2016), pero en Pinus cembroides Zucc., es Bio 9 (García- Aranda et al., 2018) y Bio 1 (Aceves-Rangel et al., 2018). Alba, González, Ramírez y Castillo (2003), al realizar análisis de componentes principales, con 10 variables ambientales y 660 registros de presencia de nueve especies de coníferas, encuentran que el primer componente se relaciona con lluvia y el segundo con la temperatura.
En otros géneros como Abies, en seis de ocho especies y dos variedades, Bio 19 y Bio 2 (precipitación del cuarto más frío e intervalo de temperatura media diurna), son las más comunes (Martínez et al., 2016).
Áreas idóneas actuales de Pinus engelmannii Carriére
Chihuahua, Durango y Sonora, en ese orden son los estados donde ocurren las áreas más extensas e idóneas para P. engelmannii. La alta idoneidad de esta especia ha sido encontrada en estos mismos lugares (Aguirre y Duivenvoorden, 2010; Aceves-Rangel et al., 2018), quienes indican que su modelo es bueno al obtener AUC de 0.991 y de 0.778 respectivamente. Los primeros autores encuentran una extensión de 318 203.8 km2 de áreas idóneas para esta especie en México, cuya cifra difiere a la de este estudio por la metodología, número de registros empleados y la no estratificación de estos autores. Otros autores (Rzedowski, 2006; González- Elizondo et al., 2012; Bustamante-García et al., 2012), demuestran que la distribución de esta especie ocurre principalmente en el estado de Durango. No obstante, Sánchez-González, (2008) indica que además de estas regiones, la especie también se encuentra en las regiones I y III (BC y Sierra Madre Oriental); lugares en los cuales no se encontraron registros.
Las modalidades de la predicción potencial de Pinus engelmannii Carriére y de otras especies
La modalidad NE (no extrapolación) predice mejor la distribución potencial de P. engelmannii, mientras que EL y EC predicen de manera ilógica, este mismo patrón fue encontrado por Owens et al. (2013) , las modalidades EL y EC, exageraron las predicciones de Bactrocera invadens Drew, Tsuruta & White y Anopheles gambiae Giles, optando por elegir la predicción NE; al parecer esto depende de los cambios que ocurren en las variables bioclimáticas cuando el modelo se trasfiere a otras áreas o tiempos, ya que Escobar et al. (2014) y Vilges et al. (2017) encontraron que EL y EC, predicen de la misma forma, mientras que Cobos et al. (2019) indican que la modalidad que mejor predicen la distribución de sus especies fue EL.
La respuesta de las especies al cambio climático
La respuesta al cambio climático varía entre especies, en la mayoría de los casos la migración es una estrategia a la sobrevivencia (Aitken et al., 2008), esto solo puede identificarse si se realizan más predicciones. Sáenz-Romero et al. (2010) encuentran que el nicho P. chiapensis para el año 2060 estaría ubicada a mayores latitudes, por el contrario, Pinus leiophylla. Schiede ex Schltdl. & Cham. tiende a migrar a menores latitudes (Sáenz-Romero et al., 2015). En un estudio realizado por Cruz-Cárdenas et al. (2016) en doce especies de coníferas, se encontró que la reducción de las áreas de idoneidad podría ascender hasta 40% al 2075-2099, si las emisiones de CO2 son altas (RCP 8.5), siendo Abies religiosa (Kunth) Schltdl. & Cham, P. leiophylla y P. teocote Schiede ex Schltdl. & Cham., las más afectadas. Lenoir et al. (2008) indican que, en Europa, como resultado del calentamiento global, las especies han migrado hacia mayores altitudes a una tasa de 29 m por década. Al parecer P. engelmannii Carriére podría ser una de las pocas especies que no es vulnerable al cambio climático; la modelación de nicho de este estudio, con climatologías de cuatro MCG’s dan evidencias de esto, no se predicen nuevas áreas idóneas, ni se pierden las áreas idóneas existentes (Fig. 5), incluso si existiera un cambio climático drástico (Fig. 5b); en regiones cercanas a la distribución actual de la especie, se observan áreas idóneas, cerca del área de estudio, pero no es indicio de migración.
Conclusiones
Las variables que determinan la distribución de Pinus engelmannii Carriére son: Bio 1 y Bio 17 con 75.6% y 6.3%, respectivamente. La idoneidad climática más alta se registra en Chihuahua, Durango y Sonora, pero de forma discontinua. Se rechaza la hipótesis planteada; a diferencia de la mayoría de las especies de Pinus, al parecer esta especie no es susceptible al cambio en el clima, a pesar de un incremento de casi 2.0 °C en la temperatura media anual al 2050 (RCP 8.5), donde actualmente se distribuye la especie («M»), e independientemente de si las emisiones de CO2 al 2050 sean altas (RCP 8.5) o bajas (RCP 4.5). Dentro de («M»), no se predicen nuevas áreas idóneas, ni se pierden las áreas idóneas existentes, es posible que la discontinuidad observada de las áreas de idoneidad sea indicio de fragmentación del hábitat. Al transferir el modelo, se observa la existencia de nuevas áreas idóneas en México, Estados Unidos (42 ºN) y Sudamérica (44 ºS), en particular las áreas idóneas actuales y las nuevas áreas idóneas del país deberían ser consideradas prioritarias para conservar e incrementar la distribución de la especie, considerando su reducida distribución actual.

