











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
Ante la deforestación y cambio de uso de suelo, la conservación de la vegetación arbórea, en general, tiene entre sus múltiples beneficios, el almacén de carbono orgánico en la corteza terrestre, ya que, dadas las dimensiones de la forma de vida de los árboles, estos constituyen las mayores reservas de los ecosistemas terrestres por unidad de área (Sullivan et al., 2017).
En lo que se refiere a los sistemas agroforestales (SA) más similares a la selva original (tipo rústico) en particular (sensu Rice y Greensberg, 2000), a escala global según Albrecht y Kandji (2003), estos podrían contribuir a almacenar entre 1 100 000 Mg y 2 200 000 Mg de carbono (C) si se establecieran entre las 585 × 106 ha y 1215 × 106 ha del mundo con potencialidad para esta práctica. De esta manera, una hectárea de SA de cacao (Teobroma cacao L., Malvaceae) almacena más C aéreo (270 Mg ha-1 - 337 Mg ha-1) (Duguma, Gockowski y Bakala, 2001; Yquise-Pérez, Pocomucha y Vargas, 2009) que un pastizal tropical (4.25 Mg ha-1 - 9.0 Mg ha-1, 2.25 Mg ha-1) (Alegre, Arévalo y Ricse, 2002; Ramos-Hernández y Martínez-Sánchez, 2020) o campo de cultivo (42 Mg ha-1) (Duguma, et al., 2001).
Las selvas medianas subperennifolias que caracterizan la vegetación original de la zona cacaotera del estado de Tabasco poseen una media de almacenamiento de carbono en vegetación arbórea entre 21 Mg ha-1 y 50 Mg ha-1 (Montagnini y Nair, 2004), las cuales, debido a la presión socioeconómica, han estado en riesgo permanente de conversión a pastizales para la ganadería. En este sentido, los SA de T. cacao del estado de Tabasco adquieren mayor importancia para la conservación de vegetación arbórea que los pastizales establecidos de ganadería extensiva.
Los SA, en el estado de Tabasco, son particularmente importantes como proveedores del servicio ambiental de almacenamiento de carbono debido a la vegetación arbórea presente para proveer sombra. Si bien, el almacén de carbono en el suelo generalmente es mayor que en la parte aérea para cacaotales de 25 años en Perú (González-Huiman, Chávez-Matías y Bringas-Paredes, 2011; Yquise Pérez et al., 2009), en el presente trabajo se consideró evaluar el almacén de carbono aéreo debido a que, en presencia de cambio de uso del suelo, es más variable que el carbono subterráneo que es más estable.
La mayor diversidad y almacén de carbono superficial en un ecosistema arbóreo se encuentra en su edad madura, lo cual también puede ser cierto para un SA de cacao de tipo rústico. El carbono vegetal se almacena principalmente en el componente arbóreo adulto proveedor de la sombra al cacao y, en menor proporción, en la plantación de cacao. Haciendo una cronosecuencia de edad de los SA de cacao para los almacenes de C a partir de los estudios disponibles al momento, Duguma et al. (2001) registraron, en SA de 5 años en Camerún, 5.85 Mg de carbono por hectárea; Beer et al. (1990) 11 Mg ha-1 en Costa Rica. Koskela, Nygren, Berninger y Luukkanen (2000), en SA de 10 años con especies arbóreas como Cordia alliodora y Erythrina poeppigiana, estimaron 42.8 Mg ha-1 y 130.8 Mg ha-1 en el arbolado respectivamente. En Perú, González-Huiman et al. (2011) cuantificaron almacenes de C de 80 Mg ha-1 en el arbolado de SA de 11 años, Concha, Alegre y Pocomucha (2007) entre 12 Mg ha-1 y 32.4 Mg ha-1 en SA de 5 años y 20 años, respectivamente e Yquise Pérez et al. (2009) 216.3 Mg ha-1 en un SA de 25 años, mientras que 337.5 Mg ha-1 en una selva primaria. En Camerún, un SA de cacao de 26 años almacenó 152 Mg de carbono por hectárea (Duguma et al. 2001). Norgrove y Hauser (2013) cuantificaron 231.3 Mg ha-1 en un SA de 35 años en Camerún, de lo cual, 52.4% correspondió a los árboles de sombra (121.1 Mg de carbono por hectárea) y 5.4% a los árboles de cacao (12.5 Mg ha-1).
Objetivos
El objetivo del presente trabajo fue analizar la relación de la diversidad y los almacenes de carbono de la vegetación arbórea de sombra, a lo largo de una cronosecuencia de SA de cacao en el estado de Tabasco, México. La hipótesis por probar fue que, a medida que aumenta la edad de los SA de cacao, también aumentan la diversidad arbórea y almacenes de C de su vegetación de sombra, debido a la similitud con la estructura madura de su vegetación original.
Materiales y métodos
El trabajo se realizó en SA de cacao establecidos en los municipios de Comalcalco y Cunduacán, Tabasco, México (18° 10’ 33.56’’ y 18° 15’ 34.05’’ N - 93° 40’ 36.69’’ y 93° 34’ 57.88’’ O), donde prevalece un clima cálido subhúmedo con lluvias en verano (Am), temperatura media anual de 26 °C y precipitación anual de 2500 mm (Instituto Nacional de Estadística y Geografía [Inegi], s/f). Los suelos dominantes son cambisoles, histosoles y fluvisoles (Palma-López et al., 2017), y se encuentran entre10 m y 20 m s.n.m. La vegetación dominante son pastizales para ganadería extensiva y, en menor proporción, relictos de selva mediana subperennifolia (Flores y Geréz, 1994).
La cronosecuencia de los SA de T. cacao se construyó a escala de plantación o finca, a partir de la selección de 11 SA con edades diferentes de 10, 13, 15, 20, 30, 30, 35, 40, 45, 48 y 70 (todas ellas expresadas en años), a decir de sus dueños, y con una extensión de 0.04 ha a 2.85 ha cada una. La vegetación de sombra de los SA, en general, está constituida por especies arbóreas originales seleccionadas mediante manejo forestal como poda y derribo. Las especies frutales arbóreas, fueron plantadas. En cada SA se ubicaron entre 3 y 239 unidades de muestreo (258 en total; Tabla 1) de diferentes tamaños (10 m × 10 m, 20 m × 20 m, y 20 m × 50 m), ya que los datos provienen de tres estudios independientes en tiempos distintos. Se midió el DN (aprox. a 1.40 cm de altura) y altura total de todos los árboles > 5 cm de DN, y cada árbol se determinó mediante ejemplares del Herbario de la Universidad Juárez Autónoma de Tabasco. Los SA en general presentaban poca perturbación a no ser por poda de ramas en dos sitios de Comalcalco. Debido a que su objetivo es la producción de cacao, en el estado de Tabasco su longevidad máxima es de alrededor de 50 años (Córdova-Avalos et al., 2001; Ramírez-Meneses et al., 2013), después de lo cual, la plantación se renueva con árboles jóvenes.
TABLA 1 Número de unidades de muestreo y superficie total muestreada en sistemas agroforestales de cacao de diferente edad en Tabasco, México.
| Edad (años) | Parcelas | Superficie (ha) |
| 10 | 2 | 0.8 |
| 13 | 1 | 0.4 |
| 15 | 2 | 0.8 |
| 20 | 2 | 0.8 |
| 30 | 2 | 0.8 |
| 30 | 2 | 0.08 |
| 35 | 1 | 0.4 |
| 40 | 5 | 0.5 |
| 45 | 239 | 2.85 |
| 48 | 1 | 0.4 |
| 70 | 1 | 0.04 |
Con los datos obtenidos se determinó la densidad arbórea, la riqueza de especies observada, la riqueza de especies estimada por el algoritmo Chao 1 y la diversidad de Shannon y Simpson para cada uno de los 11 SA ubicados en los dos municipios, por medio del programa EstimateS Version 8 (Colwell, 2009). Para el análisis, se usaron 100 reiteraciones al azar sin reemplazo de muestras y la corrección de desviación para el estimador de riqueza Chao 1; esto debido a que las unidades de muestreo de los 11 SA fueron de diferente tamaño, número y superficie. Finalmente, se realizaron curvas de acumulación de especies para probar la estimación adecuada de la diversidad arbórea en relación con la superficie muestreada.
Para estimar la biomasa aérea arbórea, se utilizó la ecuación alométrica general para selvas tropicales de Chave et al. (2014) y 24 diferentes ecuaciones específicas (Tabla 2) obtenidas de las bases de datos de Henry et al. (2013) y Rojas, de Jong, Martínez-Zurimendí y Paz-Pellat (2015). Para la selección de la ecuación específica dentro de un gran número de ecuaciones disponibles para una misma especie, se consideró que fuera específica para la especie de interés, o en su defecto, para su género taxonómico, así como un adecuado tamaño de muestra y coeficiente de determi-nación (r2). El valor de la densidad de madera para las especies arbóreas se obtuvo de la base de datos de Henry et al. (2013) y Anónimo. El valor de la biomasa de cada árbol se multiplico por 0.5 para obtener el valor del carbono orgánico (Pearson, Walker y Brown, 2005). Los valores de biomasa aérea obtenidos para cada árbol > 5 cm de DN, independientemente de su DN, categoría diamétrica o edad, se sumaron para todas las unidades de muestreo de cada SA, con el cual se estimó el valor para una hectárea por simple regla de tres para cada uno de los 11 SA. Esto se hizo por separado para los valores estimados con la ecuación general y con la ecuación específica.
TABLA 2 Ecuaciones alométricas específicas utilizadas en la estimación de biomasa aérea (BA, kg) de cada especie arbórea.
| Especie | Ecuación | Autor | ||
| 1 | Muchas (Ecuación general) | 1 | BA = 0.0673 * (WD*D2H)0.976 | Chave et al. (2014) |
| 2 | Annona muricata L. | 2 | log BA=-1.716+2.413*LOG((DBH)) | Nogueira, Fearnside, Nelson, Barbosa y Keizer (2008) |
| 3 | Artocarpus altilis | 3 | BA=0.0704*((DBH)^(2)*(H)*(WD))^(0.9389) | Hung, Son y Hung (2012) |
| 4 | Bursera simaruba (L.) Sarg. | 4 | BA=(0.064808*(DBH)^2.46998) | Rodríguez, Jiménez, Meza, Aguirre y Razo (2008) |
| 5 | Byrsonima crassifolia | 5 | log BA=1.087*exp(-2.232+2.422*log((DBH))) | Sierra et al. (2007) |
| 6 | Bixa orellana | log BA=-1.716+2.413*LOG((DBH)) | Nogueira et al. (2008) | |
| 7 | Castilla elastica | log BA=1.087*exp(-2.232+2.422*log((DBH))) | Sierra et al. (2007) | |
| 8 | Cecropia obtusifolia Bertol. | 6 | BA=12.764 + 0.2588 [DBH(2.0515)] | Pearson et al. (2005) |
| 9 | Cedrela odorata | 7 | BA=0.0013*(DBH)^(2)*(H)^(0.9218)+0.0072*(DBH)^(2)*(H)^(1.0451)+ 0.0029*(DBH)^(2)*(H)^(1.0172)+0.0010*(DBH)^(2)*(H)^(0.8038)+ 0.0893*(DBH)^(2)*(H)^(0.5326) | Cole y Ewel (2006) |
| 10 | Ceiba pentandra (L.) Gaerth | 8 | BA=0.194072*(DBH)^(2.20172)*((None)D)^(0.172693) | Henry et al. (2010) |
| 11 | Citrus aurantium | 9 | BA=EXP(-3.08+1.007*LN(D)*Ln(H)*LN(WD)) | Chave et al. (2005) |
| 12 | Citrus sinensis (L.) Osb. | BA=EXP(-3.08+1.007*LN(D)*Ln(H)*LN(WD)) | Chave et al. (2005) | |
| 13 | Cocos nucifera | 10 | BA= 6.666 + 12.826 / height [0.5(ln(height))] | Pearson et al. (2005) |
| 14 | Colubrina arborescens (Mill.) Sarg. | BA=EXP(-3.08+1.007*LN(D)*Ln(H)*LN(WD)) | Chave et al. (2005) | |
| 15 | Cordia alliodora (Ruis y Pav.) Oken | 11 | BA=0.0391*(DBH)^(2)*(H)^(0.5151)+0.0085*(DBH)^(2)*(H)^(1.045)+ 0.0853*(DBH)^(2)*(H)^(0.5345)+0.0427*(DBH)^(2)*(H)^(0.6437) | Cole y Ewel (2006) |
| 16 | Cupania dentata | 13 | log BA=-1.9968+(2.4128*LOG((DBH))) | Nelson et al. (1999) |
| 17 | Diphysa robinioides Benth | BA=EXP(-3.08+1.007*LN(D)*Ln(H)*LN(WD)) | Chave et al. (2005) | |
| 18 | Erythrina americana Mill | 14 | BA=(0.37*(DBH)^1.96) | Návar (2009) |
| 19 | Erythrina fusca | BA=(0.37*(DBH)^1.96) | Návar (2009) | |
| 20 | Gliricidia sepium (Jacq.) | 15 | BA=0.1185*DBH^2 | Foroughbakhch et al., (2006) |
| 21 | Gmelina arbórea Roxb | 16 | BA=0.1274*(DBH)^(2.3655) | Hung et al. (2012) |
| 22 | Guazuma ulmifolia Lam. | 17 | BA=(0.232435*(DBH)^2.21906) | Rodríguez et al. (2008) |
| 23 | Heliocarpus donell-smithii | 18 | BA=(Exp(4.9375)*(DBH)^2.1166)*1.14e-006 | Hughes, Kauffman y Jaramillo (1999) |
| 24 | Inga jinicuil | 19 | log BA-2.904+0.993*LOG(DBH)^(2)*H*(WD) | Overman, Witte y Saldarriaga (1994) |
| 25 | Lysiloma bahamensis | 20 | BA=Exp[-1.566]*[DBH^2.276] | Gómez, (2008) |
| 26 | Manilkara zapota | log BA=-2.904+0.993*LOG((DBH)^(2)*(H)*(WD)) | Overman et al. (1994) | |
| 27 | Mangifera indica L. | 21 | BA=0.0968*(((DBH)^(2))*((H)^(0.7))*(WD))^10037 | Hung et al. (2012) |
| 28 | Persea americana Mill. | BA=EXP(-3.08+1.007*LN(D)*Ln(H)*LN(WD)) | Chave et al. (2005) | |
| 29 | Pimenta dioica | BA=EXP(-3.08+1.007*LN(D)*Ln(H)*LN(WD)) | Chave et al. (2005) | |
| 30 | Pouteria mammosa | 22 | BA=(0.0358*((DBH)^2*(H))^1) | Cairns, Olmsted, Granados y Argaez (2003) |
| 31 | Pouteria sapota | BA=(0.0358*((DBH)^2*(H))^1) | Cairns et al. (2003) | |
| 32 | Psidium guajava Nied. | 23 | BA=(0.246689*(DBH)^2.24992) | Rodríguez et al. (2008) |
| 33 | Sabal mexicana Mart | BA= 6.666 + 12.826 / height [0.5(ln(height))] | Pearson et al. (2005) | |
| 34 | Samanea saman | BA= 6.666 + 12.826 / height [0.5(ln(height))] | Pearson et al. (2005) | |
| 35 | Spondias mombin | log BA=-1.716+2.413*LOG((DBH)) | Nogueira et al. (2008) | |
| 36 | Swietenia macrophylla | 24 | BA=(-1.32)+(2.65*log((DBH))) | Krisnawati, Adinugroho y Imanuddin (2012) |
| 37 | Tabebuia rosea | log BA=-1.716+2.413*LOG((DBH)) | Nogueira et al. (2008) | |
| 38 | Theobroma cacao | 25 | log BA=−1.684+ 2.158∗log (𝑑30)+0.892∗log (𝐻) | Cerda, Espin y Cifuentes (2013) |
WD = densidad de madera.
Debido a la existencia básica de dos tipos de modelos alométricos (generales y específicos), se usó la prueba de t de Student con la intensión de comparar la diferencia en su estimación. Para el análisis de la relación de la diversidad arbórea y almacenes de carbono con la edad de los SA se usó la correlación de Pearson (STATGRAPHICS® Centurion XVI y Microsoft Excel®).
Resultados
Estructura arbórea en una cronosecuencia de sistemas agroforestales de cacao
La superficie muestreada en cada SA fue suficiente para representar su diversidad arbórea, ya que, a partir de determinado número de unidades de muestreo, se alcanzó la asíntota, no encontrándose especies nuevas con el incremento del área de muestreo; se ilustran tres casos (Fig. 1). Aparte del cacao, también se plantaron otras especies frutales como naranja, limón, mango, palma de coco, entre otras.
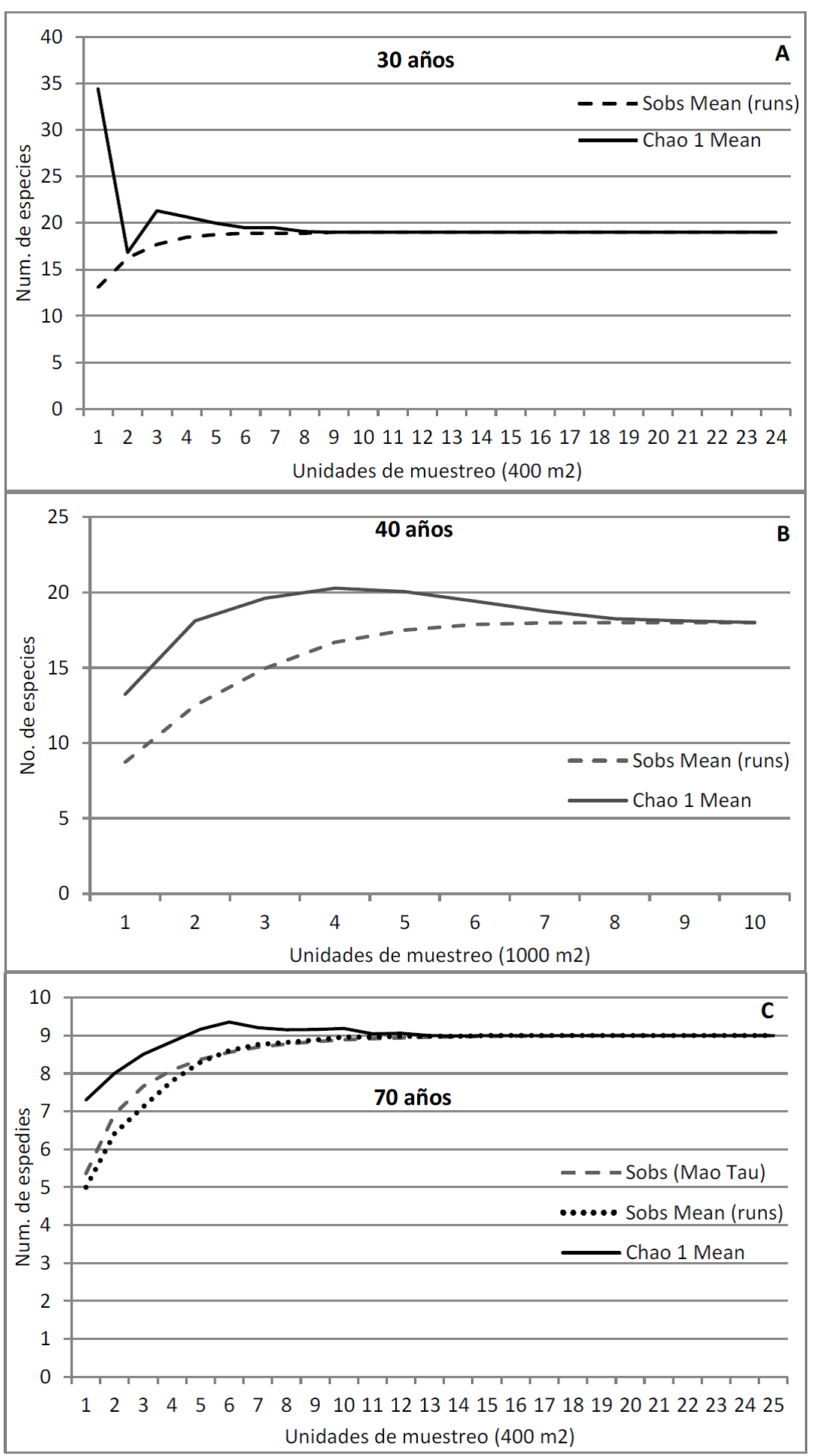
Las curvas describen cómo a medida que aumenta la superficie muestreada ya no se encuentran nuevas especies y la línea se hace horizontal.
FIGURA 1 Curvas de acumulación de especies arbóreas en tres SA de cacao de diferente edad en el estado de Tabasco, México.
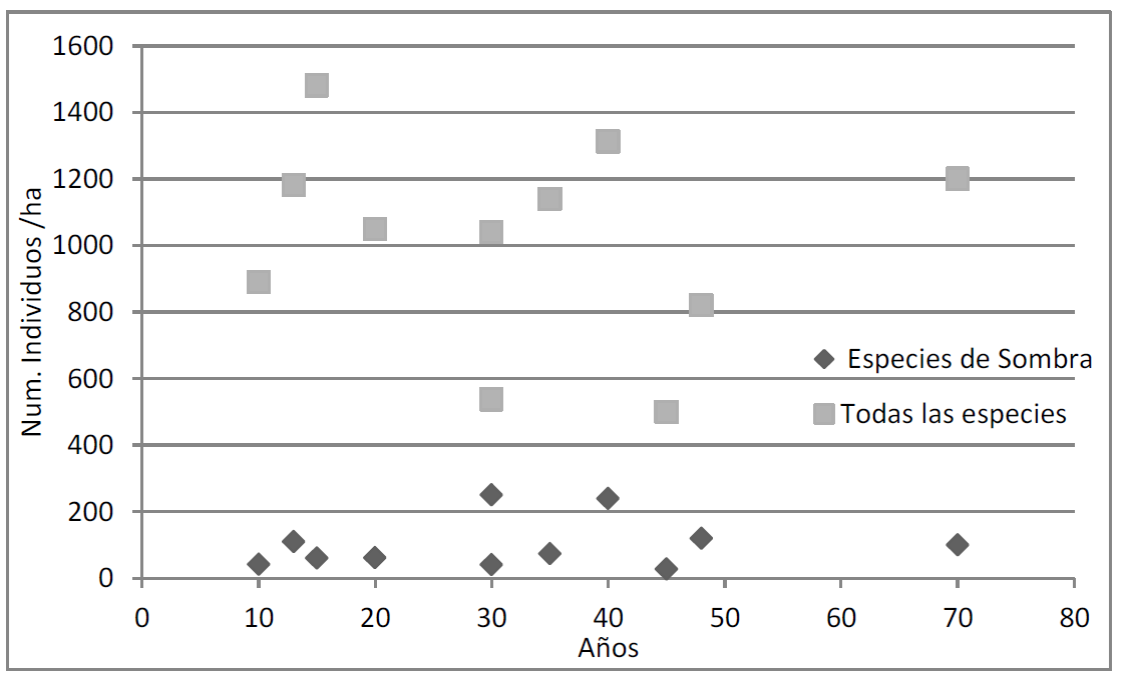
La densidad total de especies cultivadas (mayormente cacao) en los 11 SA fue entre 287 ind ha-1 y 1420 ind ha-1, con una densidad media de 911.2 ind ha-1 ± 318.34 ind ha-1 (Tabla 3), mientras que la densidad media de individuos de las especies arbóreas de sombra fue de 106.4 ind ha-1 ± 76.4 ind ha-1 y la densidad media de individuos de todas las especies fue de 1013.5 ind ha-1 ±305.1 ind ha-1 (Fig. 2).
TABLA 3 Densidad por hectárea de individuos de especies plantadas, en sistemas agroforestales de cacao de diferente edad en Tabasco, México.
| Edad (años) | Árboles (ha-1) |
| 10 | 848 |
| 13 | 1070 |
| 15 | 1420 |
| 20 | 988 |
| 30 | 999 |
| 30 | 287 |
| 35 | 1066 |
| 40 | 1072 |
| 45 | 473 |
| 48 | 700 |
| 70 | 1100 |
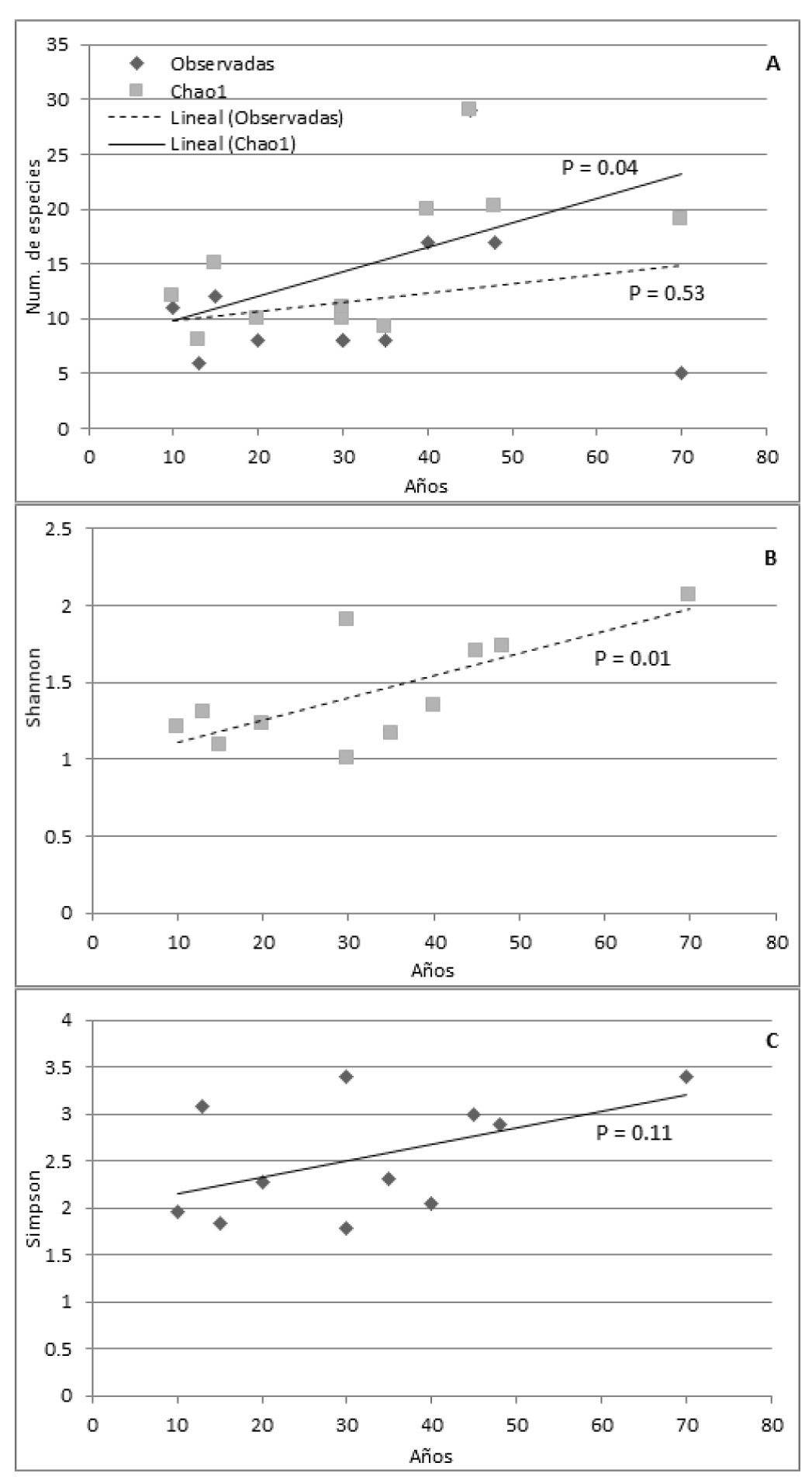
A diferencia de la riqueza observada de especies arbóreas por hectárea (p = 0.53; 11.73 ± 7), la riqueza estimada por Chao 1 presentó una relación positiva con la edad de los SA de cacao (p = 0.04; 14.85 ± 6.5; Fig. 3A). La edad de los SA presentó relación con la diversidad a través del índice Shannon (p = 0.01; 1.4 ± 0.36; Fig. 3B) pero no a través del índice Simpson (p = 0.11; 2.5 ± 0.64; Fig. 3C).
Almacenes de carbono en una cronosecuencia de sistemas agroforestales de cacao
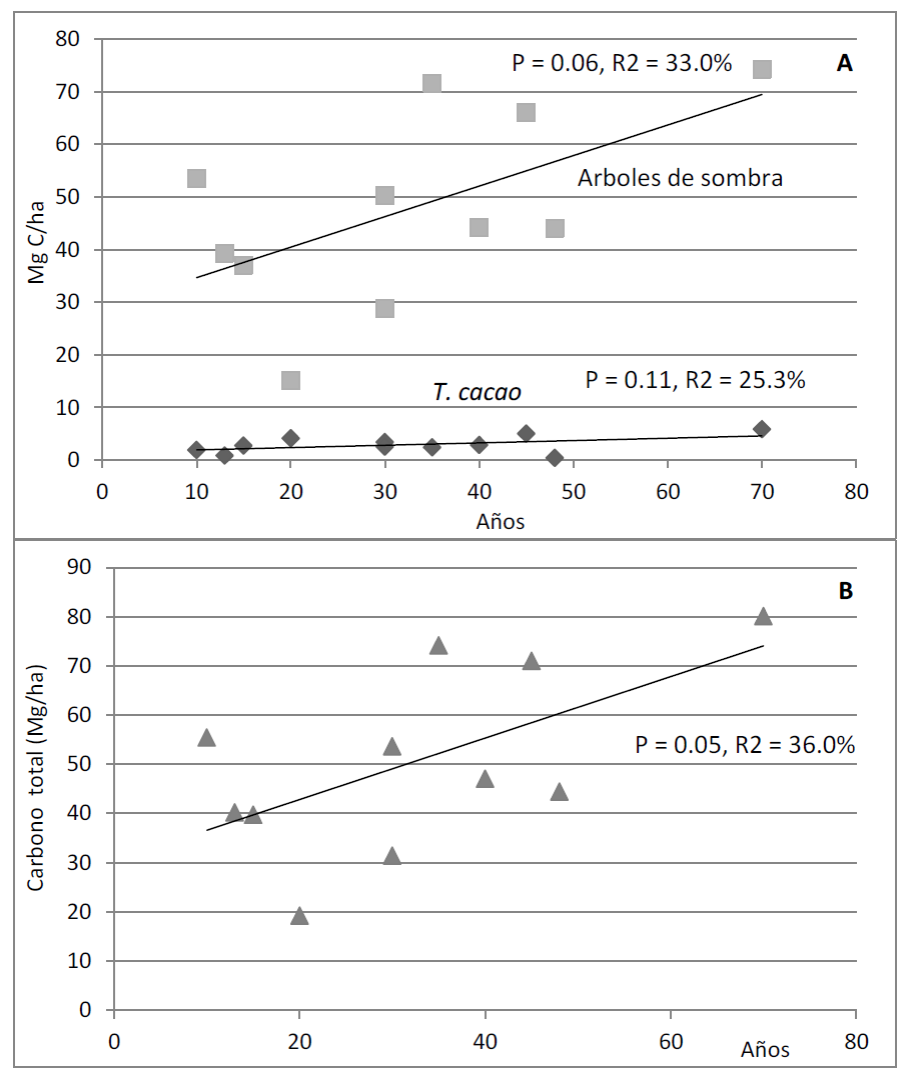
La estimación del almacén de carbono arbóreo (Mg ha-1) a partir de las ecuaciones para cada especie resultó ser 27.7% mayor que el valor estimado a través de la ecuación general de Chave et al. (2014) para selvas tropicales (p = 0.05; t = 1.76; 47.25 vs. 34.2; Fig. 4).
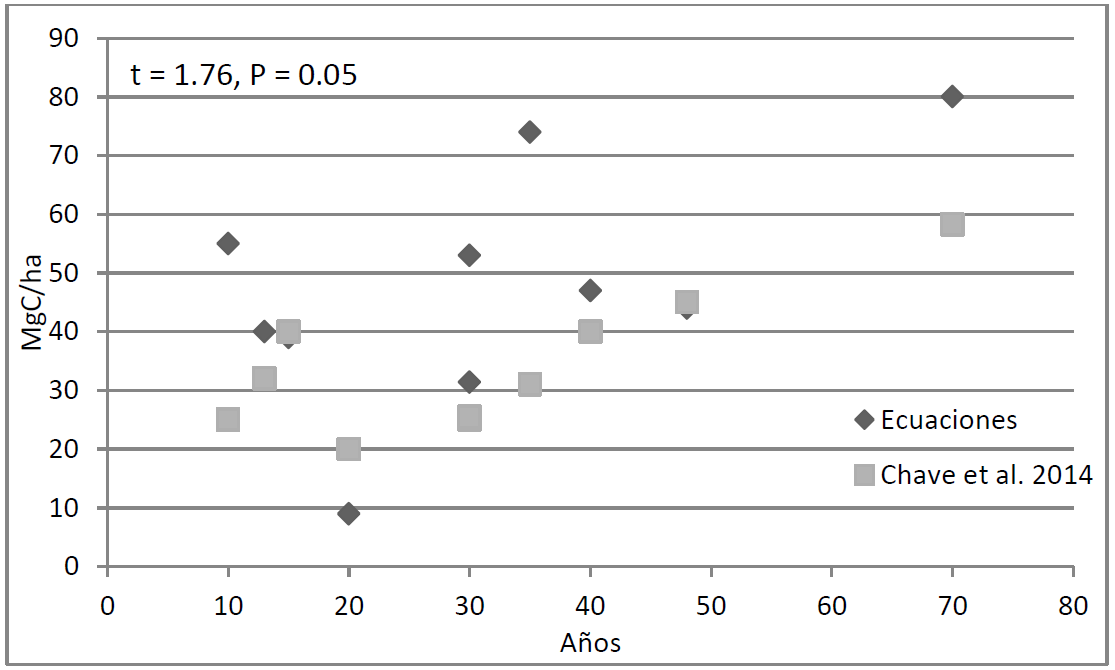
FIGURA 4 Estimación de carbono en los sistemas agroforestales de cacao de diferente edad, mediante el método de la ecuación general de Chave et al. (2014) y el método de ecuaciones específicas de biomasa arbórea.
La acumulación del carbono arbóreo respecto a la cronosecuencia de los SA no tuvo una relación particular con las especies arbóreas de sombra (p = 0.06), tampoco con la especie de T. cacao (p = 0.11), pero sí presentó una relación positiva con el SA o la combinación de los dos componentes arbóreos (árboles de sombra y de T. cacao juntos; p = 0.05; Fig. 5).
Discusión
Diversidad arbórea de los sistemas agroforestales de cacao
Similarmente a la selva madura, el SA de cacao guarda una relación positiva entre su edad y la diversidad arbórea que presenta (Fig. 3 A, B). Es bien conocida la gran ventaja biológica y ambiental que representa la conservación de la diversidad arbórea en los ecosistemas terrestres (Asase y Tetteh, 2010; Asase, Asitoakor y Ekpe, 2012). En este sentido, se puede decir que los SA de cacao de esta región se encuentran entre los que presentan mayor diversidad arbórea a escala mundial (Tabla 4), lo cual es relevante para fines de conservación biológica. Si se considera que una hectárea de vegetación arbórea original de 40 años en esta región presenta alrededor de 27 especies arbóreas (Martínez-Sánchez y Cámara, 2012), la riqueza de 20 especies arbóreas por hectárea (Fig. 3A) en un SA de cacao de 40 años, o 29 especies arbóreas por hectárea en uno de 45 años, no está muy lejos de lo que era su riqueza arbórea original.
TABLA 4 Sistemas agroforestales de cacao en otros países del mundo.
| Especies arbóreas de sombra presentes | País | Autor |
| Cordia alliodora | Costa Rica | Alpizar, Fassbender, Heuveldop, Folster y Enríquez (1986) |
| Erythrina fusca | Brazil | Delabie et al. (2007) |
| Erythrina glauca | Brazil | Kummerow, Kummerow, y Da Silva (1982) |
| Erythrina poeppigiana | Costa Rica | Muñoz y Beer (2001) |
| Inga edulis o Guazuma crinita | Perú | Yquise Perez et al. (2009) |
| Albizia zygia o Milicia excelsa o Newbouldia laevis | Ghana | Isaac, Timmer y Quashie-Sam (2007) |
| Erythrina glauca y E. poeppigiana, o Genipa americana o Spondias lutea | Brazil | Santana y Cabala (1982) |
| Una a cuatro especies | Costa de Marfil | Herzog (1994) |
| Pithecelobium saman, Erythrina americana, Gliricidia sepium, Inga jinicuil e I. vera | Tabasco, México | Muñoz, Estrada y Naranjo (2005) |
| Cinco especies arbóreas en cacaotales de 20 años | Perú | Concha et al. (2007) |
| Cinco a 10 especies arbóreas | Costa Rica | Beer (1988) |
| Promedio de 5.4 y 6.5 especies arbóreas > 15 cm de dap | Costa Rica | Reitsma, Parrish y McLarney (2001) |
| 8 a 29 especies arbóreas | Tabasco, México | Este estudio |
| 60 especies arbóreas en 12 ha | Tabasco, México | Greenberg et al. (2000) |
| 32 especies arbóreas | Tabasco, México | Muñoz et al. (2006) |
| 35 especies arbóreas | Costa de Marfil | Herzog (1994) |
| 53 especies arbóreas en 0.2 ha | Camerún | Norgrove y Hauser (2013) |
Almacenes de carbono de los sistemas agroforestales de cacao
El almacén de carbono de los árboles de sombra a partir de las ecuaciones alométricas para cada especie de 11 SA entre 10 años y 70 años resultó ser 27.7% mayor que el estimado a través de la ecuación general de árboles para selvas tropicales de Chave et al. (2014). Esto sugiere que el utilizar las ecuaciones de cada especie, al menos para este tipo de SA, es más preciso que la estimación a través de un modelo general construido a partir de un gran número de especies arbóreas de forma diferente, provenientes de un gran número de tipos de selvas húmedas y subhúmedas de los continentes americano y asiático. También Martínez-Sánchez, Martínez-Garza, Cámara y Castillo (2020) llegaron a esta conclusión, a partir de una comparación global de ecuaciones específicas y ecuaciones generales para la estimación de la biomasa arbórea en vegetación de selvas tropicales del sureste de México. Por lo anterior, se recomienda seguir comparando ambos métodos en otros SA en general. Los resultados muestran que tanto los árboles de cacao como la vegetación arbórea de sombra por sí mismos, no representan una ganancia en la acumulación del carbono aéreo de estos SA, sino más bien, el SA como tal, a partir de estos dos componentes, es el que sí constituye una ganancia en la acumulación de carbono aéreo con la edad. Esto indica que el manejo adecuado de la vegetación original con inclusión de cacao puede representar una ventaja en el almacén y conservación del carbono en los ecosistemas arbóreos tropicales.
Comparación con otros sistemas agroforestales de cacao en el mundo
Las ventajas y cualidades de estos SA en particular respecto a la conservación de su biodiversidad arbórea y carbono orgánico son más evidentes cuando se comparan con otros SA de cacao en el mundo. Al contrastar los SA de cacao de Tabasco, México, con SA en otros países importantes en la producción de cacao, la mayoría de los SA en el mundo presentan plantaciones uniespecíficas o de pocas especies arbóreas proveedoras de sombra a diferencia de México (Tabla 4).
Los SA de cacao en regiones como África y Sudamérica, generalmente están asociados con una baja diversidad de una o dos especies de árboles, a diferencia de México donde se asocian con una mayor diversidad. En este sentido, cuanto mayor diversidad arbórea tengan los SA de cacao, ofrecen un mayor servicio ecológico ya que la diversidad arbórea se encuentra asociada a la diversidad de otros grupos de plantas vasculares y de fauna, ornitológica principalmente (Alves, 1990; Majer, Delabie y Smith, 1994; Estrada, Coates-Estrada y Meritt, 1993; Estrada, Coates-Estrada, Meritt, Montiel y Curiel, 1993; Estrada, Coates-Estrada y Meritt, 1994; Estrada, Coates-Estrada y Meritt, 1997; Parrish, Reitsma y Greenberg, 1998; Muñoz, Estrada, Naranjo y Ochoa, 2006; Van Bael, Bichier y Greenberg, 2007), de tal manera que cuanto mayor sea la diversidad arbórea, mayor será la diversidad biológica en general. Si bien, la alta riqueza arbórea de los SA rústicos de cacao, como los aquí estudiados, no mantiene la misma biodiversidad animal que la vegetación original, esta se asemeja más que los SA no rústicos con baja riqueza de especies arbóreas, o los sistemas tecnificados sin sombra (Perfecto, Rice, Greenberg y van der Voort, 1996; Rice y Greensberg, 2000).
México no sobresale a escala mundial en la producción de cacao (57 841 ha dedicadas a esta actividad hasta 2019, Servicio de Información Agroalimentaria y Pesquera [SIAP], 2017) encontrándose muy por debajo de los países con la mayor producción tales como Costa de Marfil (2 150 000 ha) y Ghana (1 200 000 ha) (Rice y Greensberg, 2000). Sin embargo, con el resultado de este trabajo, es posible afirmar que sobresale como el mejor proveedor de servicios ambientales de conservación de biodiversidad y almace-namiento de carbono, ya que, al practicar el sistema de producción bajo sombra, es de los países con mayor densidad de especies arboladas en el sistema de producción, a diferencia de los otros países que basan su producción en el sistema de pleno sol y en menor proporción de sombra (Tabla 4).
Conclusiones
Los SA de cacao en Tabasco, México presentan una alta diversidad arbórea que favorece el almacén de carbono a medida que sus especies de sombra maduran o son de mayor edad. Esto, indudablemente, constituye un ejemplo de un uso del suelo más adecuado en el trópico húmedo, para la conservación biológica y mitigación del calentamiento global actual, por lo que se hace una enfática recomendación para su conservación y aumento.

