











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El género Eucalyptus se compone de más de 700 especies y muchas de ellas se han distribuido en el mundo, debido a las buenas propiedades de la madera y su rápido crecimiento (Brooker, 2000). En México el eucalipto se introduce desde el siglo XIX pero se cuestiona su impacto en el ciclo del agua, efecto alelopático y sobre la degradación del suelo (Food and Agriculture Organization [FAO], 1995; Ruiz, Rivero, Alcalá y Espinoza, 2006; Vega y Baez, 2016). Sin embargo, el uso de agua por los árboles depende de la tasa de crecimiento y de sus estrategias de defensa ante el estrés (Smith, 2015), siendo entonces relativo el uso del recurso por cada especie (Dvorak, 2012; Smith, 2015).
Después de la radiación, el agua es el principal recurso limitante para el crecimiento vegetal. El agua debe estar disponible en el suelo para ser aprovechada por las plantas, además de los aportes dados por la lluvia, el riego o el manto freático superficial, las características fisicoquímicas del suelo pueden alterar su disponibilidad (salinidad, textura arcillosa, etc). La capacidad de almacenamiento de agua en el suelo está dada por su profundidad, su contenido de materia orgánica, su textura y estructura, entre otros. Por otra parte, una planta es competitiva en la medida que pueda vencer un potencial hídrico con un alto valor absoluto, sea fotosintéticamente eficiente en el uso de agua y sus raíces puedan explorar un gran volumen de suelo (Kramer y Boyer, 1995).
Las buenas prácticas de manejo silvícola son importantes para atender la densidad de plantación, fertilización o cosecha y controlar el efecto negativo sobre la recarga hídrica, erosión y competencia entre especies (Wilkinson, Schofield y Kanowski, 2014). En este sentido, la decisión de plantar una especie forestal en un sitio específico obedece a la compatibilidad entre las características de la especie y el medio biofísico del sitio; que puede comprobarse o investigarse mediante modelos de crecimiento, parcelas de introducción, estudios de caso o la evaluación de la producción forestal.
El crecimiento y uso de agua pueden predecirse usando modelos de productividad forestal basados en los procesos fisiológicos que controlan el crecimiento, como son la transpiración, fotosíntesis, nutrición, fenología, etc. (Landsberg y Waring, 1997). Un modelo es una representación de la realidad, pero al basarse en los procesos, los modelos pueden mejorarse con base en el entendimiento y medición de las variables que determinan los procesos fisiológicos. La utilidad de un modelo radica en seguir representando la realidad cuando cambia el valor de variables de importancia para su funcionamiento, variables denominadas forzantes. Así, un modelo basado en procesos puede brindar estimados con incertidumbre tolerable no solo en el sitio original donde se propuso, sino en condiciones diversas.
En este sentido, el modelo 3-PG (Physiological Principles in Predicting Growth) es un modelo basado en procesos, usado para el manejo forestal, es simple y de libre acceso, se ha aplicado a especies de Pinus (Bryars et al., 2013; Chan et al., 2018), pero particularmente se ha enfocado a eucaliptos; Eucalyptus globulus y E. grandis (Sands, 2004). El crecimiento de E. globulus se ha calibrado en 3-PG como función de variables alométricas, estructura y procesos del dosel, tasas de conversión de energía, conductancia y respuesta estomática, entre otras (Sands y Landsberg, 2002). Además, el modelo 3-PG utiliza variables ambientales para determinar el estado inicial del sistema y entonces predice el crecimiento y la función de la plantación hasta el fin del turno (Vega-Nieva et al., 2013).
Este modelo es atractivo porque necesita menos datos en comparación con otros modelos, realiza cálculos sobre la producción primaria usando un submodelo biofisiológico de tipo mecanístico y también usa relaciones empíricas, para asignar el carbono a las reservas de biomasa (aérea o subterránea), dependiendo de las condiciones ambientales de crecimiento (Almeida, Landsberg y Sands, 2004; Almeida et al., 2004).
Por otra parte, los modelos empíricos de crecimiento y producción son una herramienta tradicional para el manejo forestal, porque usan datos sencillos para medir la biomasa (Zianis y Mencuccini, 2004). Sin embargo, la capacidad predictiva de estos modelos disminuye cuando las condiciones ambientales o el manejo cambian, porque se basan en el supuesto de estabilidad climática y ambiental (Almeida et al., 2010). Por simplicidad, pueden incorporarse relaciones empíricas dentro de un modelo basado en procesos, con lo que se tendría un modelo híbrido.
Cada herramienta de modelación ofrece una representación del desempeño o función, resultando atractivo combinar las predicciones de manera consensuada y probabilística (Araújo y New, 2007). La combinación o ensamble de modelos se usa en distintas circunstancias, por ejemplo, para predecir el área de distribución potencial de una especie o para la predicción de la temperatura global debido al cambio climático (Klesse et al., 2018; Pearson y Dawson, 2003). Al valorar escenarios múltiples y ponderarlos en cuanto a su probabilidad de ocurrencia se reduce la incertidumbre (Beaumont, Hughes y Pitman, 2008). No obstante, cualquier modelo difiere de la realidad y, en la mayoría de los casos, es posible obtener modelos competitivos en su habilidad para representar aspectos claves del sistema (Araújo y New, 2007).
Objetivos
En el presente estudio la hipótesis fue probar el modelo 3-PG como representación de variables medidas en una plantación de E. globulus durante tres años en Jalisco, México. Un primer objetivo fue determinar el efecto de la humedad mínima del suelo sobre la modelación de la biomasa de E. globulus con el modelo 3-PG. Un segundo objetivo fue considerar la variabilidad ambiental anual y generar un ensamble de modelos, cada modelo basado en datos ambientales medidos en distinto año y comparar contra datos observados del diámetro a la altura del pecho.
Materiales y métodos
Sitio de estudio
El trabajo se realizó en el Rancho las Agujas en el municipio de Zapopan, Jalisco (N 20° 45' 39.6'', W 103° 32' 33.5'' y 1640 m de altura sobre el nivel del mar). El clima es subtropical, con lluvias en verano con una media anual de 816 mm, temperatura media de 23.5 grados °C, máxima de 35 °C y mínima de 5.4 °C. De 2007 a 2009 se realizaron mediciones en dos bloques de pastizal de Chloris gayana (Kunth) de 18 ha cada uno plantados con E. globulus en el 2004. La densidad de plantación fue de 950 ha-1 a un espaciamiento de 3 m × 3.5 m. En marzo del 2007 el rodal se caracterizó por una altura de 7 m ± 1 m, diámetro a la altura del pecho (DAP) de 13.4 cm ± 7 cm, área basal de 13.4 m2 ha-1 y mortalidad de 3%. La pendiente del sitio fue menor a 2%, los suelos profundos, de textura migajón-arenosa, sin evidencia de erosión y clasificados como Feozem háplico asociado a Regosol eútrico. En el estrato 0 mm - 75 mm el contenido de materia orgánica fue de 1.5%, el pH de 5.2 y la densidad aparente de 0.9 Mg m3. En el sitio se encontró un pozo artesiano con un espejo de agua a 15 m, considerando ésta la profundidad del manto freático, quedando fuera del alcance de los eucaliptos.
Variables medidas
Se midió dos veces al año el DAP (cm) y el Índice de Área Foliar (IAF), se muestrearon los individuos en tres parcelas de 380 m2 en cada bloque (n = 6). No se consideraron en la modelación los árboles defectuosos, con diámetro a la altura del pecho menor a 8 cm o con bifurcaciones. El IAF del eucalipto se midió con un analizador de dosel LI-2000 (Li-cor Inc.). Las mediciones se realizaron en hora crepuscular en cada una de las parcelas obteniéndose tres mediciones en un transecto de 1.5 m perpendicular a las hileras de árboles (n=6). El registro de referencia se obtuvo con otro LI-2000 ubicado fuera de la influencia de los árboles y con registro automático. Los valores de IAF obtenidos con el LI-2000 se ajustaron con un factor de 1.51 (Battaglia, Cherry, Beadle, Sands y Hingston, 1998).
Las variables ambientales se midieron dentro de la plantación y lejos de la influencia de los árboles. Con multisensores WXT510 (Vaisala Inc.) se registró cada 20 minutos la precipitación pluvial acumulada (P), humedad relativa (Hr), presión atmosférica (Pa), temperatura del aire (Ta), velocidad y dirección de viento (Vv y Dv, respectivamente). La radiación se midió con un piranómetro LI 200X (Campbell Scientific Inc.). La evapotranspiración potencial (ET0) en 2007 y 2008 se estimó usando la ecuación Penman-Monteith (Allen, Pereira, Raes y Smith, 1998). Cada 20 minutos se registró el contenido de agua del suelo (θ, % v/v) con sondas de reflectometría modelo CS-216 (Campbell Scientific Inc.) instaladas horizontalmente a 0.1 m, 0.2 m, 0.4 m, 0.8 m, 1.2 m y 1.8 m de profundidad. El contenido de θ como lámina (mm) se integró en función del volumen del suelo en el estrato 0 m - 1.8 m y se calculó el cambio diario en θ (Δθ).
La evapotranspiración del cultivo (ETc) se determinó con un balance hídrico para un intervalo de tiempo igual a un mes: P = ETc ± Δθ, donde P es la precipitación (mm). El cálculo se realizó de abril del 2007 a marzo de 2008. La profundidad del perfil se determinó tomando la experiencia de Almeida, Soares, Landsberg y Rezende (2007) para E. grandis de siete años de edad en donde la extracción de agua fue hasta 1.6 m, según sus mediciones con sonda de neutrones. En el balance se consideraron como mínimas las contribuciones del drenaje y escurrimiento superficial. En este balance se incluyeron las pérdidas por intercepción de la lluvia debidas al dosel del eucalipto y del sotobosque de pasto. El coeficiente del cultivo (kc) se calculó como el cociente de ETc / ET0 y el índice de humedad climática (IHC) como P/ ET0 (White, Battaglia, Ren y Mendham, 2016).
Modelación
El modelo 3-PG usa como condiciones iniciales para la simulación: a) datos del estado del sitio, b) datos de inicialización para la plantación y c) datos climáticos. Los valores de los parámetros utilizados en la modelación fueron los definidos por defecto en 3-PG para E. globulus. El resultado de la modelación son predicciones de distintas variables durante un periodo de crecimiento a intervalos mensuales o anuales. Las variables modeladas fueron DAP, IAF, ET0, Incremento Medio Anual (IMA), biomasa total, aérea y de raíces. Estas variables fueron comparadas con las mediciones en campo para explorar la capacidad del modelo 3-PG. Finalmente, la biomasa se multiplicó por un factor de 0.5 para estimar el contenido de carbono (Návar, González y Graciano, 2005).
Datos de estado del sitio
Las variables para caracterizar el estado del sitio son la latitud, la fertilidad del suelo, la textura del suelo, la humedad máxima y mínima del suelo (θmax y θmin, mm). La fertilidad del suelo (FR) es un atributo cualitativo con valores de 0 a 1 y ponderado de acuerdo con limitantes en: fertilidad (0.4), agua (0.2), oxígeno (0.1), manejo (0.2) y topografía (0.1, Almeida et al. 2010). Para el sitio de estudio se valoraron las limitantes como: FR= 0.4 × 0.8 + 0.2 × 0.4 + 0.1 × 1 + 0.2 × 0.8 + 0.1 × 1 = 0.76. Los valores de θmax y θmin se obtuvieron con los valores extremos para la lámina de agua contenida en el estrato 0 m - 1.8 m durante el periodo de tres años.
Datos de inicialización
Las variables de inicialización de la plantación son: la especie de árbol, la fecha de plantación, la fecha inicial de modelación, la duración de la rotación, la densidad de plantación, la humedad del suelo al inicio de la simulación (θini, mm), la biomasa aérea y de raíces. Los modelos se diseñaron para cada año por lo que la fecha de inicio pudo ser enero de 2007, 2008 o 2009. La fecha de plantación fue agosto del 2004 y la duración de la rotación se estableció en 15 años. Como valores de θini se consideró la lámina de agua contenida en el estrato 0 m - 1.8 m del mes de enero, febrero o marzo con el fin de representar distintos escenarios de humedad remanente del año anterior. Debido a que las lluvias se presentan de junio a octubre, la humedad del suelo en el mes de marzo representaría menor disponibilidad de agua en relación con el mes de enero.
Para el sitio de estudio no fue posible realizar un muestreo destructivo para medir la biomasa aérea o de raíces y por esta razón, los valores de biomasa se obtuvieron con funciones alométricas y escalando a unidad de área (Mg ha-1). En la literatura existen distintos modelos de regresión para determinar la biomasa (kg) en función del DAP, área basal, altura, densidad de la madera o alguna combinación de estos parámetros (Zianis y Mencuccini, 2004). Los modelos en función del DAP han explicado más de 90% de la variación en la biomasa en el género Eucalyptus, pero en ocasiones se ha usado la altura y la densidad de madera para mejorar la regresión; aunque la variación explicada aumenta poco (Eamus, Burrows y McGuinness, 2000). El valor de otras variables en la relación entre la biomasa y diámetro a la altura del pecho es marginal, sobre todo cuando se considera el costo y esfuerzo para obtener datos adicionales al DAP.
La función más común para representar la biomasa es de la forma B = a × (DAP) b , donde B es la biomasa, a y b son coeficientes de escalamiento (Zianis y Mencuccini, 2004). La transformación doble logarítmica en la relación entre la biomasa y DAP se utiliza con frecuencia para resolver problemas de homogeneidad en la varianza de la biomasa o DAP, pero para la transformación inversa y obtener estimados certeros de la biomasa, se necesita realizar ajustes en función de la varianza (Beauchamp y Olson, 1973). Senelwa y Sims (1999) concluyen que no hay diferencia entre especies de Eucalyptus en el modelo alométrico de biomasa basada en el DAP de plantaciones jóvenes, aunque la relación específica en la tasa de crecimiento implica una relación variable entre la biomasa y DAP conforme crece el árbol (Zianis y Mencuccini, 2004). Con estas consideraciones se usaron las relaciones propuestas por Bennett, Weston y Attiwill (1997) para los componentes de la biomasa aérea de E. globulus en plantaciones de edad similar a la examinada en el presente estudio. Para calcular la biomasa de las raíces mayores a 2 mm se aplicó la función; biomasa de raíces = 5.97 × DAP2.8068 × 10-3, usada por Forrester, Bauhus y Cowie (2006) para E. globulus.
Datos climáticos
Las variables climáticas necesarias son valores mensuales para radiación neta (Mj m2 d-1), déficit de presión de vapor (mBar), precipitación pluvial (mm), número días con helada, temperaturas mensuales promedio (°C): diaria (T), mínima y máxima. El déficit de presión de vapor y el número de días de helada, se calcularon con las rutinas de herramienta implementadas en la hoja electrónica de 3-PG. Para cada año se generó una base de datos correspondiente usando los datos medidos de la estación meteorológica del sitio.
Escenarios
Se modeló el desarrollo de la plantación de E. globulus de acuerdo con las variables θmin y θini. Para cada año se utilizó una base de datos climática y la biomasa calculada con el DAP, medido al inicio del año. Los datos climáticos se asumen como constantes durante el desarrollo de la plantación. Para cada año se obtuvieron tres modelos según el valor de θini para representar la incertidumbre de θini en la modelación. Hipotéticamente el crecimiento de la plantación sería proporcional a θini. La magnitud de θini depende de la distribución y variación anual en la precipitación. La simulación del año 2007 posiblemente predeciría mejor desempeño en comparación con 2008 o 2009, si fuera insuficiente el almacenaje de agua del suelo en relación con la mayor demanda evaporativa conforme crecen los árboles jóvenes. Los tres valores de θini se obtuvieron con base en la lámina de agua del suelo promedio mensual de enero a marzo (Tabla 1). También se exploró el efecto de la θmin sobre la biomasa de E. globulus. Se utilizaron cuatro valores de θmin en el intervalo de 110 mm a 170 mm, correspondientes a los valores mínimos de θ observados en 2007 a 2009.
Tabla 1 Contenido de agua del suelo en el estrato 0 m - 1.8 m, usado en el modelo 3-PG para el contenido inicial de humedad del suelo (θini).
| Mes | 2007 | 2008 | 2009 |
| mm | |||
| Enero | 240 | 190 | 229 |
| Febrero | 200 | 150 | 191 |
| Marzo | 187 | 137 | 159 |
Los modelos se evaluaron comparando los valores estimados de DAP, ET e IAF con los promedios medidos en la plantación a diferentes tiempos en función del porcentaje de error relativo (e %, ecuación 1) y el error promedio absoluto (EPA%), donde O son los valores observados, Pr son las predicciones y K el número de observaciones (ecuación 2) (Miehle et al., 2009):
El modelo 3-PG genera predicciones para una variable dada en función del tiempo (años) y de las condiciones iniciales. De esta manera la relación entre las predicciones obtenidas se representó como (ecuación 3):
donde las M funciones simuladas se promedian para la x variables de interés (Zhang, Patuwo y Hu, 1998). Para cada escenario se estimaron los coeficientes de regresión entre el DAP o la biomasa total y la edad de la plantación. Se calculó el criterio de información Bayesiano (BIC (M k )). Se asumió una distribución uniforme para la probabilidad anterior (P(M k ) = 1/3). La probabilidad posterior (P (M k | y)) se calculó como (ecuación 4) (Clyde, 2003):
Cada coeficiente del modelo ensamblado se obtuvo como (ecuación 5):
donde E(β | y, M k ) es el estimado específico para cada modelo.
Resultados
Datos ambientales
La lluvia registrada fue de 1002 mm, 729 mm y 703 mm en los años 2007 a 2009. La variación en la precipitación anual se debió a la lluvia ocurrida de mayo a octubre, correspondiente a 95% de la lluvia (Fig. 1). La lluvia de noviembre a abril resultó en Δθ menor a 0.3%v/v en el estrato más superficial del suelo (0-0.1 m). La ET0 fue de 1235 mm y 1140 mm en 2007 y 2008. Para el periodo de abril del 2007 a marzo del 2018 la P fue de 959 mm, la ET0 de 1303 mm y la ETc de 514.3 mm; siendo para este periodo entonces el kc de 0.39 y el IHC de 0.73. La temperatura promedio mensual fue similar entre los años siendo los meses más calientes los antecedentes a las lluvias (Fig. 1). La temperatura promedio anual en el periodo fue de 18.1 °C. A 1.8 m el θ aumentó de 6.6% a 10.5%v/v del inicio al final de la temporada de lluvia, este contenido de humedad del suelo indicó ausencia de drenaje del perfil examinado.

Humedad del suelo
La modelación de la biomasa de la plantación con 3-PG fue sensible al valor de θmin pero no para θini en los intervalos explorados. En la Figura 2 se muestra el resultado para la biomasa total, la tendencia en el resultado fue semejante para DAP, IMA, IAF y los componentes de la biomasa. Los resultados de la modelación usando las mediciones del 2007 sugirieron una inconsistencia entre el año de la base de datos y θmin. Entre el año 2008 y 2009 la magnitud en diferencia entre la biomasa total modelada se conservó, pero no fue el caso para los estimados basados en el año 2007. De los tres años, el 2007 fue el de mayor precipitación, pero 2008 y 2009 fueron años parecidos en las condiciones ambientales.

Figura 2 Biomasa total predicha para la plantación de E. globulus a cinco años (enero del 2010) usando diferentes valores de humedad del suelo inicial (A) y mínima (B).
A los cinco años el intervalo en biomasa total fue de 103.7 Mg ha-1 a 139.6 Mg ha-1 cuando θmin fue 110 mm o 170 mm, respectivamente (Fig. 2). La profundidad y textura del suelo limitan la capacidad de almacenamiento, pero también la homogeneidad en la distribución de la lluvia es importante para el almacenamiento y mayor θmin. Los suelos del sitio estudiado son planos y profundos y permiten almacenar gran cantidad de agua. Para suelos someros la importancia de θmin puede ser menor, pero también será menor la biomasa producida. Poca agua almacenada en el suelo también es un indicador de condiciones de estrés hídrico. Por estas razones, se decidió utilizar solamente θmin para generar los escenarios y un valor fijo para θini. Para cada año se utilizó el nivel mínimo de θ alcanzado durante el periodo: 110 mm en el horizonte 0 m -1.8 m; este valor resultó en estimados conservadores del desempeño de la plantación (Fig. 2B).
Escenarios
El volumen fustal, IMA y DAP modelados con 3-PG fueron mayores con los datos del 2008 en relación con los otros años (Tabla 2). El mayor volumen con datos del 2008 no se debió al año anterior lluvioso pues el efecto de θini no fue importante (Fig. 1A).
Tabla 2 Volumen fustal (VF), incremento medio anual (IMA) y diámetro a la altura del pecho (DAP) de E. globulus predichos por el modelo 3-PG.
| 2007 | 2008 | 2009 | |||||||||
| Edad (años) | VF (m3 ha-1) | IMA (m3 ha-1 año-1) | DAP (cm) | VF (m3 ha-1) | IMA (m3 ha-1 año-1) | DAP (cm) | VF (m3 ha-1) | IMA (m3 ha-1 año-1) | DAP (cm) | ||
| 4 | 94 | 21 | 15 | 119 | 27 | 16 | 106 | 24 | 15 | ||
| 8 | 188 | 22 | 19 | 233 | 28 | 21 | 197 | 23 | 20 | ||
| 12 | 283 | 23 | 23 | 350 | 28 | 25 | 290 | 23 | 23 | ||
| 15 | 354 | 23 | 25 | 439 | 28 | 28 | 360 | 23 | 25 | ||
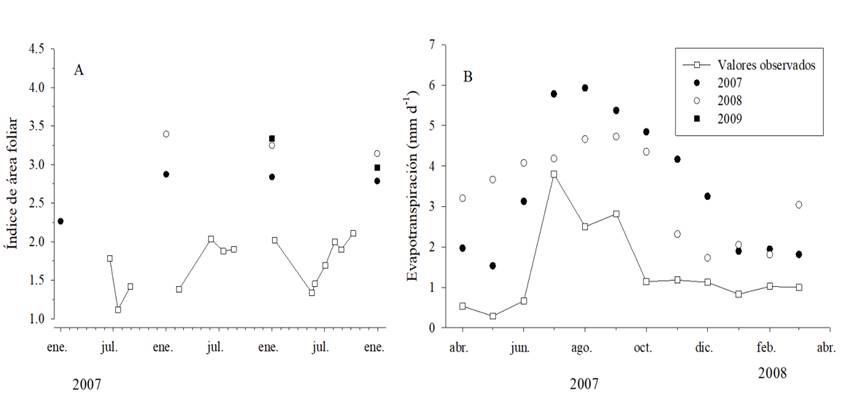
En la Figura 3 se presentan las series de tiempo para DAP y la biomasa total; el e% para el DAP fue de 4.8%, 0.4% y 2.5% para el 2007 al 2009 y el EPA% fue de 5.5%, 2.8% y 2.5 %, respectivamente. El BIC para los modelos fue de 1.8, 2.4 y -0.7 para el 2007 al 2009. A juzgar por los errores para el DAP, los mejores modelos fueron los del 2008 y 2009 pero la biomasa total y DAP a los 15 años fueron similares entre los modelos del 2007 y 2009 (Fig. 3). Las representaciones del IAF y la ET por 3-PG no guardaron relación con las mediciones realizadas con el LI-2000 o con el balance hídrico de 2007-2008 (Fig. 4).

*DAP: Diámetro a la altura del pecho.
Figura 3 Predicciones de diámetro a la altura del pecho (A) y biomasa total (B) utilizando mediciones climáticas de los años 2007, 2008 o 2009 para la inicialización del modelo 3-PG.
Ensamble
Conceptualmente, un modelo de ensamble es deseable, pues los escenarios se basan en datos ambientales obtenidos en cada año y el modelo 3-PG asume que permanecen constantes durante el turno. El modelo de cada año resultó en una sobreestimación, pero el modelo ensamblado se acercó más a la relación 1:1 (Fig. 5). La validez del presente estudio recae en la habilidad del modelo 3-PG para modelar distintas variables y por eso se comparó con las medidas del IAF y la ET; sin embargo, el resultado no fue bueno. Sobre la modelación de estas variables con 3-PG se ha enfocado a validar del modelo respecto del DAP y la biomasa con buenos resultados (Moreira de Oliveira et al., 2018), para el balance de agua se ha modificado el cálculo para mejorar la estimación (Silva, Ribeiro, Neves, Barros y Leite, 2013). Con estas consideraciones se realizó un ensamble para la biomasa total, asumiendo una capacidad del modelo 3-PG para representar la biomasa semejante a la presentada para el DAP en la Figura 5.

Figura 5 Relación entre los valores de diámetro a la altura del pecho (DAP) observados y los representados por los escenarios y el ensamble del modelo 3-PG.
La probabilidad posterior para los escenarios de biomasa total de 2007 a 2009 fue 0.19, 0.15 y 0.66, estos valores se usaron para obtener el promedio ponderado sobre todos los escenarios y teniendo más peso según el ajuste de cada escenario. El modelo ensamblado para la biomasa total fue y = -153.07 + 112.64 x 0.5 donde x es la edad de la plantación (Fig. 3). La plantación podría cosecharse para celulosa a partir del año ocho de edad; en ese momento la predicción fue de 82.7 megagramos de carbono por hectárea, de los cuales 23.1 Mg ha-1 son de raíces y 59.5 Mg ha-1 de la biomasa aérea. Considerando un contenido de 58% de carbono en la materia orgánica del suelo, se estimaron 5.87 Mg ha-1 de este elemento en el estrato 0 mm - 75 mm; a mayor profundidad, la contribución a la poza de carbono está dada principalmente por la biomasa de raíces. Tomado en cuenta 5.7 Mg ha-1 de biomasa producida por C. gayana durante la temporada de crecimiento (Guevara-Escobar, Cervantes-Jiménez, Suzán-Azpiri, González-Sosa y Saavedra, 2012) y suponiendo 40% de carbono, la poza sería 2.28 Mg ha-1 para el sotobosque. De esta forma el total de carbono en el sistema fue del orden de 90.85 Mg ha-1.
Discusión
La modelación de la plantación con 3-PG fue sensible al valor de θmin pero no para θini en los intervalos explorados. Este resultado fue acorde al análisis de sensibilidad de modelos 3-PG para E. grandis (Esprey, Sands y Smith, 2004). El efecto de la humedad mínima del perfil de suelo sobre la biomasa a cinco años fue importante y por eso se necesitan datos precisos de la humedad mínima. El valor de IHC fue de 0.73 y el intervalo de 0.7 a 1.0 es indicativo de una temporada seca pronunciada con ET0 excediendo a P durante varios meses; en estas circunstancias, la evapotranspiración se pude sobreestimar, si no se consideran los cambios en θ y el acompañante aumento de la resistencia del dosel (White et al., 2016). La θmin es un parámetro que necesita instrumentación como las sondas usadas aquí (reflectrometría de dominio de tiempo); sin embargo, la medición con sensores remotos y asimilación de datos puede ser una fuente de información importante (Entekhabi et al., 2014).
La ETc se mantuvo por debajo de 1 mm d-1 con excepción de los meses de julio a septiembre con valores de 3.8 mm d-1, 2.5 mm d-1 y 2.8 mm d-1 (Fig. 4b). Los valores observados de ETc estuvieron dentro del intervalo registrado en otros estudios (de Almeida y Riekerk, 1990; Maier et al., 2017; Moreaux, O’Grady, Nguyen-The y Loustau, 2013; Silva et al., 2013) y en concordancia con una distribución estacional marcada en respuesta a la precipitación pluvial y variación del área foliar (Almeida et al., 2007; Moreaux et al., 2013).
Posiblemente los valores altos de biomasa modelados para 2008 fueron resultado de un sesgo en la medición del DAP, pues el error estándar de la media fue mayor en este año (0.17, 0.44 y 0.15, para 2007, 2008 y 2009). No obstante, la combinación de variables ambientales pudo influir en la conversión de energía representada por 3-PG. En este sentido, Miehle et al. (2009) señalan que diferentes condiciones y tiempos durante la rotación del cultivo inciden en mayores errores de 3-PG para la modelación de biomasa aérea de E. globulus, en comparación con otras herramientas de modelación.
Las relaciones alométricas determinadas por Bennett et al. (1997) para E. globulus se han utilizado en varios estudios para obtener estimados de la biomasa en función del diámetro a la altura del pecho (Bauhus, Van Winden y Nicotra, 2004; Forrester, Bauhus y Khanna, 2004), pero en el presente estudio posiblemente se sobreestimó la biomasa de follaje utilizada como valor inicial en 3-PG y también resultó en valores elevados del índice de área foliar. Estos aspectos necesitan revisarse obteniendo muestreos destructivos de los componentes de la biomasa. En este aspecto, la biomasa total y el diámetro a la altura del pecho son representados bien por 3-PG (Moreira de Oliveira et al., 2018), pero la estimación del modelo es pobre para otras variables (ET e IAF), posiblemente debido a una combinación de factores (Almeida et al., 2010).
El IAF medido fue bajo, en comparación con los valores de 3 a 6 para E. globulus registrados en Australia; en donde los valores disminuyen en proporción inversa al déficit hídrico y temperatura, llegando al valor óptimo cerca de los 13 °C (Battaglia et al., 1998). Para el sitio de estudio, el IAF medido fue menor al modelado por 3-PG posiblemente por el efecto de mayor temperatura y los seis meses de temporada seca. Si el efecto de déficit hídrico fuera el único implicado, el IAF del sitio de estudio debió aumentar durante las lluvias después de la sequía, de manera parecida a la observada para plantaciones de Eucalyptus en Brasil (Almeida et al., 2010). Posiblemente, las ecuaciones de ajuste obtenidas en zonas templadas de Australia no son representativas de las condiciones de Jalisco, debido a una arquitectura foliar diferente, ocasionada por las condiciones subtropicales secas (Maseda y Fernández, 2016). No obstante, en el sitio la temperatura promedio anual se encontró en el límite superior del intervalo 10 °C - 18 °C para la producción óptima del dosel de eucalipto, en términos de IAF (Battaglia et al., 1998).
Para explicar la discrepancia entre los valores observados y las predicciones de IAF se debe considerar la distribución del follaje y la transferencia de la radiación en el género Eucalyptus como procesos reguladores de la fotosíntesis y la transpiración durante la sequía (Whitehead y Beadle, 2004). El LI-2000 tiene dificultades para evaluar la agregación de los elementos del follaje y la inclinación vertical de las hojas de E. globulus (Macfarlane et al., 2007). Aunado a esto, la inclinación del follaje aumenta durante la sequía como mecanismo de adaptación (Whitehead y Beadle, 2004). Aunque se utilizó la corrección para el LI-2000 en rodales de eucalipto (Battaglia et al., 1998), los valores de IAF medidos fueron distintos a los estimados por 3-PG. Macfarlene et al. (2007) encontraron una relación diferente entre el IAF determinado destructivamente y las mediciones del LI-2000, pero al ajustar los datos del presente estudio con esa ecuación, resultó en valores aún más bajos de IAF en comparación con la ecuación de regresión de Battaglia et al. (1998).
Los estimados de evapotranspiración generados por el modelo 3-PG dependen directamente del IAF, pero también resultaron diferentes debido a la metodología usada en 3-PG para calcular el balance hídrico. Valente, David y Gash (1997) observaron valores de intercepción de la lluvia de 10.8% y una relación intercepción/índice de área foliar de 3.4 para una plantación de E. globulus en Portugal, donde la precipitación fue de 800 mm al año. El modelo 3-PG utiliza dos valores para incorporar el efecto de la intercepción de la lluvia, uno es la proporción máxima de precipitación interceptada y el segundo es el umbral de IAF para la máxima intercepción de la lluvia; los valores usados para la parametrización de E. globulus en 3-PG fueron 0.15 y 0.0; sin embargo, estos valores no imponen una restricción para alcanzar la máxima intercepción de lluvia en función del índice de área foliar. La representación del proceso de intercepción en 3-PG es simple y no considera los efectos de duración e intensidad de la lluvia, además de la capacidad de almacenamiento de agua del dosel (Crockford y Richardson, 2000).
El error de 3-PG en las predicciones en plantaciones jóvenes de Eucalyptus también se atribuye a la dificultad de simular la intercepción de la lluvia y la fotosíntesis antes del cierre del dosel (Miehle et al., 2009). De manera más importante, en el modelo 3-PG no hay consideración para la vegetación asociada del sotobosque para calcular la evapotranspiración. Así, el valor de evapotranspiración obtenido con el balance hídrico del suelo representó las pérdidas de agua por la vegetación, ya fuera el eucalipto o el sotobosque de pasto; la incorporación de un módulo en 3-PG para las interacciones entre especies forestales y del sotobosque sería útil para resolver la contribución del pasto a la ET (Luedeling et al., 2016). Por último, las mediciones del contenido de humedad obtenidas brindan un balance hídrico respecto de un estrato del suelo (0 m - 1.8 m) y no se puede descartar la extracción de agua a mayor profundidad. Al respecto, el contenido de agua a 1.8 m de profundidad aumentó hasta 25%v/v, valor inferior al de saturación del suelo (Guevara-Escobar et al., 2012), pero el movimiento de agua en el suelo es complejo y los flujos preferenciales a mayor profundad no pueden descartarse.
El carbono estimado para la biomasa aérea de eucalipto fue de 59.5 Mg ha-1, este dato es comparable con lo publicado por Miehle et al. (2009), quienes encontraron 45 Mg ha-1 en la biomasa aérea para el promedio de sitios del sur de Australia con buena productividad, aunque algunos sitios alcanzaron 79 Mg ha-1 y 125 Mg ha-1. En plantaciones de eucalipto en la costa atlántica de Brasil el carbono almacenado en la madera fue de 33.5 Mg ha-1 a 47.3 Mg ha-1 para sitios clasificados de baja productividad y en sitios con alta productividad fue de 57 Mg ha-1 a 102 Mg ha-1 (Stape, Binkley y Ryan, 2004). Para el trópico húmedo de México, Seppänen (2002) establece escenarios optimistas para plantaciones de Eucalyptus a los siete años de edad donde el carbono almacenado en la plantación, sotobosque y suelo es de 320 Mg ha-1 - 610 Mg ha-1.
Conclusiones
Los resultados mostraron la utilidad de las relaciones alométricas y 3-PG para modelar el desarrollo de E. globulus fuera del contexto ambiental en el que se generaron. La modelación del índice de área foliar y la evapotranspiración son importantes para el conocimiento fisiológico de la población arbórea y su representación en 3-PG puede mejorarse incorporando ajustes adecuados para la estimación indirecta del índice de área foliar, balance hídrico con partición de los componentes vegetales y modelos de la intercepción de lluvia. La relevancia de la humedad mínima del suelo en el modelo 3-GP se mostró con valores observados en campo. El ensamble de escenarios mostró un mejor desempeño para la modelación del diámetro a la altura del pecho. El sitio se describió como tropical seco por el IHC de 0.73, se consideró conservador el uso de agua del eucalipto por su kc de 0.39 y, sin embargo, como de alta productividad por los 90.85 Mg ha-1 almacenados en el sistema.

