











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
Erythrina americana es una leguminosa arbórea de la familia Fabaceae que se utiliza como parte de los cercos vivos que limita una pradera en las unidades de producción pecuaria localizadas en la región tropical (Llera Zavala y Mélendez Nava, 1994; Grande Cano, Villanueva López, Maldonado García y Hernández Daumás, 2013). El follaje de E. americana es de importancia en la alimentación de pequeños rumiantes, debido a que representa una fuente de carbohidratos estructurales y de proteína cruda. El follaje de esta planta tiene alta aceptación y palatabilidad, por lo que es consumido con facilidad por los ovinos y puede incluirse en la dieta de estos hasta en 30% sin que se afecte el estado de salud de los animales (Grande Cano, 2010; Best et al., 2017). En adición, esta leguminosa puede producir follaje todo el año cuando es sometida a un proceso de poda controlada (Meléndez Nava, 2003), lo que representa una gran ventaja en la alimentación de rumiantes por la disponibilidad y calidad nutricional de la planta.
Estudios previos en E. americana, indican que la edad de rebrote del follaje afecta su rendimiento de materia seca (MS), encontrándose el mayor rendimiento de MS a una edad de rebrote de 90 días y 120 días (Meléndez Nava, 2003). Sin embargo, se desconoce la influencia de la edad de rebrote sobre la composición química, contenido de polifenoles y de taninos en el follaje. Los taninos son metabolitos secundarios de las plantas, su presencia en el follaje ha despertado interés en la alimentación de rumiantes debido a su capacidad de unirse y precipitar proteínas solubles y carbohidratos, situación que pudiera representar una ventaja al proteger a la proteína de la degradación ruminal, siempre y cuando la proteína sea liberada para su degradación en el abomaso e intestino delgado (Frutos, Hervás, Giráldez y Mantecón, 2004; Naumann, Tedeschi, Zeller y Huntley, 2017) y la concentración de estos metabolitos no excedan los 50 g kg-1 de MS (Li, Tanner y Larkin, 1996). Particularmente, en las hojas de E. goldmanii se ha detectado la presencia de fenoles totales (6 g kg-1 MS) y taninos condensados, 16.3 g kg-1 MS, (Pinto, Ramírez, Ku Vera y Ortega, 2002; Pinto et al., 2003). Además, en rumiantes existen evidencias que indican que el consumo de follaje con taninos condensados puede contribuir a controlar a los nematodos gastrointestinales (Hoste et al., 2015; Borges y Borges, 2016).
Objetivos
El objetivo del estudio fue determinar la influencia de la edad de rebrote de E. americana sobre el contenido de compuestos fenólicos y la composición química en el follaje.
Materiales y métodos
Localización
El estudio se realizó durante el período de noviembre de 2016 a marzo de 2017 en la Unidad Experimental Ovina del Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias (Inifap), localizada en el municipio de Huimanguillo, Tabasco, México (17° 50’ LN, 93° 23’ LO), con clima cálido húmedo, lluvias todo el año (Af) y temperatura ambiente media anual de 27.8 °C (Instituto Nacional de Estadística y Geografía [Inegi], 2017).
Árboles, poda y follaje
Se utilizaron 24 árboles de E. americana que forman parte del cerco perimetral de las praderas manejadas para el pastoreo de ovinos. Los árboles tenían en promedio (± DE) 1.6 m ± 0.1 m de altura, una distancia entre árboles de 1.0 m ± 0.3 m y un diámetro a los 0.2 m y 1.3 m de altura del suelo de 5.8 cm ± 1.4 cm y 4.1 cm ± 1.0 cm, respectivamente. Los árboles se podaron antes de iniciar el estudio con el fin de obtener uniformidad en la edad de rebrote. La poda se realizó con pinzas de podar e implicó el corte de todas las ramas.
Edad de rebrote
Las edades de rebrote del follaje estudiadas fueron 60 días, 90 días y 120 días. Los días de las edades del rebrote se contaron a partir del corte de uniformidad aplicado el 21 de noviembre de 2016.
Composición química del follaje
En cada edad de rebrote se obtuvieron cuatro muestras compuestas de follaje (hojas y peciolos sin incluir tallos tiernos). Cada muestra compuesta estuvo formada por el follaje de dos árboles. Las muestras se secaron a 50 °C. Posteriormente, el material fue molido en un molino (Thomas-Wiley®) con malla de 1 mm. Se determinó por duplicado MS, cenizas, materia orgánica y proteína cruda con métodos de la AOAC (Latimer Jr., 2012); fracciones de fibra detergente neutro y fibra detergente ácido con las técnicas de Van Soest, Robertson y Lewis (1991).
Compuestos fenólicos
Para la determinación de la concentración de compuestos fenólicos se utilizaron seis árboles por edad de rebrote. Cada árbol se consideró una repetición. El follaje de cada árbol, por edad de rebrote, se secó a 50 °C, se trituró, pasó a través de una malla de 1 mm, en molino Wiley, y se conservó en oscuridad a 4 °C hasta el análisis. Las variables evaluadas fueron las concentraciones de polifenoles totales, fenoles no taninos, taninos condensados, hidrolizables y totales. Todos los análisis en el laboratorio se realizaron por triplicado.
Polifenoles totales. Las muestras se desengrasaron con éter de petróleo (Múzquiz, Burbano, Cuadrado y De la Cuadra, 1993). Los polifenoles se determinaron con el método de Folin-Ciocalteu, y ácido gálico como estándar en muestras libres de grasa. La absorbancia se midió a 765 nm en un espectrómetro (Thermoelectron, Genesys 10 UV) usando ácido gálico (0.5 mg mL-1) para preparación de estándares (Makkar, Blummel, Borowy y Becker, 1993).
Fenoles no taninos. A las muestras libres de grasa se les adicionó polivinilpirrolidona para separar fenoles taninos de los fenoles no taninos, después de agitarlas en vórtex se incubaron por 15 min en oscuridad a 4 °C y se centrifugaron (10 000 rpm) a 25 °C por 10 min (Makkar et al., 1993). En el sobrenadante se cuantificó la concentración de fenoles no taninos con el reactivo de Folin-Dennis a 725 nm. Los estándares se prepararon a partir de una solución estándar de 0.5 mg mL-1 de ácido gálico.
Taninos condensados (proantocianidinas). Para cuantificar los taninos condensados, muestras libres de grasa se extrajeron con metanol al 80%; a 500 μL de ese extracto se le agregaron 3 mL de butanol: HCl (95:5, v/v) y 100 μL de reactivo férrico (sulfato de amonio férrico a une concentración de 2% en HCl 2 N), se cubrieron y mantuvieron en baño María a ebullición por 60 min y se dejaron enfriar a temperatura ambiente. Un blanco preparado con butanol: HCl y sin calentar se incluyó (Porter, Hrstich y Chan, 1985). La absorbancia se midió a 550 nm. La concentración de taninos condensados se calculó como leucocianidina equivalente con la ecuación:
Donde:
TC = Taninos condensados (g kg-1 MS)
A = Absorbancia (nm)
78.26 = factor de corrección
MS = materia seca (%)
El coeficiente extinción molar (E 1%, 1 cm, 550 nm) de leucocianidinas es de 460.
Taninos totales e hidrolizables. Los taninos totales se calcularon de la diferencia de fenoles no taninos con polifenoles totales. Los hidrolizables fueron estimados como la diferencia de taninos totales y condensados (Singh, Sahoo, Sharma y Bhat, 2005; Rana, Wadhwa y Bakshi, 2006).
Condiciones climáticas
Se consultaron los registros climáticos de la estación meteorológica de la Comisión Nacional del Agua [Conagua] (2017) localizada en el Campo Experimental Huimanguillo, Inifap. Con los datos climáticos se determinó el promedio en la temperatura ambiente (mínima, media y máxima, °C), humedad relativa (mínima, media y máxima, %), la precipitación pluvial acumulada (mm) y la radiación solar (Wm-2) desde la fecha de corte de uniformidad hasta la duración de la edad de rebrote.
Diseño experimental y análisis estadístico
Todos los análisis se efectuaron con el paquete estadístico Statistical Analysis System (SAS, 2002). Las variables dependientes fueron: proteína cruda (%), MS (%), cenizas (%), materia orgánica (%), fibra detergente neutro (%) y fibra detergente ácido (%), polifenoles totales (g kg-1 de MS), fenoles no taninos (g kg-1 de MS), taninos condensados (g kg-1 de MS), hidrolizables (g kg-1 de MS) y totales (g kg-1 de MS). A los datos se les aplicó la prueba de Shapiro-Wilk’s para probar que tuvieran una distribución normal y la prueba de Levene para probar la homogeneidad de las varianzas. Para el análisis de los datos se utilizó un modelo de efectos fijos, en donde el factor fue la edad de rebrote del follaje. En los datos de composición química del follaje la unidad experimental fue la muestra compuesta de follaje. Se utilizaron cuatro unidades experimentales por edad de rebrote. En los datos de fenoles y sus fracciones la unidad experimental fue el árbol. Se utilizaron seis unidades experimentales por edad de rebrote. Los análisis se realizaron con el procedimiento ANOVA. Las medias se compararon con la prueba de Tukey. Los valores de las medias se consideraron estadísticamente significativos cuando P < 0.05. La variable fenoles no taninos no tuvo varianza homogénea y los taninos hidrolizables no mostraron una distribución normal por lo que se analizaron con la prueba de Kruskal-Wallis. Cuando se detectó influencia (P < 0.05) de la edad de rebrote sobre la variable de respuesta, se aplicó el test de la suma de rangos de Wilcoxon para datos no pareados (Milton, 2007).
Resultados
En las Figuras 1 y 2 se describen los valores de temperatura ambiente, humedad relativa, precipitación pluvial y radiación solar durante el período de estudio. El promedio de temperatura ambiente fue similar durante el período de estudio. Sin embargo, la temperatura ambiente mínima disminuyó en 1.3 °C y la máxima aumentó en 2.5 °C, cuando las edades de rebrote fueron 90 días y 120 días con relación a lo detectado en 60 días. A estas edades de rebrote (90 días y 120 días), la humedad relativa promedio y mínima también disminuyeron con relación a lo registrado a los 60 días de iniciado el estudio. La oscilación en la temperatura ambiente y humedad relativa fue mayor en las edades de rebrote 90 días y 120 días con respecto a 60 días. La precipitación pluvial acumulada en las edades 90 días y 120 días representaron 147.2% y 184.5%, respectivamente de la precipitación pluvial registrada a los 60 días.
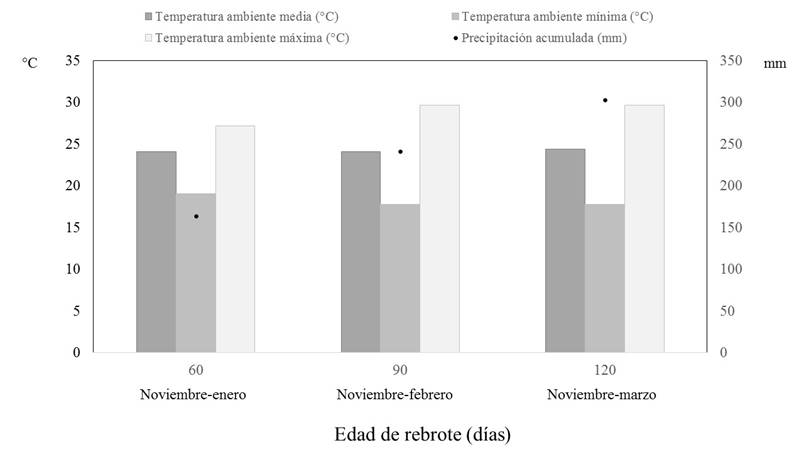
Figura 1 Temperatura ambiente (°C) y precipitación pluvial acumulada (mm) durante el período de estudio considerando la duración de la edad de rebrote.

Figura 2 Humedad relativa (%) y radiación solar (Wm-2) durante el período de estudio considerando la duración de la edad de rebrote.
Los promedios generales (± DE) de proteína cruda, fibra detergente neutro, fibra detergente ácido, MS, cenizas y materia orgánica fueron: 16.2% ± 1.0%, 59.3% ± 7.5%, 41.3% ± 7.8%, 27.6% ± 3.9%, 8.4% ± 0.7%, 91.6% ± 0.7%, respectivamente. Bajo las condiciones ambientales en las que se desarrolló el estudio, la edad de rebrote afectó (P < 0.01) los contenidos de proteína cruda, fibra detergente neutro y fibra detergente ácido. Sin embargo, los contenidos de MS, cenizas y materia orgánica del follaje no fueron influidos (P > 0.05) por la edad de rebrote (Tabla 1).
Tabla 1 Composición química del follaje de Erythrina americana en diferentes edades de rebrote.
| Componente | Edad de rebrote (días) | ||
| 60 | 90 | 120 | |
| Proteína cruda (%) | 15.7b, 1 ± 0.4 | 17.3a ± 0.2 | 15.4b ± 0.2 |
| Fibra detergente neutra (%) | 66.9a ± 3.5 | 52.9b ± 2.1 | 59.1ab ± 2.7 |
| Fibra detergente ácida (%) | 50.4a ± 3.5 | 37.2b ± 1.2 | 36.2b ± 0.5 |
| Materia seca (%) | 26.7 ± 2.9 | 26.2 ± 1.6 | 29.9 ± 0.3 |
| Cenizas (%) | 8.8 ± 1.0 | 8.0 ± 0.7 | 8.2 ± 0.3 |
| Materia orgánica (%) | 91.2 ± 1.0 | 92.0 ± 0.7 | 91.8 ± 0.3 |
a, b, medias ± EE con letras diferentes dentro de la misma línea indican diferencias (P < 0.01).
1, cada valor es el promedio de cuatro muestras compuestas por edad de rebrote.
El mayor contenido de proteína cruda se presentó en la edad de rebrote de 90 días con respecto a 60 días y 120 días. Así mismo, no se detectaron diferencias (P > 0.05) en la proteína cruda del follaje en las edades de rebrote 60 días y 120 días (Tabla 2).
Tabla 2 Fenoles y sus fracciones (g kg-1 de MS) en follaje de Erythrina americana en diferentes edades de rebrote
| Componente | Edad de rebrote (días) | ||
| 60 | 90 | 120 | |
| Polifenoles totales (g kg-1 de MS) | 22.4b, 1 ± 1.3 | 20.7b ± 1.3 | 35.5a ± 2.9 |
| Fenoles no taninos (g kg-1 de MS) | 0.4 ± 0.1 | 0.3 ± 0.0 | 0.3 ± 0.1 |
| Taninos condensados (g kg-1 de MS) | 9.7b ± 0.8 | 6.0c ± 1.0 | 14.2a ± 1.0 |
| Taninos hidrolizables (g kg-1 de MS) | 12.2d ± 1.6 | 14.5d ± 1.1 | 21.0e ± 2.1 |
| Taninos totales (g kg-1 de MS) | 21.9b ± 1.3 | 20.5b ± 1.3 | 35.2a ± 2.8 |
a, b, c, medias ± EE con letras diferentes dentro de la misma línea indican diferencias (P < 0.01).
d, e, medias ± EE con letras diferentes dentro de la misma línea indican diferencias (P < 0.05).
1, cada valor es el promedio de seis repeticiones por edad de rebrote.
El contenido de fibra detergente neutro fue menor (P < 0.01) en la edad de rebrote de 90 días con respecto a 60 días y la fibra detergente ácido fue menor (P < 0.01) a los 90 días y 120 días con relación a los 60 días.
Con excepción de los fenoles no taninos, el resto de las variables fueron afectadas (P < 0.05) por la edad de rebrote (Tabla 2). En el follaje con una edad de rebrote de 120 días se detectó la mayor concentración de polifenoles totales, taninos condensados, hidrolizables y totales con respecto a 60 días y 90 días (P < 0.05).
El contenido de taninos totales a los 60 días y 90 días representaron 62.3% y 58.1%, respectivamente, de los detectados a los 120 días. De manera similar a los taninos totales, los taninos hidrolizables a los 60 días y 90 días fueron 58.2% y 69.0%, respectivamente, de los registrados a los 120 días. Sin embargo, los taninos condensados a los 60 días y 90 días correspondieron 68.5% y 42.2%, respectivamente, de los cuantificados a los 120 días (Tabla 2).
Discusión
Condiciones climáticas
La temperatura ambiente máxima, oscilación térmica y radiación solar promedio incrementaron conforme avanzó el período de estudio, este tipo de cambios reflejan la transición entre el final de la época invernal con el inicio de la primavera. Estas fluctuaciones en las condiciones climáticas durante el desarrollo del estudio corresponden a las de la estación invernal en el estado de Tabasco (Moguel y Molina-Enríquez, 2000; García, 2004; Díaz Padilla, Ruíz Corral, Medina García, Cano García y Serrano Altamirano, 2006) y son consideradas como adecuadas para el crecimiento y desarrollo de E. americana. Esta especie se encuentra distribuida en climas cálidos húmedos, cálidos subhúmedos y cálidos secos, en tierras bajas y con alta elevación (1000-2100 msnm) (García-Mateos, Soto-Hernández y Vibrans, 2001; Comisión Nacional para el conocimiento y uso de la Biodiversidad [Conabio], 2017; Universidad Autónoma de la Ciudad de México [Uacm], 2017).
Composición química
El contenido de proteína cruda en el follaje resultó inferior a lo indicado por Grande Cano (2010), quien encontró 20.1% de proteína cruda en Erythrina spp. sin antecedentes de poda en la región de la Sierra, Tabasco. Sin embargo, en E. americana sin antecedentes de poda en suelos de tipo Fluvisols en Huimanguillo, Tabasco, el contenido de proteína cruda (14.5%) fue ligeramente inferior al registrado en el presente estudio (Palma et al., 2017; Oliva Hernández, López Herrera, Velázquez Jiménez, López Enríquez y Vélez Pérez, 2019). Respecto a las diferencias encontradas en la proteína cruda, en las leguminosas arbóreas se ha encontrado variación atribuida a la especie, posición de la hoja en el tallo, parte de la planta, variación genética dentro de la misma especie, tipo de suelo y clima en el cual crecen (Wood, Stewart y Vargas, 1998; Carmona Agudelo, 2011).
Los valores de fibra detergente neutro y fibra detergente ácido en el follaje resultaron mayores a los indicados en follaje de E. goldmanii y E. indica, sin antecedentes de poda (Pinto et al., 2002, 2003; Ayala Burgos, Capetillo Leal, Cetina Góngora, Zapata Campos y Zandoval Castro, 2006), aunque similares a los observados en follaje de E. americana, sin antecedentes de poda (Oliva Hernández et al., 2019). En leguminosas forrajeras el contenido de carbohidratos estructurales es afectado por la edad de corte y la época del año, detectándose un incremento conforme aumenta la edad de corte y en casos específicos, como en Stylosanthes guianensis, el contenido de fibra detergente neutro no presenta cambios durante la sequía en diferentes edades de corte (García-Ferrer, Bolaños-Aguilar, Ramos-Juárez, Osorio y Lagunes-Espinoza, 2015). Sin embargo, en el follaje de E. americana no se observó un incremento lineal en el contenido de fibra detergente neutro y fibra detergente ácido conforme avanzó la edad de rebrote, debido tal vez a que la relación hoja peciolo disminuyó, lo que llevó a una mayor área foliar a los 90 días respecto a los 60 días.
En leguminosas tropicales de tipo herbáceo y arbustivo no se han detectado cambios en el contenido de fibra detergente neutro por influencia de la edad de corte (Mojica-Rodríguez, Castro-Rincón, Carulla-Formaguera y Lascano-Aguilar, 2017). No obstante, Juárez Lagunes, Montero Lagunes, Alpirez Mendoza, Contreras Jácome y Canudas Lara (2004) mencionan que en estas leguminosas el contenido de fibra detergente ácido se incrementa conforme avanza la edad de corte, lo cual puede estar asociado al proceso de lignificación de las hojas. Por otro lado, en las edades de rebrote de 120 días y 320 días, Choque Durand, Huaita Patiño, Cárdenas Villanueva y Ramos Zúñiga (2018) no detectaron cambios en el contenido de fibra detergente neutro y fibra detergente ácido en follaje de Erythrina sp. Al parecer, el contenido de carbohidratos estructurales en los follajes de especies arbustivas y arbóreas tiene mayor estabilidad con respecto a la mayor parte de las gramíneas debido a que el proceso de lignificación ocurre principalmente en los tallos y no tanto en las hojas (Botero y Russo, 1999).
En los follajes de E. americana, goldmanii e indica, sin antecedentes de poda, se ha detectado un mayor contenido de cenizas a los encontrados en el presente estudio (Pinto et al., 2002; Ayala et al., 2006; Oliva Hernández et al., 2019), lo que puede deberse al efecto de la especie, condiciones edáficas o de manejo donde se desarrollan las especies. Adicionalmente, es probable que la variación en la composición química por edad de las hojas en árboles sin podar sea diferente a la de árboles sometidos a podas a intervalos controlados, lo que explicaría las diferencias en la composición química del follaje entre estudios.
Compuestos fenólicos
Desde el punto de vista de la estructura química, los polifenoles totales comprenden una serie compuestos como las cumarinas, flavonoides, lignina y taninos. Este tipo de compuestos están implicados en las interacciones planta-herbívoro y cumplen diversas funciones, tales como la protección y defensa de la planta frente a predadores y diferentes patógenos (Ávalos García y Pérez-Urria Carril, 2009). El contenido de polifenoles totales se incrementó con la edad de rebrote. Este incremento con la edad de rebrote se observa también en otras leguminosas (García-Ferrer et al., 2015), y puede ser asociado a la fase de crecimiento de la planta (carbono disponible para sintetizar metabolitos secundarios basados en carbono) y a las condiciones ambientales que afectan la fotosíntesis (fuente de carbono) (Bryant, Chapin y Klein, 1983). Las plantas distribuyen más carbono hacia la formación de metabolitos secundarios basados en carbono (por ejemplo, los taninos condensados) cuando estas acumulan carbono en mayor cantidad a las demandas de crecimiento, esto es, durante períodos con baja disponibilidad de nutrientes, alta luminosidad y elevado C02 (Herms y Mattson, 1992; Massad, Dyer y Vega, 2012).
La diferencia en la tendencia del contenido de taninos condensados e hidrolizables en las hojas sugiere que el uso de sustratos en la ruta bioquímica de síntesis del tipo de tanino, condensados e hidrolizables, es influido por la edad de rebrote y probablemente por las condiciones climáticas que imperaron durante el estudio (Makkar, Dawra y Singh, 1991; Estiarte, De Castro y Espelta, 2007; Ávalos García y Pérez-Urria Carril, 2009).
E. americana es un árbol caducifolio que florece durante el inicio de la primavera (García-Mateos et al., 2001), período del año en donde hay alta radiación solar (Fig. 2). Una alta luminosidad e inicio de la floración estimulan la síntesis de taninos condensados (Iason, Hartley y Duncan, 1993; Frutos et al., 2004; Estiarte et al., 2007). Además, Makkar et al. (1991) señalan que un incremento en la edad de madurez de las hojas corresponde con un mayor contenido de taninos condensados. La mayor edad de rebrote de las hojas y la época en que fue cosechado el follaje, transición entre las épocas de nortes y sequía (Fig. 1 y 2), pudieran explicar la mayor concentración de taninos condensados a los 120 días con respecto a 60 días y 90 días.
El contenido de taninos condensados detectado en las diferentes edades de rebrote estudiadas en E. americana se encuentra dentro de los valores registrados en follaje de E. goldmanii y resultó mayor al indicado en E. indica. Para estas dos especies de Erythrina no se estudió la edad de rebrote del follaje (Pinto et al., 2003; Ayala Burgos et al., 2006). En follaje de Gliricidia sepium, leguminosa arbórea de amplio uso en los cercos vivos de la ganadería localizada en el trópico de México (Enríquez Quiroz, Meléndez Nava, Bolaños Aguilar y Esqueda Esquivel, 2011; Grande Cano et al., 2013), se ha encontrado un mayor contenido de taninos condensados (Romero Lara, Palma García y López, 2000) con respecto al detectado en E. americana en el presente estudio. Mientras que Pinto et al. (2002) señalan que el follaje de E. goldmanii contiene mayor cantidad de fenoles totales con respecto a G. sepium. Los resultados anteriores corroboraron que la variación en el contenido de compuestos fenólicos en el follaje de leguminosas arbóreas se atribuye, en parte, a género, especie, tipo de manejo de la poda y suelo en el cual se desarrolla el árbol.
La presencia y cantidad de taninos condensados en el follaje de árboles forrajeros ha adquirido mayor atención debido a que su consumo puede beneficiar o perjudicar el nivel productivo y estado de salud de los pequeños rumiantes (Hoste et al., 2015; Naumann et al., 2017). El consumo de plantas con alto contenido en taninos condensados (> 50 g kg-1 MS) puede reducir el consumo voluntario debido a sus propiedades astringentes (Barry y Duncan, 1984; Waghorn et al., 1994; Frutos et al., 2004). En este sentido, el contenido de taninos condensados detectado en el follaje de E. americana, con edades de rebrote entre 60 días y 120 días, puede ser considerado como intermedio (entre 6 g kg-1 MS y 14 g kg-1 MS) por lo que es posible que el consumo voluntario en pequeños rumiantes no se vea afectado, favoreciendo con ello su empleo como recurso alimenticio en sistemas de producción de tipo silvopastoril (Benavides, 1999; Botero y Russo, 1999).
Con respecto al uso de follajes con alto contenido en taninos condensados para el control de los nematodos gastroentéricos en pequeños rumiantes, estudios realizados in vivo con hojas de Lysiloma latisiliquum, leguminosa de áreas tropicales con alto contenido en taninos (34 g kg-1 de MS), indican que estos permiten reducir la excreción de huevos en heces, la fecundidad de las hembras y pueden generar alteraciones estructurales en la cutícula de hembras adultas del nematodo gástrico Haemonchus contortus, y con ello afectar negativamente su movimiento y nutrición, con posibles consecuencias sobre su reproducción (Ayala Burgos et al., 2006; Martínez-Ortíz-de-Montellano et al., 2010, 2013). En el caso del follaje de E. americana se desconoce si su contenido de taninos condensados permite ejercer un control sobre algún proceso vital de los nematodos gastroentéricos que evite el uso de antihelmínticos para su control; así como, el número de días y cantidad de follaje que deben consumir los pequeños rumiantes para reducir los efectos negativos que les causan este tipo de nematodos.
Conclusiones
La edad de rebrote de E. americana afectó el contenido de proteína cruda, carbohidratos estructurales y compuestos fenólicos en el follaje. A los 90 días se detectó el mayor contenido de proteína cruda con respecto a 60 días y 120 días. El menor contenido de carbohidratos estructurales se presentó entre 90 días y 120 días. Conforme se incrementó la edad de rebrote la concentración de taninos condensados fue mayor. En la edad de rebrote de 90 días, el follaje de E. americana representa una alternativa de complemento alimenticio para pequeños rumiantes manejados en sistemas de producción de tipo silvopastoril debido a su contenido de proteína cruda, carbohidratos estructurales y taninos condensados.

