











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
Las comunidades vegetales características de las zonas áridas y semiáridas de México incluyen a los mezquitales y a distintos tipos de matorrales como el desértico micrófilo, el submontano y el espinoso tamaulipeco (Rzedowski, 1978). De hecho, es común que estos tipos de vegetación se encuentren distribuidos a manera de mosaico en el territorio, dependiendo de las características del terreno y del ambiente. Por consiguiente, es posible encontrar los llamados ecotonos entre los límites de una comunidad y otra (González, 2004).
Dado que las zonas de transición entre matorrales y mezquitales son quizá los ambientes mejor representados en la Planicie Costera del noreste de México, es común que las comunidades campesinas desarrollen sus actividades productivas en estos ecosistemas (Briones y Villarreal, 2001). Esto por el amplio potencial de aprovechamiento que tienen las especies presentes, como forraje, madera, combustible, etc. (Reid, Stafford, Beyer-Münzel y Marroquín, 1990). Y en este sentido, no es raro que las actividades silvopastoriles se desarrollen frecuentemente en zonas de mezquital y matorral (Jiménez et al., 2012; Molina et al., 2013; Pequeño et al., 2012).
Desafortunadamente, el uso antropogénico excesivo o no controlado de estas comunidades vegetales, puede traer como consecuencia el rápido deterioro de los recursos naturales, con la disminución o pérdida posterior de los ingresos económicos de los productores (Arriaga, 2009). Por esta razón, es de suma importancia conocer con detalle el funcionamiento y dinámica ecológicos de estos ecosistemas y, así, planificar adecuadamente su uso y manejo (Alanís et al., 2017). De esta manera, las actividades pastoriles y silvícolas pueden desarrollarse desde un punto de vista sostenible, a la vez que se contribuye también a su conservación (Del Río, Montes, Cañellas y Montero, 2003; Moreno et al., 2010; Sarmiento, Alanís, Mata, Jiménez y Treviño, 2015).
A pesar de que los estudios ecológicos silvopastoriles se llevan a cabo en muchas regiones del mundo (Calle et al., 2013; Fornara et al., 2018), en gran medida se desarrollan con especies tropicales como sucede en Sudamérica y México (Solymosi, Braun, Dijk y Grulke, 2016; Olivares-Pérez et al., 2016; Aryal, Gómez-González, Hernández-Nuriasmú y Morales-Ruiz, 2018). Sin embargo, en las zonas áridas y semiáridas aún existe la necesidad de realizar este tipo de estudios por su importancia y contribución para a un mejor aprovechamiento y conservación de sus comunidades vegetales (Krishnamurthy, Krishnamurthy, Rajagopal y Peralta-Solares, 2017). Por lo anterior, el objetivo de la presente investigación fue caracterizar el componente arbóreo de un sistema silvopastoril en una zona de transición de matorral espinoso tamaulipeco y mezquital en el centro - sur de Nuevo León mediante su composición vegetal y su estructura vertical y horizontal.
Materiales y Métodos
Área de estudio
El estudio ser llevó a cabo en el ejido El Guajolote que se encuentra ubicado en la Carretera Linares - Cd. Victoria km. 132 en el municipio de Linares, Nuevo León (Noreste de México), entre las coordenadas geográficas extremas 24° 40´ 08.46” N y 99° 31´ 14.41” W (Fig. 1). La comunidad estudiada es una zona de transición entre matorral espinoso tamaulipeco y mezquital con un sistema silvopastoril. Posee suelo de tipo vertisol pélico (Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias-Comisión Nacional para el Conocimiento y uso de la Biodiversidad [Inifap-Conabio], 1995), la vegetación es de tipo mezquital (Conabio, 1999), posee un clima tipo (A)C(wo)x´ (García, 1998) y temperatura media semicálida (Vidal, 1990).
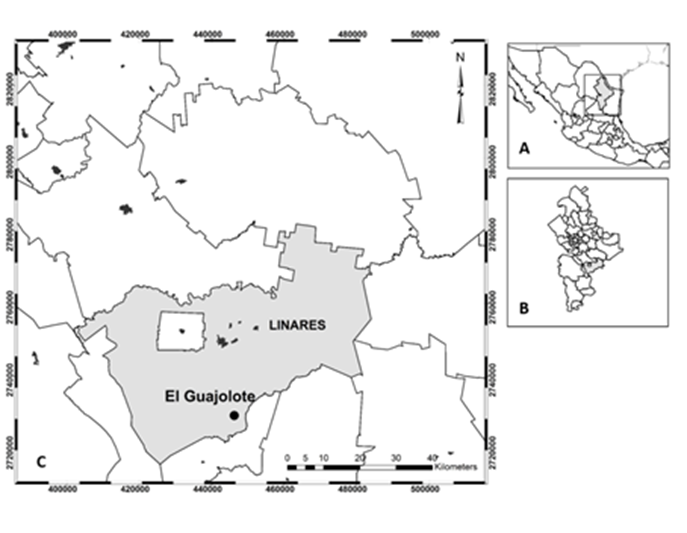
Figura 1 Localización del área de estudio. A) norte de México y sur de Estados Unidos de América, B) estado de Nuevo León, noreste de México y C) ciudad de Linares, ejido El Guajolote.
En el año 2009 se inició el manejo de la comunidad vegetal de este predio privado de 3.9 ha, para el manejo de ganado bovino (Bos taurus L.). El manejo consistió en realizar aclareos por lo bajo, talando los individuos de dimensiones menores a 5 cm de diámetro a la altura del pecho. El objetivo de este tratamiento fue la promoción del mezquite (Prosopis laevigata (Humb. & Bonpl. ex Willd.) M.C. Johnst. y el ébano (Ebenopsis ebano (Berland.) Barneby & J.W. Grimes), para crear espacios abiertos donde se puedan establecer pastos nativos y permitir el desplazamiento del ganado. Los árboles con dimensiones mayores a 5 cm no fueron talados; y en primera instancia se realizó una poda formativa eliminando la parte baja de la copa, no excediendo 50% de la biomasa de los elementos arbóreos nativos, posteriormente el manejo ha sido principalmente cada año realizar la poda de las ramas con crecimiento descendente; lo anterior con la intención de aprovechar sus ramas [principalmente para leña (Alanís-Flores y Velazco-Macías, 2013) y de obtener postes (Mendoza-Ortiz, Lara-Bueno, Uribe-Gómez y García-Nuñez, 2018), así como frutos y follaje (para alimento del ganado) y sombra, debido que las copas del arbolado favorecen la creación de un microclima para el resguardo del ganado en temporadas de calor (Sarmiento et al., 2015). Autores refieren que Prosopis laevigata se encuentra presente en los agostaderos comúnmente acompañado de otras especies arbóreas, manejadas de forma tradicional y con alto valor empírico, en este caso Ebenopsis ebano (Mendoza-Ortiz et al., 2018).
Método de muestreo
En abril de 2014 se establecieron al azar cinco sitios de muestreo cuadrangulares de 50 m × 50 m cada uno (2500 m2), este tamaño de sitio es el mínimo recomendable para masas mixtas de acuerdo con el número de especies, de manera que incluya un mínimo de 30 árboles inventariables dentro del sitio y el tamaño cuadrangular tiene la ventaja de tener una relación perímetro-superficie menor. El número de sitios se determinó mediante el siguiente modelo matemático (Mostacedo y Fredericksen, 2000), tomando en consideración la variable cobertura.
Donde:
n= |
número adecuado de parcelas de muestreo |
E = |
error (20%) |
t = |
valor extraído de las tablas de t de Student (p < 0.05) |
N = |
total de unidades muestrales en toda la población |
CV = |
coeficiente de variación |
Se realizó un inventario de todas las especies arbóreas ≥ 7.5 cm de diámetro a la altura de 1.30 m (DAP). Se obtuvo información dasométrica como: especie, altura (h), diámetro basal (d0.10m), DAP y cobertura de copa (dmenor y dmayor). Cada árbol fue localizado espacialmente mediante la medición de la distancia y azimut de cada individuo desde el centro del sitio.
Para cada especie se calcularon los siguientes parámetros estructurales de la vegetación: abundancia relativa (AR i ) de acuerdo con el número de individuos; dominancia relativa (DR i ) en función a la cobertura de copa; frecuencia relativa (FR i ) basada en su presencia en los sitios de muestreo y el índice de valor de importancia relativo (IVIR) que integra los parámetros anteriores y que adquiere valores porcentuales en una escala del 0 al 100 (Müeller y Ellenberg, 1974; Magurran, 1988).
Para la estimación de la abundancia relativa (AR i ) de la especie i respecto a la abundancia total se empleó la siguiente ecuación:
donde A i es la abundancia absoluta, N i es el número de individuos de la especie i y S la superficie de muestreo en hectáreas.
La dominancia relativa (DR i ) de la especie i respecto a la dominancia total se evaluó mediante:
donde D i es la dominancia absoluta y Ab i el área de copa de la especie i (A=π/4*d 2 ) (Romahn, Ramírez y Treviño, 1994).
La frecuencia relativa (FR i ) de la especie i respecto a la frecuencia total se obtuvo con la siguiente ecuación:
donde F i es la frecuencia absoluta, P i es el número de sitios en los que está presente la especie i y NS es el número total de sitios de muestreo.
El índice de valor de importancia relativo (IVIR) se determinó mediante la fórmula:
El índice de valor forestal (IVF), se aplicó para evaluar la estructura bidimensional de la vegetación arbórea (Corella et al., 2001). Su fórmula es:
Para caracterizar los efectos de la poda y aclareo de los individuos a nivel horizontal en la superficie de una copa, se desarrollaron modelos entre las variables diámetro normal y área de copa, siendo los modelos más comunes el de Prodan, Lineal, Schumacher y Schreuder. A continuación, se detallan las ecuaciones:
Análisis de la información
Para el ajuste de los modelos se utilizó el software R v3.1.2 (R Development Core Team, 2011). Se ajustaron los modelos anteriormente descritos para toda la base general, mediante dos procedimientos. El primero es el procedimiento lm (linear models en R) para el modelo lineal y el procedimiento nls, para los modelos no lineales. Para la comparación entre los modelos se utilizó el coeficiente de determinación R2.
Resultados
Se registraron en total 295 individuos en el área, lo que representa una abundancia 236 ind. ha-1. Se registraron cuatro especies, siendo las más abundantes Prosopis laevigata (Humb. & Bonlp. ex Willd.) M. C. Johnst. (mezquite) y Ebenopsis ebano (Berland.) Barneby & J. W. Grimes (ébano). Las especies Acacia berlandieri Benth (guajillo) y Guaiacum angustifolium Engelm (guayacán) representaron 0.7% de la abundancia total (Tabla 1). Referente a la frecuencia, Prosopis laevigata y Ebenopsis ebano se registraron en 100% de los sitios de muestreo. De acuerdo con la dominancia, el área presentó una dominancia de 10367.8 m2 ha-1, siendo P. laevigata la especie más dominante con 7789.7 m2 ha-1 y G. angustifolium la especie menos dominante con tan solo 4.5 m2 ha-1. La especie ecológicamente más importante de acuerdo con su abundancia, frecuencia y dominancia fue P. laevigata que presentó un IVIR de 56.56%, seguida por E. ebano con 37.57%.
Tabla 1 Valores de abundancia, frecuencia, dominancia e índice valor de importancia relativo (IVIR) de las especies en el área de estudio. Las especies están ordenadas de acuerdo con su valor de IVIR.
| Especie | n | Abundancia | Frecuencia | Dominancia | IVIR | |||
|---|---|---|---|---|---|---|---|---|
| ABS | REL | ABS | REL | ABS | REL | |||
| Prosopis laevigata | ||||||||
| (Humb. & Bonpl. ex Willd.) M. C. Johnst. | 156 | 124.8 | 52.88 | 5 | 41.67 | 7789.7 | 75.13 | 56.56 |
| Ebenopsis ebano (Berland.) Barneby & J. W. Grimes | 137 | 109.6 | 46.44 | 5 | 41.67 | 2551.8 | 24.61 | 37.57 |
| Acacia berlandieri Benth. | 1 | 0.8 | 0.34 | 1 | 8.33 | 21.9 | 8.88 | 2.96 |
| Guaiacum angustifolium Engelm. | 1 | 0.8 | 0.34 | 1 | 8.33 | 4.5 | 8.72 | 2.91 |
| Total | 295 | 236 | 100 | 12 | 100 | 10367.8 | 100 | 100 |
ABS. = absoluta, REL = relativa, IVIR = índice de valor de importancia relativo.
De acuerdo con la distribución de las especies, la distribución de puntos en la Figura 2 muestra tendencia a la distribución aleatoria en los sitios S1 (42 ind.), S2 (75 ind.) y S3 (58 ind.); sin embargo, para el caso de los sitios S4 (69 ind.) y S5 (51 ind.) se observa una tendencia al agrupamiento. Asimismo, los sitios S1, S3 y S5 muestran que existen individuos de mayores diámetros, mientras que en los sitios S2 y S4 las especies muestran diámetros menores.
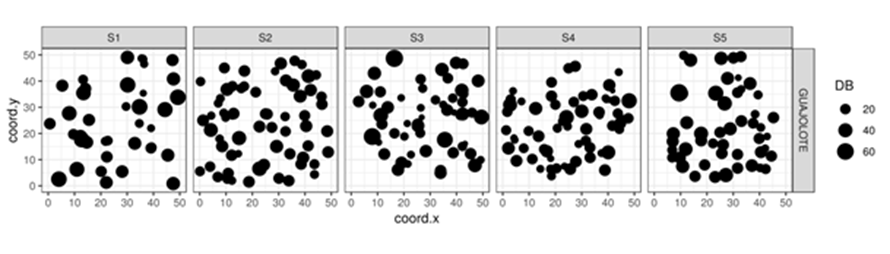
Distribución escalada a diámetro basal, los ejes “x” y “y” indican la distancia en metros.
Figura 2 Mapa de localización espacial de todos los árboles muestreados por sitio.
El índice de valor forestal (IVF), indicó que el diámetro relativo y altura relativa mayores los presentó Prosopis laevigata con 63.06% y 56.49% respectivamente. La cobertura de copa en el área fue de 10266.97 m2 ha-1, de los cuales 2519.70 m2 ha-1 corresponden a Ebenopsis ebano que posee una media de diámetro de copa de 5.21 m2, por su parte los individuos de P. laevigata presentaron una media de 8.49 m2, con una cobertura de copa de 7721.31 m2 ha-1; ambas especies representaron 99.75% de la cobertura de copa del área de estudio y la media para el diámetro de copa de los individuos en el área de estudio fue de 7.17 m2. Los resultados anteriores indican que existe 100% de la superficie cubierta por el dosel y sobreposición de copas.
Los modelos de diámetro normal - área de copa se emplearon para hacer una proyección horizontal de las copas, ya que los efectos de aclareo y crecimiento de copa se encuentran relacionados directamente con el crecimiento en diámetro. Para esto se realizó un modelo general (todas las especies, Tabla 3), siendo el modelo Lineal el que mejor se ajustó al presentar los mayores valores en coeficiente de determinación R2 (0.621), además sus parámetros son estadísticamente significativos (Fig. 2).
Tabla 2 Valores de diámetro, altura y área de copa e índice de valor forestal (IVF) de las especies en el área de estudio.
| Especie | Diámetro | Altura | Cobertura de copa | IVF | |||
|---|---|---|---|---|---|---|---|
| ABS | REL | ABS | REL | ABS | REL | ||
| Prosopis laevigata | |||||||
| (Humb. & Bonpl. ex Willd.) M. C. Johnst. | 2765.04 | 63.06 | 898.88 | 56.49 | 7721.31 | 75.21 | 194.75 |
| Ebenopsis ebano (Berland.) Barneby & J. W. Grimes | 1604.34 | 36.59 | 684.64 | 43.02 | 2519.70 | 24.54 | 104.16 |
| Acacia berlandieri Benth. | 8.96 | 0.20 | 4.72 | 0.30 | 21.63 | 0.21 | 0.71 |
| Guaiacum angustifolium Engelm. | 6.16 | 0.14 | 3.12 | 0.20 | 4.32 | 0.04 | 0.38 |
| Total | 4384.50 | 100 | 1591.36 | 100 | 10266.97 | 100 | 300 |
ABS. = absoluta, REL = relativa, IVF = índice de valor forestal
Tabla 3 Estadísticos y parámetros de los modelos probados
| General | Parámetros | ||||
|---|---|---|---|---|---|
| Función | b0 | prob | b1 | prob | R2 |
| Prodan | 4.56 | < 2e-16 | -7.19 | < 2e-16 | 0.620 |
| Lineal | 2.20 | < 2e-16 | 0.26 | < 2e-16 | 0.621 |
| Schumacher | 13.61 | < 2e-16 | 14.60 | < 2e-16 | 0.618 |
| Schreuder | 4.56 | < 2e-16 | -5.89 | < 2e-16 | 0.620 |
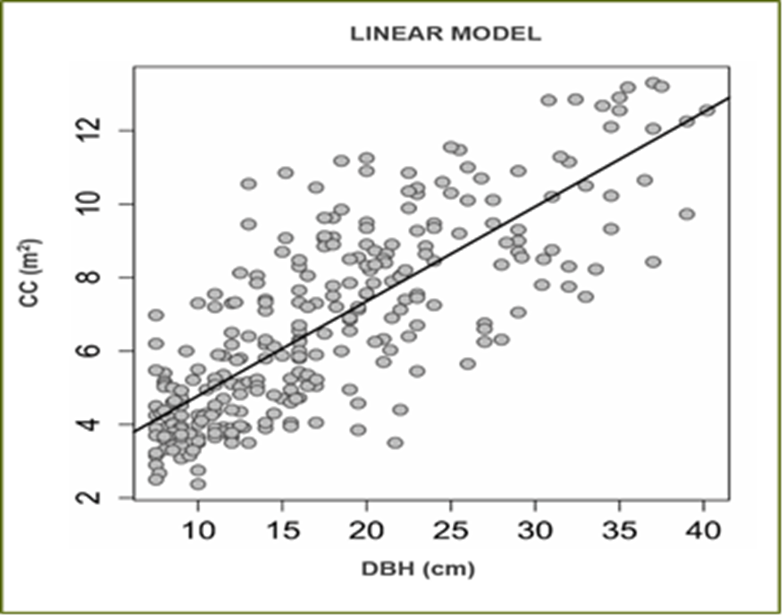
Figura 3 Modelo lineal para diámetro normal (DBH)-área de copa (CC) para todas las especies en el área de estudio.
La estructura vertical del componente arbóreo muestra una distribución cercana a la normal, donde se encuentran pocos individuos en las clases de altura, bajas y altas, y un elevado número en las intermedias (Fig. 3). La altura dominante del arbolado se ubica en la clase de altura de 6 m - 8 m, siendo la altura media para el área de estudio de 6.78 m; la clase de altura dominante estuvo representada por Prosopis laevigata y Ebenopsis ebano, que presentaron una altura media de 7.20 m y 6.25 m, respectivamente. Del arbolado presente, 77.7% se encontró distribuido entre los estratos de 4 m - 6 m y 6 m - 8 m, los cuales estuvieron conformados en 99% por P. laevigata y E. ebano. Las otras dos especies se encontraron representadas solo en los primeros dos estratos (2 m - 4 m y 4 m - 6 m). Esta distribución de las especies dominantes indica que tanto el mezquite como el ébano se encuentran mezclados en los diferentes estratos de altura.

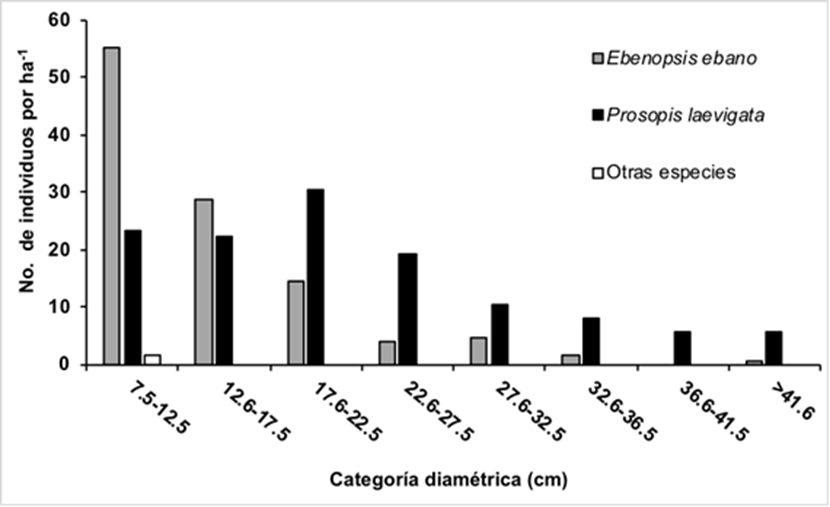
La distribución diamétrica de la comunidad arbórea muestra una línea logarítmica con tendencia negativa en la densidad conforme aumenta el diámetro (Fig. 4). Las densidades mayores de individuos (˃ 40 ind. ha-1), se concentraron en las clases diamétricas < 22.5 cm de diámetro, siendo la media para el DAP de 18.41 cm. La clase menor (7.5 cm - 12.5 cm) se encuentró dominada por la especie E. ebano, que presentó una media para el DAP de 14.63 cm; sin embargo, conforme aumenta la categoría diamétrica el mayor número de individuos por hectárea lo representó P. laevigata, cuyos individuos presentaron una media para el DAP de 22. 15 cm.
Discusión
A pesar de que los sitios de transición entre varios tipos de vegetación son ricos en biodiversidad (Toledo, 1995) en el presente estudio únicamente se registraron cuatro especies (A. berlandieri, G. angustifolium, E. ebano y P. laevigata), el manejo en el área de estudio propició que se eliminaran aquellos individuos que el manejador no consideró importantes para el ganado o bien que por sus características impedían el desplazamiento de este en el área.
Las especies E. ebano y P. laevigata representaron 99.3% de la abundancia de la comunidad. A pesar de que, dichas especies no suelen tener alta representación en la composición florística de las comunidades vegetales de matorral espinoso tamaulipeco en el noreste de México, si se presentan de manera típica en la región y suelen tener una importancia económica significativa debido a sus diferentes usos, sobre todo como fuente de carbón (Estrada, Yen, Delgado y Villarreal, 2004; Alanís et al., 2017), sobre todo en zonas de transición con mezquitales y en zonas con algún tipo de manejo. Estudios como el de Domínguez et al. (2013) registraron a P. laevigata con 10.2% y 1.4% de abundancia para dos municipios en Nuevo León en comunidades vegetales maduras sin actividad productiva en el matorral espinoso tamaulipeco y no registraron a E. ebano como parte de la comunidad, al igual que Briones y Villarreal (2001) que no encontraron presencia de E. ebano en comunidades de matorral mediano y alto de Prosopis-Acacia; a diferencia de Alanís et al. (2017) que si registraron a E. ebano en un mezquital de norte de Nuevo León, pero no como especie abundante. Por otra parte, Montaño, García, Ochoa y Monroy (2006) registraron una densidad similar para dos matorrales semiáridos en el centro de México, los autores mencionan que en el pasado el mezquite era una de las plantas con mayor presencia; sin embargo, a pesar de su importancia ecológica y económica, en la actualidad sus poblaciones han disminuido en los ecosistemas semiáridos, debido a la fuerte perturbación de estas comunidades vegetales.
Mora et al. (2013) registraron baja abundancia de P. laevigata (1.15%) para un matorral espinoso tamaulipeco de referencia sin actividad productiva activa en el centro-sur de Nuevo León; sin embargo, los mismos autores observaron una abundancia mayor (61.18%) para un área de matorral espinoso tamaulipeco con actividad ganadera, abundancia similar a la registrada en este estudio. Es decir que, estas especies se vuelven muy importantes en áreas con manejo silvopastoril, debido a los beneficios que aportan, ya que son muy apreciadas por los productores de ganado doméstico por su elevado valor nutricional. En el área de estudio mediante el manejo se favorecieron las especies P. laevigata y E. ebano, al no ser taladas al inicio del manejo del área, para de esta manera aprovechar su sombra y sus frutos que son empleados como forraje en las épocas que escasea el alimento y la madera para leña (Alanís, Cano y Rovalo, 1996; Estrada, Villarreal y Jurado, 2005).
A pesar de que A. berlandieri suele presentarse como un taxa abundante sobre grandes extensiones, asociada a diversas comunidades de matorral xerófilo (Estrada et al., 2005) y G. angustifolium suele formar parte de algunos tipos de vegetación de esta zona (González, Ramírez, Cantú, Gómez y Uvalle 2010; Mora et al. 2013; Alanís et al., 2017), no fueron especies dominantes en el área de estudio. Lo anterior es resultado del manejo de las especies, ya que se les ha dado manejo a aquellas que se consideraron más convenientes para el uso agroforestal de la zona y por sus distintos usos, se eliminaron al inicio aquellas que no se consideraron importantes para tal uso; en este caso el mezquite y el ébano se han manejado a través de podas anuales para aprovechar principalmente sus ramas para leña; por lo que la comunidad estudiada presentó una distribución de abundancias en la cual unas cuantas especies son dominantes y las restantes raras.
De acuerdo con la frecuencia de las especies, los resultados fueron similares a lo registrado por Foroughbakhch, Alvarado, Carrillo, Hernández y Guzmán (2013) quienes señalaron a P. glandulosa como la especie más frecuente en distintas comunidades de matorral en estado de transición entre matorral submontano y bosques de mezquite en el noreste de México. Por lo tanto, el manejo del área y que esta sea una zona de transición de matorral espinoso tamaulipeco y mezquital favorecieron el alta la presencia de P. laevigata. Asimismo, estudios como el de Osuna y Meza (2003) y Carrillo, Gómez y Arreola (2007) mostraron que, Prosopis laevigata y Ebenopsis ebano se encuentran altamente presentes en los sistemas agroforestales del noreste de México, debido a su importancia económica como recurso maderable (leña, carbón, postería para cercas y material de construcción), registrándose sistemas agroforestales con Prosopis glandulosa Torr. como especie forestal en el norte de Nuevo León (Comisión Nacional Forestal [Conafor], 2013); de igual manera destacan usos como enseres domésticos, mangos para hachas, azadones, figuras decorativas de ébano (Estrada et al. 2005). Además, los beneficios que otorgan a las áreas que habitan, tales como estabilización y mejoramiento del suelo al incrementar el contenido de materia orgánica, así como su importancia forrajera en la alimentación de ganado en pastoreo y fauna silvestre.
La densidad arbórea del presente estudio fue mayor a la registrada por Sarmiento et al. (2015) (173 ind. ha-1), quienes evaluaron una comunidad arbórea de un sistema silvopastoril pero en un matorral submontano del noreste de México; sin embargo, la densidad ha sido menor en relación con lo hallado por Domínguez et al. (2013), quienes registraron un mayor número de individuos y de especies para comunidades sin actividad productiva de matorral espinoso tamaulipeco para diferentes zonas de Nuevo León, también menor a lo registrado por Alanís et al. (2017) para una comunidad de mezquital en el centro de Nuevo León, quienes registraron una densidad de 375 ind. ha-1. Por lo tanto, es evidente que las áreas con algún tipo de manejo presentaron menor densidad, ya que la diferencia fundamental entre un bosque bajo manejo y o sin este es el control de la densidad (Becerra, 1986). El área de estudio, al presentar manejo agroforestal tanto el número de especies como de individuos, ha disminuido. Lo anterior ha sucedido ya que se ha buscado el mantenimiento de los individuos de mejores dimensiones a través de la primera corta que se realizó de los individuos pequeños y de la poda de los elementos arbóreos a través de los años.
La comunidad vegetal presentó un área de copa de 10367.8 m2 ha-1, lo que equivale a una cobertura de 100% y sobreposición de copa. Prosopis laevigata posee la mayor dominancia en el área con 7789.7 m2 ha-1, posteriormente se encuentra E. ebano con 2551.8 m2 ha-1, lo que representa 99.7% de la dominancia total del área. Estudios como el de Montaño et al. (2006) obtuvieron una cobertura mucho menor (438 y 175 m2 ha-1) para dos matorrales semiáridos en el centro de México y Alanís et al. (2017) registraron 6600 m2 ha-1 para una comunidad de mezquital en el centro de Nuevo León, siendo P. glandulosa la especie más dominante en el área junto con A. farnesiana. Por otra parte, Sarmiento et al. (2015) encontraron un valor menor de área de copa con 5215 m2 ha-1 para una comunidad con un sistema silvopastoril de matorral submontano del noreste de México, lo anterior es debido probablemente a que las especies con mayor abundancia no presentaron un área de copa predominante. Estudios indican que se observa una tendencia de crecimiento mayor en el diámetro de copa en individuos podados que en individuos sin podar (Carrillo et al., 2007), dicho comportamiento puede estar reflejado en los individuos del área de estudio al presentar una alta cobertura de copa y una cubierta de 100% de la superficie debido al manejo al que han sido sometidos desde hace cinco años, lo anterior con la intención de aprovechar sus ramas.
Con respecto al IVIR, Montaño et al. (2006) y Mora et al. (2013) señalaron a P. laevigata como la especie con mayor índice de valor de importancia en comunidades semiáridas del centro y norte de México, respectivamente. Por su parte, Foroughbakhch et al. (2013) indicaron que P. glandulosa es la especie con mayor índice de valor de importancia en el estrato arbóreo de diferentes comunidades de matorral en el noreste de México sin registro de actividades productivas, destacando también E. ebano, entre otras especies del género Acacia. Briones y Villarreal (2001) mencionan que, para un ecotono entre provincias florísticas en el norte de Nuevo León, las asociaciones de Prosopis se dan principalmente con el género Acacia, siendo estas especies las ecológicamente más importantes en matorrales altos y medianos espinosos, sin registrar a la especie E. ebano. En el presente estudio, en el área manejada no se observaron altas densidades del género Acacia, ya que para los manejadores no se consideró como especie importante para el ganado.
Los resultados del IVF fueron similares a los del índice de valor de importancia relativo, destacando Prosopis laevigata (64.92%) y Ebenopsis ebano (34.72%). Estudios como el de Domínguez et al. (2013) registraron una cobertura de 672.7 y 185.1 m2 ha-1, respectivamente, para comunidades de matorral espinoso tamaulipeco en el noreste de Nuevo León, mientras que para el presente estudio se registró una cobertura de 10266.97 m2 ha-1. Esta cobertura se considera alta para comunidades de matorral; sin embargo, es posible que se deba a que P. laevigata es la especie predominante en el sitio y los individuos presentes han sido sujetos a manejo por cinco años, lo que ha ocasionado que los individuos que conforman la comunidad hayan tenido mayor crecimiento tanto en cobertura como en diámetro (Tabla 2). Esta condición no se presenta de manera natural en las comunidades de matorral espinoso tamaulipeco para individuos de esta especie ya que no suelen ser altamente predominantes (Montaño et al., 2006).
El modelo lineal indica que conforme el diámetro normal de los individuos aumenta se presentan mayores valores de área de copa de los individuos; Benítez-Naranjo, Rivero-Vega, Vidal-Corona, Rodríguez-Rodríguez y Álvarez-Rivera (2003) reportaron esta tendencia de crecimiento, ya que el desarrollo de la copa está estrechamente unido al crecimiento en diámetro del tronco; debido que el control de la densidad a través del manejo del área ha influido en el crecimiento de los árboles (Benavides-Meza y Fernández-Grandizo, 2012), ya que al realizar podas y abrir espacios para el desplazamiento del ganado y el crecimiento de pastos ha generado que los árboles representen el estrato dominante, es decir que no existe una fuerte competencia por la luz y las copas de los individuos son grandes, por lo tanto los niveles de fotosíntesis son altos, por lo que poseen un mayor crecimiento en diámetro (Benítez et al., 2003).
La altura dominante del arbolado se ubica en la clase de altura de 6 m-8 m, representada por P. laevigata y E. ebano, lo cual se considera como alto para los matorrales del noreste de México (Domínguez et al., 2013; Mora et al., 2014). La altura registrada para la comunidad fue similar la encontrada por Alanís et al. (2017), quienes registraron la altura dominante en la clase de 5 m - 7 m, en un mezquital del centro de Nuevo León y un intervalo de altura de 2.3 m a 9 m. Estos resultados son muy similares a los del presente estudio y de igual manera a los de Briones y Villarreal (2001), para un ecotono de matorral alto espinoso de Prosopis-Acacia en el norte de Nuevo León. Carrillo et al. (2007) mencionan que, cuando se realizan podas en los individuos de mezquite, se registra un mayor crecimiento en altura; es por tal motivo que los árboles en el área de estudio presentan una altura mayor a la registrada dentro de comunidades de matorral sin manejo silvopastoril.
De acuerdo con la estructura diamétrica la comunidad se encuentra conformada por individuos de diámetros (DAP) menores o iguales a 20 cm en 73.3% de su composición, lo que indica que los árboles son jóvenes y presentan una altura favorable para el diámetro que presentan, condición generada debido al manejo. Lo anterior indica que existe un alto número de individuos de clases diamétricas menores, que podrán ocupar el espacio de árboles muertos o aprovechados, condición propiciada por la tala de los individuos maduros para la apertura de espacios para el desplazamiento de ganado, lo que ha propiciado el crecimiento de los árboles tanto en diámetro como altura, lo que trae como beneficio adicional el aprovechamiento a través de las podas y usar el material para leña. Un estudio desarrollado por Mora et al. (2013) donde evalúan el efecto de la ganadería en tres diferentes condiciones de matorral en el noreste de México, muestra que la clase diamétrica predominante en un matorral de referencia es de 4 cm- 8 cm, lo mismo para el caso de ganadería. Sin embargo, para las siguientes clases diamétricas en el caso de ganadería los individuos se encuentran escasamente representados, situación que no se presenta en el área de estudio, ya que las categorías diamétricas menores a 22.5 cm se encuentran bien representadas, lo anterior muestra el resultado obtenido a través del manejo forestal del cual ha sido objeto el área.
Conclusiones
La comunidad vegetal estudiada presenta una alta densidad de individuos comparado con otras comunidades con sistemas silvopastoriles del noreste de México y una alta cobertura de copa propiciada por el desarrollo de los árboles después de la tala de individuos no deseados al inicio del manejo. Esto permite que los individuos tengan mayor espacio y esto se muestra reflejado en el crecimiento de copa y diámetro normal de los individuos. La estructura vertical de la comunidad indica que existen una alta proporción (68.6%) de árboles altos (≥ 6 m), ya que al realizar podas anuales ha ocasionado que los árboles tengan un mayor crecimiento en altura. En cuanto a su estructura horizontal, la comunidad muestra una buena representación de arbolado en la mayoría de las clases diamétricas. Las especies con mayor índice de valor de importancia e índice forestal de acuerdo con los parámetros ecológicos evaluados fueron Prosopis laevigata y Ebenopsis ebano, ambas presentaron altos valores de cobertura aérea, diámetro y altura. Tanto en la composición horizontal como vertical, los análisis indican que ambas especies se encuentran en asociación vegetal tanto en la estratificación de altura como en las categorías diamétricas. Los resultados muestran que el sistema silvopastoril en la zona ha modificado la estructura de la comunidad vegetal con respecto a comunidades de referencia sin actividades silvopastoriles; no obstante, conserva y promueve determinados atributos ecológicos de la comunidad vegetal nativa.

