











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Paisaje de referencia, un nuevo concepto
La degradación de ecosistemas o el nivel de conservación en un territorio puede ser analizada desde la perspectiva de una unidad de bosque, como del conjunto de ecosistemas que componen un paisaje natural, según la escala del estudio (Schulte et al., 2006; Meli et al., 2017). Los estudios que pueden revisarse abundan en descripciones cualitativas de la degradación, como señalar la pérdida de masa de los bosques, la simplificación de las estructuras, o cuantitativas como la reducción de superficie cubierta de bosques, o niveles de fragmentación de los ecosistemas por mencionar algunos (González-Espinosa, Meave, Ramírez-Marcial, Toledo-Aceves e Ibarra-Manríquez, 2012; Castro-Navarro, Sahagún-Sánchez y Reyes-Hernández, 2017). Respecto a los análisis a escala de paisaje, las cuantificaciones descritas podrían perfeccionarse si para los casos específicos se supieran los intervalos posibles que las variables usadas pueden alcanzar en dichos casos. Castro-Navarro et al (2017) analizaron la relación entre la dinámica de fragmentación y su impacto sobre la distribución de aves en un caso en San Luis de Potosí, México, dividen un área de 770 km2 en cinco niveles de fragmentación, siendo el mínimo nivel 0% a 20% y el máximo 80% a 100%. Un supuesto implícito en este trabajo es que zonas con alta degradación en el momento del estudio, en el pasado fueron un solo fragmento (que pertenecía al intervalo de 0% a 20%), lo que podría ponerse en duda considerando que naturalmente el paisaje podría haber estado fragmentado en cierto grado de forma natural. Gómez Mora, Anaya y Álvarez (2005) en otro estudio para la cordillera de los Andes centrales de Colombia, estiman el estado de conservación de una zona de 257 000 ha apoyándose en métrica del paisaje principalmente fragmentación, y determinan cuantitativamente categorías de conservación, sin considerar si la situación sin degradación hubiera tenido grados de fragmentación. Esta estrategia de análisis se repite en muchos estudios similares (Mas Caussel, 2001; Echeverry y Rodríguez, 2006; Meli et al., 2017). Côrrea do Carmo, Finegan y Harvey (2001) en cambio, en su trabajo de propuesta de conservación analizando la fragmentación de la zona de estudio, propone escenarios futuros de mejoras derivadas de la restauración, y las expresa en mejoras de los mismos índices usados para evaluar la fragmentación. Lo que puede entenderse como una cuantificación de la degradación del ecosistema a escala de paisaje, o bien puede cuantificarse la reducción de la degradación si se logran los objetivos de la restauración. No obstante, los escenarios post restauración se establecieron con metas bibliográficas generales y no propias de la zona estudiada. Varios autores han publicado revisiones que proponen y analizan métodos de determinación de grados de degradación, describen la interpretación de los intervalos teóricos para diversos índices, pero no abordan la modificación que cada uno de ellos puede sufrir según el área que se estudia (Lindenmayer, 1995; Gehlhausen, Schwartz y Augspurger, 2000; Atauri y de Lucio, 2001; Del Río, Montes y Cañellas, 2003; Schulte et al., 2006; Aragón, Abuja, Belinchón y Martínez, 2015). Badii y Landeros (2007) proponen que los índices y los intervalos que adoptan deben ser ajustados a cada caso específico, pero no detallan métodos de cómo hacerlo. En la temática de restauración se ha propuesto el concepto de bosque de referencia o ecosistema de referencia, que se define como el tipo de ecosistema que posee las mejores características de conservación y que se utiliza para comparar los bosques en análisis respecto al ideal deseado. En la misma lógica, se plantea en esta discusión que el área o paisaje que se analiza se podría comparar con un “paisaje de referencia” (PR), definida como la condición sin degradación del área que se estudia, propuesta de forma teórica. De lograr establecerse un PR, podrían aplicarse sobre ella los mismos índices del paisaje que se aplicaron sobre la condición inicial, de manera que la diferencia entre los resultados de los índices sobre la condición inicial y sobre el PR daría una medida cuantitativa del estado de conservación del paisaje.
Vegetación y dinámica de los bosques
Los bosques que habitan los Andes Chilenos en su zona austral constituyen un mosaico de estados sucesionales que no deberían confundirse con degradación antrópica (Lindenmayer et al., 2006; Donoso, 2015). La colonización inicial de las zonas perturbadas coincide con las descritas por Donoso (2015), correspondiente a arbustos pioneros de rápido crecimiento y carácter temporal. A su vez, aunque de forma menos inmediata, aparecen individuos emergentes de especies arbóreas pioneras intolerantes, entre los que destacan Nothofagus nítida (Phil.) Krasser y Nothofagus betuloides (Mirb.) Oerst. En las zonas más elevadas con humedad suficiente y alta disponibilidad de semillas también se establece Fitzroya cupressoides (Molina) I.M.Johnst. Estas comunidades dominarán el dosel formando rodales de alta densidad, coetáneos y frecuentemente puros (Armesto, León y Arroyo, 1995). Décadas después, la menor exposición al viento y a la luz junto con el mayor grado de humedad proporcionado por las especies pioneras, favorece el establecimiento de especies tolerantes como Laurelia philippiana (Looser) y Saxegothaea conspicua Lindl, que se desarrollarán junto a las pioneras hasta alcanzar una condición de bosque mixto multietáneo de tolerantes e intolerantes, que Donoso (1981) clasificó como subtipo de Siempreverde con Intolerantes emergentes, y que correspondería a una sucesión secundaria del tipo. Finalmente, este bosque mixto evolucionará aproximándose a etapas de clímax con la desaparición de las especies pioneras conformando el subtipo siempreverde de tolerantes (Veblen, Schlegel y Escobar, 1980; Donoso, 1981).
Índices usados para construir la métrica del paisaje
Los índices del paisaje son herramientas que permiten establecer una métrica del estado de conservación de un área específica, cuya construcción se basa en información espacial (Côrrea do Carmo et al., 2001; Del Río et al., 2003; Lindenmayer, Franklin y Fischer, 2006).
Un índice espacial es el efecto borde que fue planteado por Leopold (1933) haciendo referencia a la alta riqueza de especies encontrada en una transición entre hábitats adyacentes. Puede definirse como el resultado de la interacción entre dos ecosistemas adyacentes (Murcia, 1995)y actualmente incluye todos aquellos procesos, influencias y flujos ecológicos capaces de originar cambios en la estructura y composición de hábitats contiguos (López-Barrera, 2004).
Otro índice basado en información espacial es la diversidad del paisaje, que mide tanto la riqueza como la uniformidad de los elementos que conforman un conjunto y se utiliza como medida de biodiversidad para una zona (McGarigal, 2002). La diversidad de especies está determinada, entre otros factores, por la heterogeneidad del paisaje (Atauri y de Lucio, 2001) que puede ser calculada a través del índice de Shannon (H) (Shannon y Weaver, 1962) (Ecuación 1).
Adaptando el concepto a términos espaciales se puede calcular la diversidad espacial, reemplazando especie por clase de ecosistema en el área de estudio, siendo finito con un total de n clases y p i la proporción de área ocupada por la clase i (Nogués Bravo y Pérez Cabello, 2001; Rebolledo y Rau, 2010). La diversidad espacial por defecto define su mínimo en cero cuando hubiera un solo elemento en el área y contuviera toda la superficie analizada. Respecto al máximo, no obstante, se requiere un supuesto en el que tanto del número de clases como la superficie de ellas alcancen las condiciones teóricas perfectas. Los trabajos revisados no presentan experiencias al respecto.
El índice de forma, otro índice del paisaje ofrece una medida de vulnerabilidad de un ecosistema frente a otro exterior sobre la base de la forma de sus límites, calculada con su relación área-perímetro. Así, la forma más compacta y menos vulnerable para un ecosistema es la circular ya que contiene la máxima superficie en el mínimo perímetro, aumentando el núcleo interior (compactibilidad) y minimizando el borde colindante con la matriz (vulnerabilidad) (Rau Acuña, 2010; Lozano Botache, Gómez Aguilar y Valderrama Chaves, 2011). Patton (1975) propuso un índice de diversidad de borde o índice de forma (IF) que relaciona la similitud entre la forma de una superficie y la de una circunferencia con la misma superficie (Ecuación Nº2) (Mas Caussel, 2001; Lozano Botache et al., 2011; Didham y Ewers, 2012).
Donde P es el perímetro y A la superficie del ecosistema, comúnmente llamado parche; IF alcanza su mínimo (óptimo) en 1, para una forma circular, y va aumentando conforme la superficie difiere del círculo. La continuidad espacial como índice de paisaje (ICE) mide el grado de fragmentación de un bosque. Conceptualmente, la fragmentación impide el intercambio genético y el tránsito entre unidades de bosque, amenazando los nichos y la supervivencia de las especies (Lozano Botache et al., 2011). Vogelmann (1995) calculó la continuidad forestal a través de la relación área-perímetro del conjunto de fragmentos que componen una clase de hábitat forestal Un alto valor de ICE (Ecuación 2) supondrá una alta continuidad espacial en la totalidad del hábitat, siendo menor su grado de fragmentación (Vogelmann, 1995; Echeverry y Rodríguez, 2006; Lozano Botache et al., 2011).
Ai es el área (m2) y Pi el perímetro (m) de todos los fragmentos del hábitat correspondiente a la clase i.
Objetivos
Analizar el potencial de índices del paisaje para determinar cuantitativamente el estado de conservación de casos específicos, mediante la descripción de su desempeño en una zona de alta conservación.
La hipótesis de trabajo fue que si era posible analizar cuál debiera ser el estado de conservación ideal de un área específica, llamada en este estudio “paisaje de referencia”, entonces sería posible cuantificar el grado de degradación de dicha área, como la diferencia entre los resultados de los índices en la condición del PR teórico y los resultados de los índices en la condición actual. Una segunda hipótesis fue que los índices del paisaje usados permiten comprender y cuantificar la variación del estado de conservación del paisaje.
Materiales y métodos
Para cumplir los objetivos, en síntesis, se elaboró la cartografía actual del área objetivo detallando los tipos de bosques y grados de alteración de ellos sobre la base del conocimiento de la dinámica de los bosques de la región. Luego se creó una segunda cartografía, sobre la base de la primera, donde se supuso que las áreas alteradas habrían mutado a bosque por la acción de restauración. Finalmente, para cuantificar las diferencias entre la condición original y el PR, es decir el segundo mapa modificado por las acciones de restauración, se analizó la diferencia que los índices del paisaje generaron entre la condición inicial y la final.
El área de estudio fue el parque privado Tagua Tagua en la provincia de Llanquihue en la Región de Los Lagos, Chile, (41°43’15’’ S 72° 09’ 32’’ O) a una altura entre 100 m y 1600 m snm aproximadamente (Fig. 1).
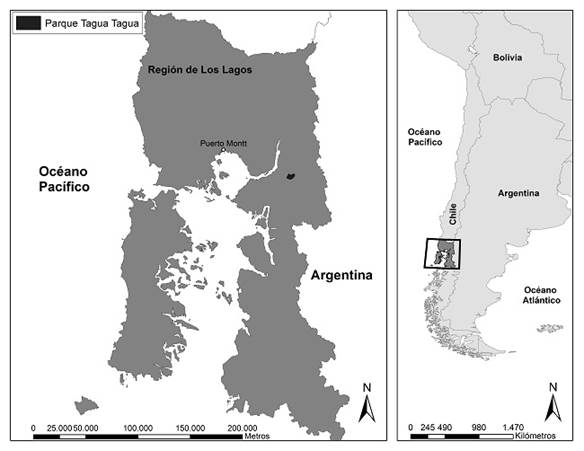
La fisiografía abarca un amplio intervalo de pendientes, desde 5% en las zonas internas del valle hasta 60% en las zonas abruptas. De acuerdo con la caracterización climática de Köepen, pertenece al Clima Templado Lluvioso con influencia Mediterránea (Cfbs) (Hendrischs Pérez, 1948). Presenta temperaturas moderadas (media anual de 10 °C variación entre 6 °C y 14 °C) (Merkel, A, 2018). Las precipitaciones son de 1802 mm anuales. Corresponde a una cuenca forestal altamente conservada, aunque existe un área alterada por actividades humanas anteriores en la zona baja del valle (Fig. 2).
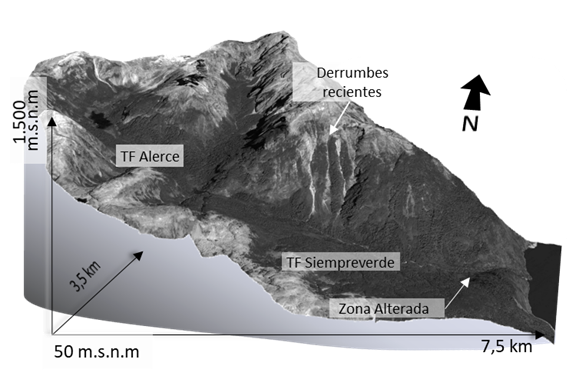
Figura 2. Esquema en visión tridimensional de la ubicación del área de estudio. Se observa la distribución de bosques siempreverdes en las zonas más bajas y el bosque del TF Alerce en las zonas altas.
La zona contiene principalmente el Tipo Forestal (TF) Siempreverde y sobre los 700 m snm el TF Alerce, ambos determinados en la Tipología forestal de Chile (Donoso, 2015).
El conjunto forestal del parque, desde un punto de vista de la dinámica del bosque, está compuesto principalmente por tres etapas sucesionales de un mismo proceso: una de área con bosque recién destruido por fenómenos naturales, principalmente derrumbes por inestabilidad de las laderas susceptibles a los movimientos telúricos, otra de formación forestal temprana y otra más avanzada; coincidiendo con la dinámica del tipo forestal siempreverde descrita por diversos autores (Veblen et al., 1980; Donoso, 1981; Armesto et al., 1995; Rebolledo y Rau, 2010; Donoso, 2015). La primera etapa, o etapa sucesional inicial, está compuesta por especies pioneras, que en la parte alta se asocian con Alerce. Un caso particular de sucesión primaria es una primera colonización de F. cupressoides, que luego de décadas permite el establecimiento de N. nitida, la que puede superar a F. cupressoides dado el lento crecimiento de F. cupressoides. La etapa más avanzada o etapa sucesional intermedia se caracteriza por una mezcla de especies pioneras adultas y en envejecimiento, mezcladas con las especies tolerantes adultas, pero con diferencias de hasta 100 años con las anteriores (análisis del autor no publicados).
Por encima de 1100 m snm y coincidiendo con el límite altitudinal de la vegetación arbórea aparece el subtipo bosques achaparrados o krummholz de Lenga que, según la clasificación de Donoso (1981), se caracteriza por la fisionomía de matorral que adopta la especie arbórea Nothofagus pumilio, debido a la severidad del clima y el sitio.
El área baja por donde existe el acceso y cercana al lago, posee áreas de pradera y matorral con especies nativas y exóticas a consecuencia de asentamientos humanos abandonados (Fig. 2).
Método
Caracterización de la composición del paisaje dentro de la cuenca
La determinación de los componentes presentes en el parque se realizó por fotointerpretación de una imagen aérea hiperespectral de 160 bandas, en un intervalo espectral de 400 nm a 990 nm con una resolución radiométrica de 16 bits y espacial de 30 cm × 30 cm. La imagen fue obtenida con el sensor Hyspex VNIR-1600 (Norsk Elektro Optikk). Se crearon claves de identificación de los diferentes tipos de bosque, y otras coberturas a partir de visitas a terreno durante septiembre de 2012. Con el programa ENVI 4.1 (Research System Inc. [RSI], 2005) se probaron diferentes combinaciones de bandas en la imagen, buscando el mayor contraste para cada una de las categorías de uso del suelo en el parque.
En la confección de la cartografía, se entendió el uso como la clase de ocupación principal en un determinado suelo, y asociación para el caso de Uso Bosque, como la predominancia de una o dos especies (sucesiones primarias), o bien cuando el bosque tiene más de dos especies y todas en una relación importante en la composición (mixtas). La confección de la cartografía se realizó en ArcGIS 10.1 (Environmental Systems Research Institute [ESRI], 2012) utilizando la herramienta “Create Ploygon Features”. Posteriormente se unificaron aquellos polígonos con el mismo tipo de cobertura mediante la opción “Dissolve” incluida en “Generalization toolset” también disponible en ArcGIS (ESRI, 2012).
Propuesta de paisaje de referencia
En el grupo de Usos definidos, se identificaron aquellos provocados por acción antrópica y se supuso que sería posible restaurarlos a una condición de bosque. Bajo este supuesto, los usos originales se cambiaron a bosques en sucesión primaria, asignándoles la asociación primaria probable para la zona donde se encontraban. Así, el PR, respecto al inicial, aumentaría la superficie de bosque y perdería los tipos de usos de origen antrópico.
Análisis del efecto borde
Para analizar las distintas opciones en la aplicación de un índice de efecto borde en un área de alta conservación, se consideraron tres escenarios:
Escenario 1: Ignorando diferentes tipos de bosque de manera que se consideraron dos hábitats adyacentes, uno compuesto por la totalidad del bosque y otro constituido por el resto de los usos fuera del hábitat forestal. La profundidad de la influencia de borde usada en este caso fue 60 m hacia el interior de bosque, considerando las recomendaciones ya descritas por diversos autores. Este escenario se consideró como la aplicación por defecto ya que no incorpora las particularidades intrínsecas específicas del área.
Escenario 2: Este escenario incorporó un análisis de las dinámicas específicas del área de estudio, determinando diferentes posibilidades de vecindad (distancia de influencia entre ecosistemas). Así, se propuso que bosques pioneros (etapa sucesional inicial) por su propia dinámica, no reciben influencia de áreas de “no-bosque”, dado que se originan en estas áreas y las influencias negativas de la matriz descritas en forma teórica no existirían. Así también, las etapas sucesionales intermedias reciben influencia de áreas vegetales de “no-bosque”, siendo menos significativa si es matorral. La influencia más significativa provendrá de praderas originadas antrópicamente (Tabla 1).
Tabla 1. Distancias de influencias de la matriz sobre el bosque, para la aplicación del efecto borde, definidas según escenarios para definir áreas de influencias.
| Escenarios | Clase de uso | Afloramiento Rocoso | Laguna | Mallín | Río | Matorral Antrópico | Matorral Natural | Pradera Antrópica |
|---|---|---|---|---|---|---|---|---|
| Escenario 1 | Bosque | 60 | 60 | 60 | 60 | 60 | 60 | 60 |
| Escenario 2 y 3 | Bosque | |||||||
| Etapa sucesional inicial | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Etapa sucesional intermedia | 0 | 0 | 0 | 0 | 30 | 30 | 60 |
Escenario 3: Proyección futura bajo el supuesto que el área se recuperará de la degradación y que constituye el PR, en que el matorral antrópico y la pradera antrópica, se suponen recuperadas y pertenecen a un bosque en su etapa sucesional final. La magnitud de la influencia para este escenario (longitud que alcanza el efecto) fue idéntica a la del escenario 2.
Análisis de la diversidad espacial
Se determinó el valor de la diversidad espacial inicial (Ecuación 1) usando las clases de uso determinadas por la fotointerpretación referidas a bosques, y la superficie que cada una de ellas tuvo. No se consideraron aquellos usos no vegetales que no tendrían dinámicas de cambio, como lagunas, rocas y otros. Luego se determinó la diversidad espacial en el PR considerando que las áreas alteradas antrópicamente son recuperadas, transformándose en bosques primarios. Finalmente, para discutir las cotas del parámetro teórico se determinó el valor de diversidad máximo para una condición en que cada uso tiene la misma proporción de superficie en el área.
Análisis de la forma y de la continuidad espacial
Para la determinación del índice de forma (Ecuación 2) se construyó una nueva cobertura vectorial uniendo todos aquellos polígonos de cualquier tipo de bosque que estuvieran adyacentes, independiente de su estado sucesional, sobre el supuesto que las influencias adversas entre estados sucesionales serían mínimas o inexistentes. Dado que con esta operación quedarían áreas de bosques separadas por dos probables situaciones:
La separación entre bosques con usos naturales del área no provocados por acción antrópica, por tanto, ambos fragmentos nunca podrían unirse.
La separación se pudo deber a zonas alteradas por acción antrópica y eventualmente con el cumplimiento de la restauración se unirían áreas separadas, en cuyo caso, el fragmento ganaría el área que inicialmente no era bosque y se uniría con la restauración en el PR.
Definida la mencionada capa de fragmentos de bosque inicial, se determinó para cada polígono el IF, para luego determinar un IF promedio ponderado por la superficie de cada fragmento para toda el área, fijándose así el índice de forma inicial para el área.
Para establecer el IF del área de referencia, bajo el supuesto fijado en b), se creó una nueva cobertura vectorial, redefiniendo las áreas alteradas (matorrales antrópicos y praderas antrópicas) como bosques. La determinación del IF para este caso también fue el IF promedio ponderado por su superficie equivalente.
Resultados
Caracterización de la composición del paisaje dentro de la cuenca y formación del paisaje de referencia
La fotointerpretación y definición de usos en el parque arrojó 11 tipos de uso (Tabla 2), cuatro de los cuales corresponden a bosques y sus correspondientes etapas sucesionales. En el Uso de Bosques Primarios se definieron tres tipos de asociaciones, dos de ellas definidas por la presencia minoritaria y mayoritaria de F. cupressoides y otra, fuera del área de distribución de F. cupressoides, formada por bosques puros de N. nítida. El Uso de Bosque mixto corresponde a bosques en estados sucesionales avanzados. Esta configuración constituyó la base para el análisis de los índices.
Tabla 2. Distribución de los componentes del paisaje en el interior del Parque Tagua Tagua, en la condición inicial en comparación con el paisaje de referencia posible.
| Superficies Paisaje inicial | Superficies Paisaje de referencia | ||||||
|---|---|---|---|---|---|---|---|
| Tipo Uso | Asociación | hectáreas | % bosque | % paisaje total | hectáreas | % bosque | % paisaje total |
| 1.Bosque mixto | Bosque Mixto | 685 | 50 | 24 | 685 | 47 | 24 |
| 2.Bosque primario | N.d. ó N.n | 374 | 27 | 13 | 459 | 32 | 16 |
| 3.Bosque primario | N.d. - F.c. | 275 | 20 | 10 | 275 | 19 | 10 |
| 4.Bosque primario | F.c. - N.d. | 39 | 3 | 1 | 39 | 3 | 1 |
| Total bosque | - | 1373 | 100 | 48 | 1458 | 100 | 51 |
| 5.Matorral antrópico | - | 83 | 2,9 | - | |||
| 6.Pradera antrópica | - | 2 | 0,1 | - | |||
| 7.Matorral Natural | - | 456 | 16,1 | 456 | 16,1 | ||
| 8.Áreas de rocas | - | 872 | 30,7 | 872 | 30,7 | ||
| 9.Mallines | - | 32 | 1,1 | 32 | 1,1 | ||
| 10.Lagunas | - | 18 | 0,6 | 18 | 0,6 | ||
| 11.Ríos | - | 4 | 0,1 | 4 | 0,1 | ||
| TOTAL | - | 2840 | 100 | 2840 | 100 | ||
Dónde: N.d.=Nothofagus dombeyi; N.n. = Nothofagus nítida; F.c. = Fitzroya cupressoides
Valores subrayados representan los usos que pueden cambiar con una potencial restauración y que aumentan el bosque total a 51%.
Los tipos de uso posibles de restaurar a bosque fueron los matorrales antrópicos y praderas antrópicas ubicadas en la parte baja de la cuenca (Fig. 2), cuya superficie llega a 3% del total del paisaje (Tabla 2). Así, los efectos de una eventual restauración del área, significaría perder dos categorías de uso, bajando a nueve en total, y un aumento de la superficie boscosa en torno a 3%, esta última cifra reflejó el estado prístino de la zona. Nótese que las áreas sucesionales primarias están en torno a 50% del total del bosque, sin mediar acción humana. Por tanto, puede asumirse que bosques similares en el área, están afectados a tal grado por la acción de la destrucción natural, que el paso de áreas sucesionales primarias a secundarias, aumentarán la superficie de las últimas, pero en la misma medida otras secundarias se estarán destruyendo. Entonces, mientras no se conozcan para este paisaje específico dicha dinámica de cambio natural, no pueden incorporarse supuestos para utilizar en la creación de un PR. De esta forma, dicha dinámica de cambio no fue considerada. De la misma forma, áreas no bosque no originadas por acción antrópica y que ocupan cerca de 49%, pueden considerarse permanentes, y de esa forma se asumieron en el PR.
Análisis del efecto borde
El efecto de la matriz sobre el bosque considerando solamente dos tipos de hábitats (escenario 1), afecta a 30% de la superficie de bosque (Tabla 3), es decir existen 408 ha de bosque periférico que está dentro de una franja de 60 m y son aledañas a algún polígono no bosque (Fig. 3, superior). No obstante, los criterios del escenario 2, que consideró influencias diferenciadas entre hábitat, redujeron el área de influenciada por la matriz a 4%. El escenario 3, que supuso el PR con los criterios del escenario 2, reduciría dicha influencia a 2%, que correspondería a un mínimo natural generado por los matorrales naturales únicamente.
Tabla 3. Distribución de la superficie de bosque analizando el efecto borde para cada escenario.
| Bosque | Superficie (ha) | % Superficie | |
|---|---|---|---|
|
Escenario
1 |
Con influencia de la matriz | 408 | 29,71 |
| Sin influencia de la matriz | 965 | 70,29 | |
| Total | 1373 | 100 | |
|
Escenario
2 |
Con influencia M. Antrópico (30 m) | 26 | 1,89 |
| Con influencia M. Natural (30 m) | 28 | 2,03 | |
| Con influencia Pradera (60 m) | 1 | 0,04 | |
| Sin influencia de la matriz | 1318 | 96,04 | |
| Total | 1373 | 100 | |
|
Escenario
3 |
Con influencia de la matriz | 28 | 1,91 |
| Sin influencia de la matriz | 1430 | 98,09 | |
| Total | 1458 | 100 |
El escenario 1 se considera por defecto; el escenario 2 hace diferencias entre las comunidades que tienen vecindad; el escenario 3 representa el paisaje de referencia.
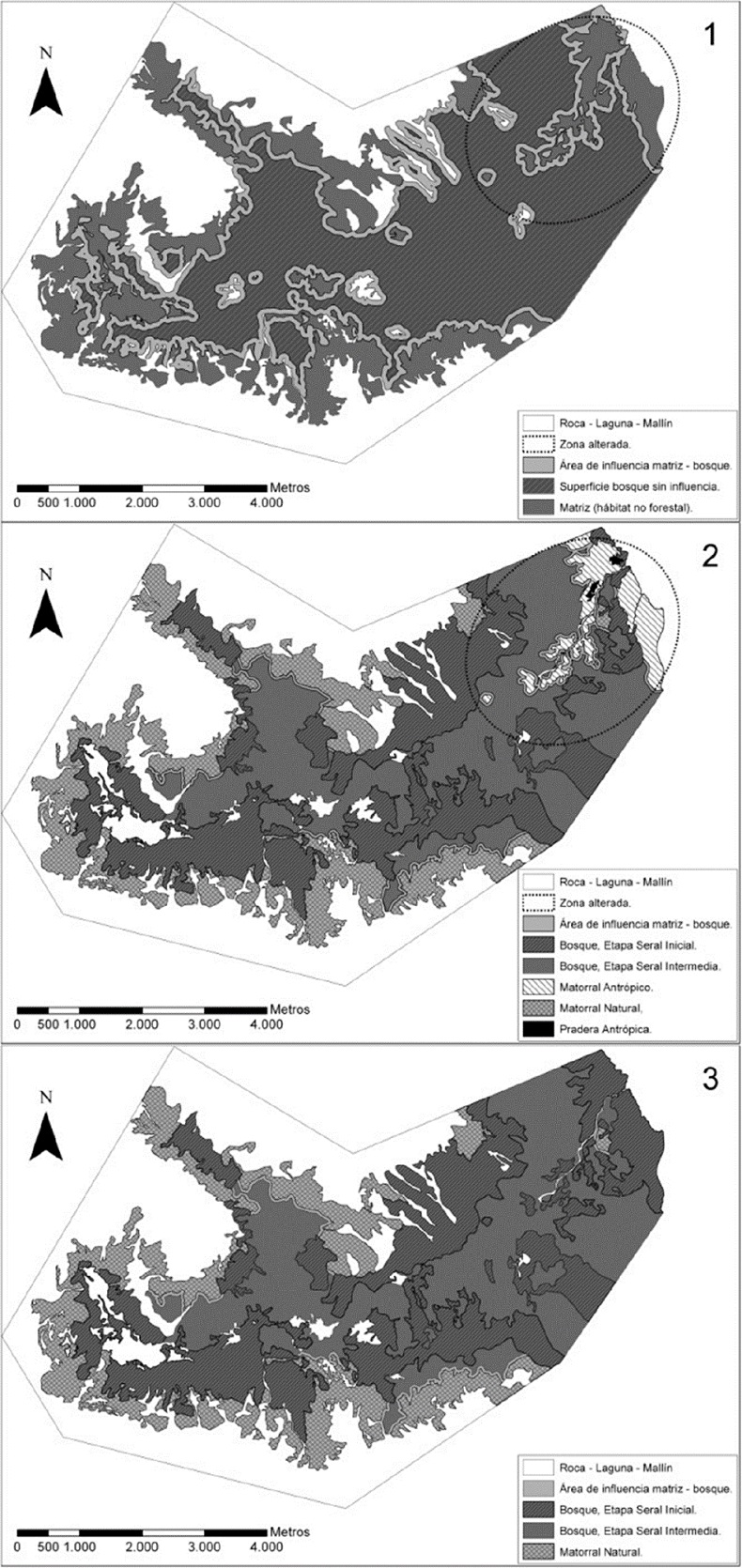
En la figura 3, el escenario 1 (superior), por defecto, solamente tiene dos áreas: bosque y no bosque (matriz) y su correspondiente área de influencia de 60 m hacia el interior del bosque. La zona marcada con línea punteada indica la ubicación de las praderas y matorrales antrópicos. El escenario 2 muestra cómo ya no se consideran áreas de influencia en unos casos y en otros se reducen a la mitad (30 m). En el escenario 3 (inferior) desaparecen las áreas de influencia de praderas y matorrales antrópicos, que en el paisaje de referencia serían bosque
La incorporación del análisis de la propia dinámica del bosque en la determinación del efecto borde provoca una drástica diferencia y que puede visualizarse claramente en la sección oeste de la figura 3 (escenarios 1 y 2), y que corresponde a la zona sin intervención de la parte alta de la cuenca (Fig. 2).
Análisis de la diversidad espacial
Las seis clases de vegetación interesantes de analizar según diversidad fueron las de bosque y de alteración antrópica. Estas clases y sus razones respecto a la superficie total resultaron en un índice de diversidad mayor a aquel del PR en el cual desaparecerían las áreas alteradas antrópicamente (de 1,29 a 1,13 en una reducción de 12% respecto al original), vale decir, la diversidad se reduce con la restauración (Tabla 4; Fig. 4). Este resultado fue evidente ya que la desaparición de dos clases por la restauración reduce la diversidad del paisaje. Por otra parte, el límite teórico de diversidad para el PR, que sería conseguir que las cuatro clases finales tuvieran la misma proporción, mejoraría en 0,16 la diversidad. No obstante, no es conocida la razón teórica que cada categoría debiera tener en condiciones de dinámica sin intervención, por tanto, el límite teórico para el índice de diversidad únicamente serviría como referencia.
Tabla 4. Variación del índice de diversidad H, según la condición actual, la condición en el paisaje de referencia, y los máximos valores teóricos que H puede alcanzar para las categorías de uso de bosques encontradas en el parque.
| Tipo Uso | Asociación | Superficie inicial (i) | Paisaje de referencia (Pr) | Paisaje de referencia Teórico (Prt) | pi | ppr | pprt |
|---|---|---|---|---|---|---|---|
| 1. Bosque mixto | F. cupressoides - N. dombeyi | 39 | 39 | 364,5 | 0,03 | 0,03 | 0,25 |
| 2. Bosque primario | N. dombeyi o N. nitida | 374 | 459 | 364,5 | 0,26 | 0,32 | 0,25 |
| 3. Bosque primario | N. dombeyi - F. cupressoides | 275 | 275 | 364,5 | 0,19 | 0,19 | 0,25 |
| 4. Bosque primario | Bosque Mixto | 685 | 684 | 364,5 | 0,47 | 0,47 | 0,25 |
| 5. Matorral antrópico | - | 83 | 0,06 | ||||
| 6. Pradera antrópica | - | 2 | 0,001 | ||||
| TOTAL | 1458 | 1458 | 1458 | 1 | 1 | 1 | |
| H | 1,29 | 1,13 | 1,39 |
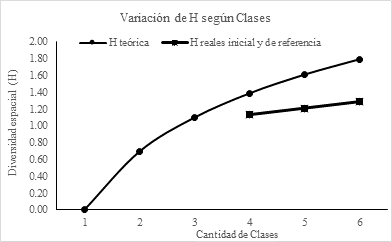
Figura 4. Variación teórica de la diversidad espacial (H) y los niveles reales actuales y del paisaje de referencia.
Nótese que, si se acepta el supuesto de alto grado de conservación del parque, sería posible afirmar que áreas no alteradas de bosques de este tipo en los Andes tienen una dinámica de alteración tal, que más de la mitad de la superficie (53% de etapas sucesionales iniciales) corresponde a bosques pioneros y el resto estaría en procesos más avanzados de la sucesión.
Análisis de la forma y de la continuidad espacial
La nueva capa vectorial para el cálculo de IF, generó ocho polígonos, de los cuales un polígono agrupa casi 95% de la superficie total de bosque, debido a la condición prístina del área (Tabla 5), y que tuvo una fuerte influencia en el IF del área original.
Tabla 5. Polígonos definidos para la determinación del índice de forma (IF), para la condición inicial como para el paisaje de referencia.
| uso | Superficie (ha) | Perímetro (m) | IF | % |
|---|---|---|---|---|
| Condición inicial | ||||
| Bosque | 1 292,1 | 75 957 | 5,96 | 94,1% |
| Bosque | 36,8 | 4 779 | 2,22 | 2,7% |
| Bosque | 27,0 | 6 538 | 3,55 | 2,0% |
| Bosque | 11,1 | 1 877 | 1,59 | 0,8% |
| Bosque | 2,6 | 711 | 1,23 | 0,2% |
| Bosque | 1,7 | 977 | 2,12 | 0,1% |
| Bosque | 0,7 | 340 | 1,12 | 0,1% |
| Bosque | 0,6 | 343 | 1,26 | 0,0% |
| Área alterada | 85,5 | 6,2% | ||
| Condición final | ||||
| Bosque | 1 380,6 | 69 078,7 | 5,2 | 95% |
| Bosque | 36,8 | 4 779,1 | 2,2 | 3% |
| Bosque | 27,0 | 6 538,5 | 3,5 | 2% |
| Bosque | 11,1 | 1 876,7 | 1,6 | 1% |
| Bosque | 2,6 | 710,6 | 1,2 | 0% |
En la categoría Área alterada se agruparon los polígonos de praderas y matorrales de origen antrópico.
Reafirma esta condición de alta conservación el hecho de que las áreas posibles de recuperar con restauración solamente comprenden 6 % del área actual de bosque. La construcción del PR (Fig. 5b) permitió determinar que el bosque no puede constituir una sola unidad, dado que tiene fragmentos separados de manera natural. Así también, la teórica restauración de las áreas alteradas provoca que aumente el polígono mayor, además de unir un polígono que en la condición inicial resultaba separado, lo que reduce de ocho a cinco los polígonos potenciales en el PR.
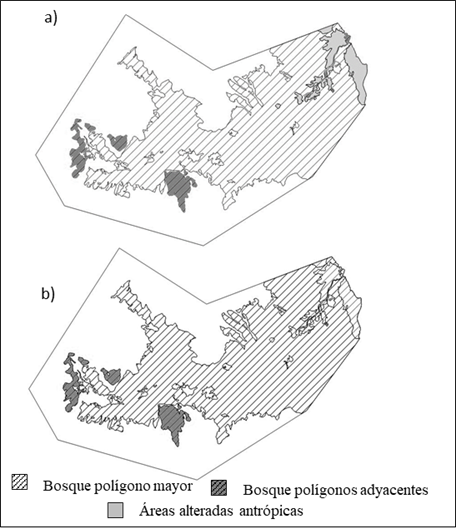
Figura 5. Coberturas vectoriales creadas para la determinación de IF. a) Condición inicial mostrando polígonos de áreas alteradas. b) Polígono del paisaje de referencia, sin áreas alteradas, que se muestran con contorno, pero sin color en la sección derecha.
La geografía de la cuenca determina que la mejor forma que pudo ofrecer el PR para potenciales bosques asemeja a una media luna horizontal muy irregular y, si bien la configuración teórica de IF indica que un resultado ideal debiera ser en torno a 1, las propias limitantes geográficas determinaron que el IF específico para el área de referencia no pueda bajar nunca de 5,0 (Fig. 5). La ganancia que ofrecería la restauración del área sería solo de 12%.
El índice de continuidad espacial (ICE) solo presentaría una leve mejoría en el PR, debido también al efecto que tiene que 95% del bosque se agrupe en un solo polígono. El ICE determinado como teórico en la tabla 6, supone que el área fuera circular, es decir, el perímetro se reduciría sustantivamente respecto al que ofreció el PR y mostró el máximo del índice para la magnitud del área involucrada, pero que no puede ser alcanzado debido a forma en que se distribuye el bosque.
Para entender el comportamiento y la sensibilidad de los índices fue confeccionada la tabla 7, en la que se suponen configuraciones teóricas de polígonos, pero con las dimensiones de los encontrados en el parque. En el supuesto que el parque sólo tuviera un polígono de bosque (línea 1), el índice de continuidad ICE sería idéntico al del PR (Tabla 6), ello porque los fragmentos que se formarían en el PR posible provocan que la superficie esté prácticamente toda en un solo polígono. Así también, el supuesto que la superficie fuera circular (Tabla 7, línea 2) representaría el máximo teórico para la escala de superficie como la del parque, pero no se lograría por las limitaciones físicas de la cuenca. El caso de suponer que los cinco fragmentos finales del PR fueran equivalentes en superficie (línea 3) y de forma circular, determinan un ICE menor (menos bueno), al supuesto que sigan siendo cinco polígonos con la superficie posible en el PR y circulares. Es decir, el comportamiento del índice del paisaje ICE ofrece una relación directa respecto a las acciones de una restauración, vale decir los valores aumentan en forma absoluta según se provoque cambios por restauración.
Tabla 6. Resultados de índice de forma (IF) e índice de continuidad espacial (ICE) para los polígonos determinados en la tabla 5, tanto para la condición inicial como para el paisaje de referencia teórico.
| Condición analizada | Área inicial | Paisaje de referencia | Variación (%) |
|---|---|---|---|
| IF promedio ponderado | 5,76 | 5,10 | 12 |
| ICE | 5,07 | 5,17 | 1,9 |
| Bosque asimilado a un círculo (teórico) | 5,84 |
Tabla 7. Posibilidades teóricas de los índices de forma (IF) y de continuidad espacial (ICE) según diferentes configuraciones en las formas y los tamaños de los polígonos en el parque Tagua Tagua.
| Tipo de configuración | Uso | Superficie (m) | Radio (m) | Perímetro (m) | ICE | IF |
|---|---|---|---|---|---|---|
| 1 | Área real unida | 14 581 787 | 82 983 | 5,17 | 6,13 | |
| 2 | Área real circular | 14 581 787 | 2 154 | 13 536 | 6,98 | 1,00 |
| 3 | 5 círculos con superficie equivalente | 2 916 357 | 963 | 6 053 | 1,00 | |
| 2 916 357 | 963 | 6 053 | 1,00 | |||
| 2 916 357 | 963 | 6 053 | 1,00 | |||
| 2 916 357 | 963 | 6 053 | 1,00 | |||
| 2 916 357 | 963 | 6 053 | 1,00 | |||
| Totales | 14 581 787 | 30 269 | 6,18 | 1,00 | ||
| 4 | Áreas reales con forma circular | 13 805 874 | 2096 | 13 171 | 1,00 | |
| 367 940 | 342 | 2 150 | 1,00 | |||
| 270 146 | 293 | 1 842 | 1,00 | |||
| 11 1359 | 188 | 1 183 | 1,00 | |||
| 26 466 | 91 | 576 | 1,00 | |||
| Totales | 14 581 786 | 18 924 | 6,65 | 1,00 |
El índice de forma IF en cambio, en el primer supuesto en que toda el área fuera un polígono (Tabla 7; línea 1), aumenta respecto al obtenido en el PR (Tabla 6), lo que resulta contradictorio al comportamiento teórico propuesto en su elaboración, es decir mientras más cercano a 1 sería mejor la forma que los parches tendrían para la conservación. Es más, es notorio como el IF aumenta en la medida que la superficie del fragmento es mayor (Tabla 5). Por otra parte, en los supuestos de que los fragmentos fueran circulares (líneas 2, 3 y 4), el IF es constante en 1, es decir, es insensible al número de fragmentos y a su superficie.
Propuesta de presentación sintética
Para representar la métrica de la variación que tendría la restauración teórica del área de interés, se propone establecer algún formato que permita visualizar todos los índices de paisaje ensayados y su variación teórica (Tabla 8).
Tabla 8. Síntesis de todos los índices del paisaje analizados y sus correspondientes tendencias.
| Índices de Paisaje para evaluar un plan teórico de restauración | Condición inicial | Paisaje de referencia | |||||
|---|---|---|---|---|---|---|---|
| sup (ha) | sup (%) | sup (ha) | sup (%) | Variación por Restauración | Tendencia deseada | ||
| Efecto borde | Con influencia | 55 | 3,96 | 28 | 1,91 | Se reduzca | |
| Matorral antrópico | 26 | 1,89 | |||||
| Matorral natural | 28 | 2,03 | 28 | ||||
| Pradera antrópica | 1 | 0,04 | |||||
| Sin influencia | 1 318 | 96,04 | 1 430 | 98,1 | 1,9% | ||
| Diversidad del paisaje | n° Usos | H | n° Usos | H | |||
| 6 | 1,29 | 4 | 1,39 | Según objetivos de la restauración | |||
| Índice de forma (IF) | 5,8 | 5,1 | 12% | Se reduzca | |||
| I. de continuidad del paisaje (ICE) | 5,07 | 5,17 | 1,9% | Aumente | |||
Donde: sup = superfice, H = índice de diversidad espacial.
No se incluyó la información del efecto borde escenario 1; sin análisis, dado que debiera ser recomendable siempre establecer la condición inicial, realizando un análisis como el propuesto.
La condición inicial se refiere a los valores de los índices en la condición de línea base y luego su variación en el paisaje de referencia.
En la utilización de la síntesis para visualizar los efectos de una teórica restauración, los índices de efecto borde y de continuidad espacial ICE, mostraron una relación positiva frente a los cambios esperados con una potencial restauración, ya que su variación tomó la dirección que teóricamente se espera de ellos según el paisaje está más conservado.
Discusión
El área con potencial para ecosistemas forestales en el parque está en su mayoría ocupada por bosques de diferentes asociaciones propias de la zona. Las del tipo Siempreverde corresponden a un mosaico de etapas sucesionales, muy similar a la descripción de diversos autores para bosques originales del tipo (Veblen, 1985; Armesto, Villagrán y Arroyo, 1993; Donoso, 2015), reflejando su alto grado de conservación (Tabla 2). La fracción de bosque respecto al total está sobre lo que Rutledge (2003) considera necesario para la persistencia de las especies en un determinado hábitat (más de 40% de la superficie). Por otra parte, en esta cuenca, los eventos naturales alogénicos que provocan retrocesos en la sucesión afectarían en torno a 50% de la superficie de bosque. Al respecto, los autores que han descrito su dinámica no han cuantificado su proporción respecto al total, como la encontrada en el parque.
La proposición de un PR en el área resultó un ejercicio simple de construir, no obstante, requirió conocimientos previos de la dinámica de los bosques involucrados. Es posible que otras áreas de la región fuera del parque, con una degradación de larga data y con mayores impactos, requieran un fuerte esfuerzo para la determinación conceptual del PR. Como fue revisado, no existen estudios que hayan realizado este tipo de propuestas.
En la determinación del efecto borde, fue sustantiva la diferencia entre la aplicación del índice por defecto como escenario 1, e incorporar supuestos basados en la propia dinámica del bosque como en el escenario 2, lo que reafirma las recomendaciones de Badii y Landeros (2007), respecto a ajustar los índices para cada caso, por una parte, y las recomendaciones de Laurance et al. (2001), Harper et al. (2005) y Aragón et al. (2015) respecto a estudiar, en este caso para Chile, las magnitudes de influencias entre hábitat. En cualquier caso, en ambos escenarios, el área sin influencia o núcleo forestal tiene la cobertura suficiente para soportar a la mayoría de los riesgos potenciales, si se acepta el estándar propuesto por Environment Canada (2013), que indica que el núcleo debe superar 50%. Se debe destacar, además, que el PR siempre mantendrá 5% en efecto borde. No hay estudios que refieran al hecho de que en algunos casos nunca podrá eliminarse el efecto borde, y se propone que dicho efecto borde inevitable en un área recuperada sea llamado “efecto borde basal”.
La diversidad del paisaje, por otra parte, fue un índice complejo de interpretar cuantitativamente al considerar una hipotética restauración. En efecto, como lo señala Bradshaw (1965), las condiciones sucesionales tenderían a desaparecer, haciendo que las proporciones de superficie entre los Usos de bosque varíen, mientras que simultáneamente las acciones alogénicas propias del territorio provocarían destrucciones de bosque y en el mediano plazo aparecerían colonizaciones de etapas sucesionales primarias de bosque y, como lo recomienda Lindenmayer (1995), estás dinámicas de destrucción propias deben respetarse. En otras palabras, el índice de diversidad resultó difícil de incorporar como un parámetro para comparar la condición inicial y el PR. Súmese a ello que la restauración eliminaría Usos, que modifican la estructura de cálculo entre la condición inicial y la final. Nogués Bravo y Pérez Cabello (2001) indican que el índice de diversidad (ID) es buen indicador, pero no debe ser una única referencia, ya que sus mejores resultados serían largas extensiones vegetales con pequeñas machas de hábitat que aportan heterogeneidad, y lo visto en el PR es una tendencia a un bosque continuo y homogéneo. Los niveles alcanzados en el parque en particular fueron altamente coincidentes con lo descrito por Nogués Bravo y Pérez Cabello (2001), especialmente por aquellas comunidades de Pilgerodendron uviferum D. Don y de F. cupressoides en otras áreas del país.
Respecto a las relaciones de forma y continuidad, los valores de IF e ICE del bosque inicial en la cuenca total fueron muy similares a los determinados para el PR (Tabla 6), confirmando su alto grado de conservación. Probablemente este alto grado de conservación no haya permitido comprender mejor el comportamiento de estos índices, ya que en la condición inicial un solo fragmento concentró 95% de la superficie en un área de más de mil hectáreas, lo que confirmó lo propuesto por varios autores que indican que fragmentos de gran superficie poseen altos IF (Cochrane y Laurance, 2002; Pincheira-Ulbrich, Rau y Peña-Cortés, 2009; Watson, 2003) y, cómo se confirma en la tabla 5, los pequeños fragmentos arrojan bajos IF, dado que tienen formas menos complejas (Corrêa do Carmo et al., 2001; Echeverry y Rodríguez, 2006). Efectivamente podría ocurrir, a la luz de estos resultados, que una zona de dimensiones similares a las del parque, pero con bosque altamente fragmentado, poseyera un IF inicial mejor que el PR, contradiciendo, en cierta medida, la tendencia teórica ideal descrita para el índice (Lozano Botache et al., 2011; Didham y Ewers, 2012) lo que parece indicar que este indice es una medida más de tipo cualitativo que cuantitativo.
La continuidad, tanto del área inicial como del PR (Tabla 6) se puede considerar alta según los trabajos de Vogelmann (1995), quien midió continuidades mayores de 4,2 para bosques con una cobertura entre 80% y 100%, así también la métrica de la continuidad en el parque refuerza la propuesta de Corrêa do Carmo et al. (2001) que consideran 4,5 como reflejo de una buena continuidad espacial del bosque, y de Echeverry y Rodríguez (2006) quienes, en su simulación de recuperación de áreas naturales proponen como valores 4,1 como de alta conservación.
Finalmente, y en consideración a los resultados expuestos, para trabajos de restauración de paisajes extensos, parece razonable realizar el ejercicio de proponer un PR que permita cuantificar el esfuerzo y los efectos de las actividades implicadas en dicho proyecto, por una parte, y luego analizar un conjunto de índices del paisaje de manera de discernir el grupo que mejor describa el cambio en conservación que se espera de la restauración. Ello, porque cada índice tendrá un comportamiento determinado por el área en específico y podrá reflejar mejor o peor dichos efectos, y porque los valores iniciales alcanzados por cada índice podrán entenderse mejor si es conocido el máximo valor potencial real que podría alcanzar en las condiciones ideales del PR.
Conclusiones
La cuenca del parque Tagua Tagua, resultó un adecuado ambiente de prueba para índices del paisaje, dado su alto grado de pristinidad y, en definitiva, permitió construir de forma sencilla un paisaje de referencia.
Las condiciones específicas del territorio del área objetivo provocaron respuestas confusas de algunos índices del paisaje, dado que contradijeron los comportamientos teóricos ideales según fueran mejor conservados los polígonos donde se usaban.
En específico, la diversidad espacial proyectada de la condición inicial a la del paisaje de referencia, debió cambiar de escala al reducirse el número de usos, además de que las proporciones de superficie en cada nivel ideal, no corresponden a las del paisaje de referencia. El índice de forma, por otra parte, aumenta al mejorar la conservación a consecuencia del crecimiento de los polígonos por uniones debidas a restauración, lo que hace que no sea totalmente efectivo a escalas de paisaje.
El efecto borde en cambio genera una métrica que es consistente con la mejora en conservación postrestauración, además de entregar una métrica fácil de entender, por lo que sería un índice recomendable para cuantificar las expectativas de cambio por restauración. No obstante, fue determinada la permanente existencia de un efecto borde basal, es decir, un mínimo que no puede eliminarse.
Así también, el índice de continuidad es consistente en su comportamiento teórico ideal y el representado por la condición de mejor conservación. A pesar que su valor es adimensional, el máximo generado por el paisaje de referencia permite usarlo como medida cuantitativa de la conservación.

