











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El bosque tropical caducifolio (BTC) en México es también llamado: selva baja decidua (Miranda, 1952), selva baja caducifolia (Flores, Jiménez, Madrigal, Moncayo y Takaki, 1971; Miranda y Hernández, 1963; Pennington y Sarukhan, 2005; Rzedowski, 1991) y bosque tropical deciduo (Rzedowski, 1996). El BTC en México contiene 31% de carbono almacenado (Becerril-Piña, Gonzáles-Sosa y Hernández-Sandoval, 2007). Se caracteriza por ser rico en especies arbóreas que operan en los distintos servicios del ecosistema (Cuevas, Núñez, Guzmán y Santana, 1998; Sánchez-Velásquez et al., 2002), como son: contribución a la regulación de la ocurrencia de temperaturas extremas altas en la temporada de seca (Ledesma-Colunga, Cambrón-Sandoval, Suzán-Azpiri, Guevara-Escoba y Luna-Soria, 2015) y capacidad de almacén de carbono (Chave et al., 2005). La diversidad de especies tiene un papel importante en los servicios ambientales que ofrece el bosque al presentar una relación; diversidad de especies con la capacidad de ofrecer servicios asociados a la regulación climática (Balvanera et al., 2006; Cardinale et al., 2011; Quijas, Schmid y Balvanera, 2010) y almacenamiento de carbono (Balvanera et al., 2006; Potvin y Gotelli, 2008; Ruiz-Jaen y Potvin, 2010).
Este tipo de bosques, además de ser diverso en especies de árboles, presenta una amplia gama de atributos como son la formas y tamaños, arquitectura de crecimiento específica de cada especie y árbol, que pueden ser útiles para un posible manejo, además de estar relacionados con los predictores de la biomasa de cada árbol, que se puede calcular con ayuda de ecuaciones alométricas comúnmente utilizadas para estimar la biomasa a escala individual o comunitaria (Zhao, Popescu y Nelson, 2009, Sullivan, Ducey, Orwig, Cook y Palace, 2017).
Se ha documentado la casi desaparición del BTC en la región Bajío de México (Rzedowski y Calderon, 1987), pero el estado actual es incierto (Trejo y Dirzo, 2000). El estado de Querétaro es parte de la región Bajío, donde existe una fuerte presión de cambio de uso de suelo por las empresas constructoras. Esta intensa actividad representa repercusiones significativas en la disminución de la superficie del BTC típico de la región bajío (Hernández-Vargas, Sánchez-Velásquez, Carmona-Valdovinos, Pineda-López y Cuevas-Guzmán, 2000; Sánchez-Velásquez et al., 2002; Stork y Samways, 1995). La disminución del bosque puede traer consigo la pérdida de especies de árboles con usos potenciales con beneficios económico y ecológico, usos que hoy en día son desconocidos o poco aprovechados y que pueden ser una alternativa en el futuro.
Los bosques y cada especie de árbol son de suma importancia, debido a que son transcendentales para mitigar el daño ocasionado por el uso de combustibles fósiles, principales gases causantes del cambio climático global (Becerril-Piña, Gonzáles-Sosa y Hernández-Sandoval, 2007; Ordóñez y Masera, 2001). Estos bosques además de contener carbono almacenado siguen participando en la fijación carbono atmosférico (CO2) (Becerril-Piña, Gonzáles-Sosa E. y Hernández-Sandoval, 2007; Gasparri y Manghi, 2004)
Conocer las principales especies de árboles que componen el BTC típico de cada región en México, su abundancia, las características de cada especie y la capacidad de captura de carbono almacenado en cada árbol es de suma importancia. Al generar un antecedente, influye en la toma de decisiones para la elaboración de estrategias de conservación y manejo a escala de especie, como por ejemplo en los inventarios de la vegetación arbórea, además de identificar y contabilizar las especies en sitios específicos. Hoy en día, ese conocimiento también ha permitido valorar diversos servicios ambientales a través de la evaluación de los atributos de las especies; uno de ellos es la capacidad de almacenamiento de carbono mediante la estimación de la biomasa aérea de los árboles (Chave et al., 2004; Gracia, Vayreda, Sabaté e Ibáñez, 2004; Houghton, 2003). Cabe mencionar que la biomasa de un árbol en promedio es de 46% de carbono, del cual 86% en su mayoría se encuentra en el fuste (Avendaño, Acosta, Carrillo y Etchevers, 2009).
Para la estimación de biomasa se recomienda utilizar modelos de regresión específicos para especies de árboles tropicales, donde se proponen modelos basados en atributos de crecimiento de los arboles (altura, DAP, diámetro basal, densidad de la madera) integrando las variables climáticas del sitio donde habitan (temperatura, precipitación e índice de aridez), estos parámetros reducen el grado de error de la estimación de carbono (Chave et al., 2004; Chave et al., 2005). Se considera que la relación diámetro-altura del árbol depende linealmente de una variable de estrés bioclimático que incluye los índices de variabilidad de la temperatura, variabilidad de la precipitación y la intensidad de la sequía, que permite la comparación entre árboles de una especie y entre especies (Bauer, Apps, Bhatti y Lal, 2006; Chave et al., 2014). Con este estudio se pretende contribuir al conocimiento específico de la composición, la diversidad y el carbono almacenado de árboles y arbustos, información que pueda ser aplicada en la planeación de actividades de reforestación y conservación del BTC típico del municipio de Querétaro, así como en los programas de pago de servicios ambientales por almacenamiento de carbono (Ávila, Jiménez, Beer, Gómez e Ibrahim, 2001; Chomitz, Buys, De Luca, Thomas y Wertz-Kanounnikoff, 2007; Esquivel, 2016). Se describen atributos de composición del BTC y se aporta información sobre los atributos específicos de las especies de árboles y arbustos que lo componen, atributos que se recomienda sean considerados en la planificación de la normativa para la reforestación o conservación, dependiendo de la necesidad y el objetivo. Por ejemplo, se proporciona información de especies con mayor capacidad de captura carbono, si el objetivo de un programa de reforestación o conservación en ambientes típicos de BTC fuera incrementar la capacidad de captura de carbono; lo anterior contribuye a la selección de especies con características idóneas para aumentar captura de carbono (Chave et al., 2005), debido que en múltiples ocasiones la reforestaciones del boque tropical caducifolio presenta dificultad para conformar un ensamble ecológico con el total de especies presentes.
Objetivos
Identificar las principales especies de árboles y arbustos del bosque tropical caducifolio del municipio de Querétaro, Qro., así como los diferenciales en abundancia y atributos de crecimiento entre especies presentes en él.
Estimar la biomasa y el carbono almacenado en dicho BTC, con ayuda de la ecuación alométrica propuesta por (Chave et al. 2005; 2014) e integrando los atributos de los árboles y arbustos a escala de especies y las variables climáticas específicas de los sitios donde habitan.
Materiales y métodos
Descripción del área de estudio
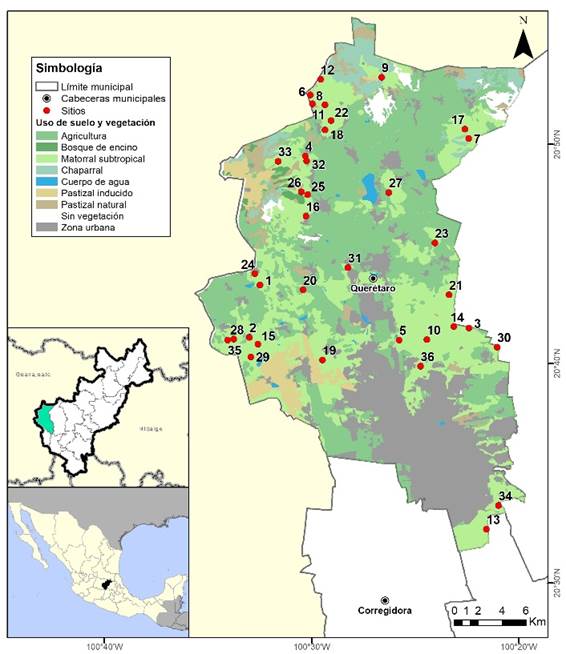
El sitio de estudio se localiza en el municipio de Querétaro, Querétaro (Fig. 1), México, dentro de las coordenadas extremas 20º 31’ a 20º 56’ de latitud norte y de 100º 19’ a 100º 36’ de longitud oeste. El área de estudio cuenta con vegetación predominante del bosque tropical caducifolio: bosque caducifolio espinoso (41 500 ha), bosque esclerófilo caducifolio (20 875 ha), pastizal mediano abierto (21 575 ha) y matorral crasicraule (17 750 ha) (Instituto Nacional de Estadística y Geografía [Inegi], 2014). El clima del municipio es templado semi-seco, caracterizado por un verano cálido. La temperatura media anual es de 18º C. Los meses más calurosos son mayo y junio. La precipitación pluvial anual promedio es de 555 mm (Instituto Nacional para el Federalismo y el Desarrollo Municipal, [Inafed], 2010). La recolección de datos se realizó en el periodo de enero a julio del 2016, utilizando un sistema aleatorio de selección de puntos.
Selección de cuadrantes de muestreo
Los 36 sitios de muestreo se seleccionaron de forma aleatoria a partir de una matriz de coordenadas geográficas de ubicación de puntos distribuidos dentro del área del municipio de Querétaro (Tabla 1). Se utilizó el programa Arcmap 10 ®Esri, para la generación de coordenadas de sitios de muestreo aleatorio; al programa se le indicaron tres criterios de exclusión para la localización de los puntos de muestreo: 1) zona rural; 2) cada punto debe estar a 100 m de cualquier vialidad o/y casa habitación; 3) una distancia mínima de 200 m entre cada punto. Para este procedimiento, el programa utilizó una capa de vegetación y uso de suelo serie V del Inegi (2014), en la cual se clasifica de manera más específica el uso de suelo (Tabla 1).
Tabla 1 Ubicación y descripción climática contemporánea (1961-1991) y futura (2030) de los sitios de muestreo del BTC del municipio de Querétaro, Qro.
| Sitio | Longitud O (Grados) | Latitud N (Grados) | Altitud (m) | MAT (°C) | MAP (mm) | IAA | ∆T (°C) | smrpb (mm) | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cont | 2030 | Cont | 2030 | Cont | 2030 | Cont | Cont | ||||
| 1 | 100.3218’ | 20.4666’ | 2317 | 15.7 | 17.2 | 709 | 629 | 0.088 | 0.106 | 11.3 | 2.2 |
| 2 | 100.2643’ | 20.4811’ | 2209 | 16.2 | 17.7 | 685 | 607 | 0.093 | 0.112 | 11.5 | 1.9 |
| 3 | 100.3186’ | 20.4331’ | 2218 | 16.2 | 17.7 | 696 | 617 | 0.092 | 0.110 | 11.6 | 2.1 |
| 4 | 100.2655’ | 20.4839’ | 2183 | 16.3 | 17.8 | 679 | 603 | 0.095 | 0.113 | 11.6 | 2 |
| 5 | 100.2623’ | 20.5286’ | 2184 | 16.4 | 17.8 | 666 | 591 | 0.096 | 0.115 | 11.8 | 2.2 |
| 6 | 100.2455’ | 20.4466’ | 2152 | 16.4 | 17.9 | 682 | 608 | 0.094 | 0.112 | 11.8 | 2.1 |
| 7 | 100.2107’ | 20.3993’ | 2133 | 16.4 | 17.9 | 692 | 613 | 0.093 | 0.111 | 11.9 | 2.1 |
| 8 | 100.221’ | 20.4129’ | 2118 | 16.5 | 18.0 | 686 | 608 | 0.094 | 0.113 | 11.9 | 2.1 |
| 9 | 100.2217’ | 20.4111’ | 2113 | 16.5 | 18.0 | 687 | 607 | 0.094 | 0.113 | 12.1 | 2.1 |
| 10 | 100.3931’ | 20.7108’ | 2226 | 16.6 | 18.1 | 586 | 524 | 0.111 | 0.131 | 12.1 | 2 |
| 11 | 100.2662’ | 20.4825’ | 2126 | 16.6 | 18.1 | 672 | 596 | 0.097 | 0.116 | 12.2 | 2.1 |
| 12 | 100.2836’ | 20.5182’ | 2116 | 16.7 | 18.2 | 659 | 586 | 0.099 | 0.118 | 12.2 | 2.1 |
| 13 | 100.2401’ | 20.4283’ | 2083 | 16.7 | 18.2 | 678 | 604 | 0.096 | 0.114 | 12.3 | 2.1 |
| 14 | 100.3199’ | 20.4288’ | 2094 | 16.8 | 18.2 | 681 | 603 | 0.096 | 0.115 | 12.2 | 2.1 |
| 15 | 100.2101’ | 20.3342’ | 2017 | 16.8 | 18.3 | 687 | 609 | 0.095 | 0.114 | 12.3 | 2.2 |
| 16 | 100.4474’ | 20.8839’ | 2241 | 16.8 | 18.3 | 490 | 443 | 0.134 | 0.157 | 12.4 | 2.1 |
| 17 | 100.2412’ | 20.4294’ | 2068 | 16.8 | 18.2 | 676 | 601 | 0.097 | 0.115 | 12.4 | 2.1 |
| 18 | 100.2292’ | 20.4038’ | 2046 | 16.8 | 18.3 | 680 | 602 | 0.096 | 0.115 | 12.3 | 2 |
| 19 | 100.33’ | 20.4159’ | 2060 | 16.9 | 18.4 | 679 | 604 | 0.097 | 0.115 | 12.5 | 2.2 |
| 20 | 100.4808’ | 20.8613’ | 2221 | 16.9 | 18.3 | 496 | 448 | 0.132 | 0.155 | 12.5 | 2.1 |
| 21 | 100.3021’ | 20.4297’ | 2057 | 16.9 | 18.4 | 676 | 600 | 0.097 | 0.116 | 12.5 | 2.1 |
| 22 | 100.3191’ | 20.4267’ | 2035 | 17.0 | 18.5 | 674 | 598 | 0.098 | 0.117 | 12.5 | 2.1 |
| 23 | 100.3304’ | 20.4144’ | 2038 | 17.0 | 18.5 | 677 | 603 | 0.098 | 0.116 | 12.6 | 2.1 |
| 24 | 100.3236’ | 20.4199’ | 2024 | 17.1 | 18.5 | 676 | 599 | 0.098 | 0.117 | 12.6 | 2.1 |
| 25 | 100.2149’ | 20.3811’ | 1948 | 17.1 | 18.6 | 668 | 591 | 0.099 | 0.119 | 12.5 | 2.1 |
| 26 | 100.2934’ | 20.4358’ | 2013 | 17.1 | 18.6 | 669 | 595 | 0.099 | 0.118 | 12.6 | 2 |
| 27 | 100.3036’ | 20.4268’ | 1989 | 17.2 | 18.6 | 669 | 594 | 0.099 | 0.118 | 12.8 | 2.1 |
| 28 | 100.4003’ | 20.7585’ | 2065 | 17.2 | 18.7 | 545 | 491 | 0.123 | 0.143 | 12.9 | 2.1 |
| 29 | 100.2212 | 20.3627’ | 1864 | 17.3 | 18.8 | 654 | 578 | 0.102 | 0.122 | 12.8 | 2 |
| 30 | 100.2701 | 20.4101’ | 1892 | 17.4 | 18.8 | 656 | 585 | 0.102 | 0.121 | 13 | 2 |
| 31 | 100.2757 | 20.4119’ | 1883 | 17.4 | 18.9 | 655 | 582 | 0.102 | 0.122 | 13 | 2 |
| 32 | 100.2836 | 20.3856’ | 1857 | 17.4 | 18.9 | 659 | 582 | 0.102 | 0.122 | 13 | 2.1 |
| 33 | 100.2995 | 20.409’ | 1914 | 17.4 | 18.8 | 665 | 589 | 0.101 | 0.120 | 13 | 2.1 |
| 34 | 100.2808 | 20.4115’ | 1888 | 17.4 | 18.9 | 656 | 582 | 0.102 | 0.122 | 13 | 2.1 |
| 35 | 100.3684 | 20.6228’ | 1959 | 17.5 | 19.0 | 604 | 539 | 0.111 | 0.132 | 13 | 2 |
| 36 | 100.3447 | 20.5886’ | 1925 | 17.6 | 19.0 | 614 | 548 | 0.110 | 0.130 | 13.5 | 2.1 |
MAT: Temperatura media anual, MAP: precipitación media anual, IAA: índice de aridez, Cont.: Clima contemporáneo, promedio 1961-1990, 2030: años 2030 (en un escenario intermedio CGCM3_A1B_2030), ∆T: cambio de temperatura, smrpb: diferencial de precipitación.
Modelación climática
La variabilidad climática contemporánea (1961-1991) de cada punto seleccionado se estimó las variables climáticas de temperatura media anual (MAT), precipitación media anual (MAP), balance de precipitación primavera/verano (smrpb (precipitación de julio + agosto / precipitación de abril y mayo)) e índice anual de aridez (IAA = (DD50.5)/ PMA; DD5 = grados día > 5 °C). La estimación del clima futuro (2030) se realizó a partir de modelos establecidos por Edenhofer (2014), utilizando el modelo A1B (modelo intermedio; A altas emisiones y B bajas emisiones carbono escenario) (Crookston, 2010 y 2014) (Tabla 1), posteriormente ajustando el modelo en ANUSPLINE (Sáenz-Romero et al., 2010; Crookstron, 2010). También se estimó la variabilidad de la temperatura máxima y mínima temperatura anual por sitio (∆T (°C)). Las variables climáticas se obtuvieron mediante los modelos climáticos desarrollados para México (Crookston, 2010), los cuales se basan en la transposición de superficies de datos climáticos mensuales normalizados de temperatura (mínima, media y máxima) y precipitación, obtenidos de aproximadamente 4000 estaciones climatológicas en el sur de Estados Unidos, México, Guatemala, Honduras y Cuba mediante el software ANUSPLINE 4.0.
Localización, método de muestreo y medición de atributos
Se localizó y georreferenció cada sitio (Tabla 1). En cada punto de muestreo se delimitó un cuadrante de 25 m × 25 m utilizando una soga de 100 m. Dentro del cuadrante se identificó a las especies arbóreas y arbustivas (Martínez et al., 2015; Rzedowski y Calderon, 1987), solo se evaluaron individuos que presentaran una altura superior a 150 cm para asegurarse que fueran arboles ya establecidos (no se consideraron cactáceas ni suculentas). Se evaluaron y estimaron variables e índices de diversidad como el de crecimiento de los árboles: diámetro basal, diámetro a la altura del pecho (DAP, suma de diámetros de todos los ejes principales del tallo a 1.30 m), altura total del árbol, altura de la copa, cobertura de copa, así como variables de arquitectura, de crecimiento como: tipo de fuste, tipo de copa e índices de Simpson, Shannon-Wiener y la determinación de carbono (Fig. 2) (Organización de las Naciones Unidas para la Alimentación y la Agricultura [FAO], 2004).

Determinación de carbono almacenado
La ecuación alométrica para estimar biomasa aérea utilizada por Chave et al. (2005, ecuación 1), y Chave et al. (2014, ecuaciones 2 y 3), en donde se utiliza la relación diámetro-altura del árbol genérico, depende linealmente de una variable de estrés bioclimático que incluye los índices de variabilidad de la temperatura, variabilidad de la precipitación, la intensidad de la sequía y la densidad de la madera, un predictor de biomasa (Chave et al., 2015). La densidad de la madera de cada especie se obtuvo a través de bases publicadas en Verduzco (2016); Ordóñez et al. (2012); Ordóñez (2015); Villalón (1992), Chave et al. (2009) y Zanne et al. (2009), permitiendo la comparación de almacén de carbono entre especies.
donde:
Estimación de índices de importancia y diversidad ecológica
Se estimaron distintos índices de diversidad e importancia ecológica en cada sitio de muestreo, a partir del número de especies y de la cantidad de individuos por especie. Dichos indicadores fueron: índice de Simpson (Peet, 1974; Magurran, 1988; Lande, 1996). Índice de Shannon-Wiener (Shannon y Weaver, 1949) y valor de importancia (Magurran, 1988). Interpretando la representatividad de las especies dentro de cada sitio.
Análisis estadístico
Para determinar las posibles diferencias significativas entre los atributos de arquitectura de desarrollo, así como en las variables de crecimiento de las especies, se realizó análisis de varianza (ANOVA). Para estimar cómo son estas diferencias en sus atributos, se utilizaron pruebas múltiples de medias de Tukey (α = 0.5) y análisis de regresión entre atributos, utilizando el programa estadístico SAS (Versión 9.3).
Resultados
Diversidad de especies y atributos de especies
En cuanto a diversidad de especies y densidad de individuos dentro del municipio de Querétaro, los resultados evidenciaron que el BTC está compuesto de 22 especies de arbustos y árboles superiores a 150 cm de altura (Tabla 2), con un índice de diversidad Simpson de 0.77 y un índice de Shannon de 0.87.
Tabla 2 Diversidad y densidad de especies representativas del BTC dentro del municipio de Querétaro, Qro.
| Especie | No. Ind | Den. | Fre. | V. de I. | No*ha | D. M. (g/cm3) | Fuente |
|---|---|---|---|---|---|---|---|
| Acacia schaffneri (L.) Willd. | 146 | 0.006 | 0.389 | 17.6 | 64.9 | 0.73 | Villalón (1992) |
| Bursera fagaroides (Kunth) Engl. | 141 | 0.006 | 0.583 | 20.8 | 62.7 | 0.32 | Verduzco (2016) |
| Forestiera phillyreoides (Benth.) Torr. | 140 | 0.006 | 0.639 | 21.8 | 62.2 | 0.63 | Ordóñez et al (2015) |
| Karwinskia humboldtiana (Willd. Ex Roem. & Schult.) Zucc. | 140 | 0.006 | 0.722 | 23.3 | 62.2 | 0.53 | Ordóñez et al. (2015) |
| Dodonea viscosa (L.) Jacq. | 138 | 0.006 | 0.194 | 13.4 | 61.3 | 1.04 | Ordóñez et al. (2015) |
| Ipomoea murucoides Roem. Shult. | 133 | 0.006 | 0.528 | 19.2 | 59.1 | 0.03 | Ordóñez et al. (2015) |
| Leucaena leucocephala (Lam.)De Wit | 88 | 0.004 | 0.139 | 8.8 | 39.1 | 0.84 | Zanne et al. (2009) |
| Acacia farnesiana (L) Willd. | 82 | 0.004 | 0.417 | 13.5 | 36.4 | 0.73 | Ordóñez et al. (2012) |
| Eysenhardtia polystachya (Ortega) Sarg. | 56 | 0.002 | 0.167 | 7.1 | 24.9 | 0.7 | Ordóñez et al. (2015) |
| Bursera palmeri S. Watson | 48 | 0.002 | 0.222 | 7.5 | 21.3 | 0.33 | Zanne et al. (2009) |
| Acacia angustissima (Schltdl & Cham.) Benth | 46 | 0.002 | 0.083 | 4.8 | 20.4 | 0.85 | Ordóñez et al. (2015) |
| Senna polyantha (Moc. & Sessé ex Collad.) Irwin & Barneby. | 43 | 0.002 | 0.417 | 10.8 | 19.1 | 0.63 | Ordóñez et al. (2015) |
| Acacia pennatula (Schltdl &Cham.) Benth. | 38 | 0.002 | 0.083 | 4.2 | 16.9 | 0.73 | Ordóñez et al. (2012) |
| Lysiloma microphyllum Benth. | 28 | 0.001 | 0.111 | 4.0 | 12.4 | 0.88 | Ordóñez et al. (2015) |
| Celtis pallidaTorr. | 26 | 0.001 | 0.333 | 8.0 | 11.6 | 0.66 | Ordóñez et al. (2012) |
| Condalia velutina I. M. Johnst | 25 | 0.001 | 0.167 | 4.9 | 11.1 | 0.63 | (Ordóñez et al. 2015) |
| Salvia sp (arbustiva)celtis caudata | 25 | 0.001 | 0.028 | 2.3 | 11.1 | 0.63 | (Ordóñez et al. 2015) |
| Prosopis laevigata (Humb. & Bonpl. Ex Willd.) M.C. Johnst. | 24 | 0.001 | 0.250 | 6.3 | 10.7 | 0.70 | Ordóñez et al. (2015) |
| Albizzia occidentalis Brandegee. | 18 | 0.001 | 0.083 | 2.8 | 8 | 0.52 | Ordóñez et al. (2015) |
| Mimosa monancistra Bent. | 14 | 0.001 | 0.056 | 2.0 | 6.2 | 0.67 | Ordóñez et al. (2012) |
| Randia thurberi S. Watson | 7 | 0.000 | 0.028 | 1.0 | 3.1 | 0.86 | Ordóñez et al. (2015) |
| Erythrina coralloides DC. | 2 | 0.000 | 0.056 | 1.2 | 0.9 | 0.38 | Zanne et al. (2009) |
No. Ind: Número de individuos, Den.: Densidad, Fre.: Frecuencia, V. de I.: Valor de importancia, No*ha: Número de individuos por hectárea, D.M.: Densidad de la madera
Se observó en la composición de la vegetación típica del BTC en el municipio de Querétaro, que las especies con mayor densidad relativa fueron A. schaffneri, B. fagaroides, F. phillyreoides, K. humboldtiana, D. viscosa, I. murucoides, mientras que las especies de menor densidad relativa fueron R. thurberi y E. coralloides (Fig. 3). Las especies que presentaron un mayor valor de importancia ecológica en el bosque tropical caducifolio dentro del municipio de Queretaro son: K. humboldtiana, F. phillyreoides, B. fagaroides, I. murucoides y A. schaffneri. Otras especies que destacan por su valor de importacia son D. viscosa, A. farneciana, Senna polyantha, L. Leucocephala, Celtis palida y P. laevigata, especies que están en todos los sitios y presentan elevada abundancia. Por el contrario, las de menor valor de importacia ecológica fueron R. thurberi y E. coralloides, estas especies solo se encontraron en un sitio y con una abundancia muy baja (Fig. 3).
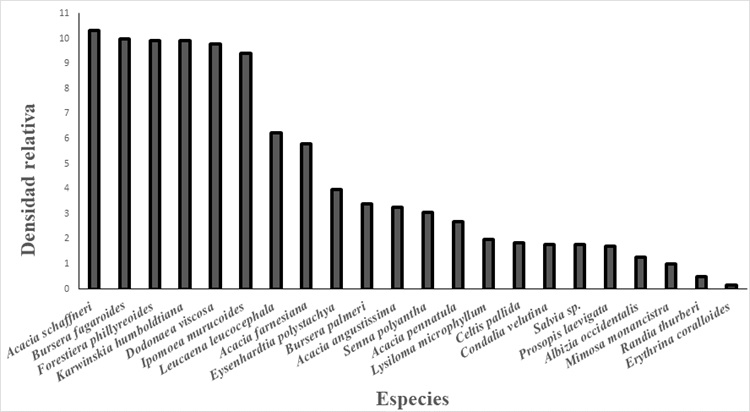
Figura 3 Comparación de la densidad relativa de arbustos y árboles nativos encontrados en el municipio de Querétaro, Qro.
Se observó que algunas de las especies presentaron un menor grado de importancia ecológica, sin embargo, la densidad relativa de dichas especies es elevada, como sucede con A. schaffneri, especie sobresaliente al ser una de las de mayor abundancia. Por otra parte, D. viscosa es una especie con alta desidad relativa y con un valor de importancia menor. Caso distinto es el de B. fagaroides, F. phillyreoides, K. humboldtiana y I. murucoides, con alta abundancia relativa y un elevado valor de importancia. Por otra parte, existen especies también importantes, pero poco abundantes con presencia en todos los sitios como S. polystachya, C. pallida, P. laevigata, y especies con poca abundacia y poca distribución como Acacia angustissima, Acacia pennatula, Lysiloma microphyllum, Condalia velutina, Albizzia occidentalis, Mimosa monancistra, R. thurberi, E. coralloides (Fig. 3 y 4).
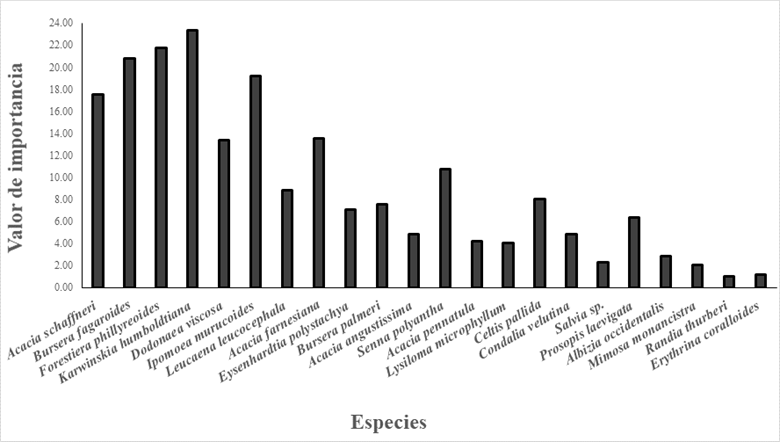
Figura 4 Comparación del valor de importancia de arbustos y árboles del bosque tropical caducifolio en el municipio de Querétaro, Qro.
Se estimaron diferencias significativas entre las especies para los atributos de forma como: altura total, altura de copa, diámetro basal, DAP, tipo de fuste, tipo de copa, biomasa aérea y carbono presente (P = 0.001, Tabla 3). Se observó que L. leucocephala fue la especie que presentó mayor diferenciación con respecto al resto de especies en las variables altura total (546.9 cm) y altura de copa (265 cm), copa más regular en comparación con las demás especies (Tabla 4; Fig. 5). Otras especies con mayor altura fueron: A. occidentalis, S. polyantha P. laevigata E. coralloides C. pallida L. microphyllum S. polyantha Bursera palmeri e I. murucoides, mientras que las especies que presentaron menor altura fueron: M. monancistra (233 cm) A. pennatula y A. angustissima. Por otra parte, R. thurberi y A. pennatula presentaron una copa más cercana al suelo.
Tabla 3 Análisis de varianza de variables de crecimiento, arquitectura y capacidad de captura de carbono del total de individuos evaluados del BTC del municipio de Querétaro, Qro.
| F. V. | g. l. | F | P |
|---|---|---|---|
| Altura | 21 | 42.37 | 0.0001 |
| Altura de copa | 21 | 21.26 | 0.0001 |
| Diámetro basal | 21 | 8.64 | 0.0001 |
| Diámetro altura del pecho | 21 | 9.73 | 0.0001 |
| Tipo de fuste | 21 | 13.52 | 0.0001 |
| Tipo de copa | 21 | 15.94 | 0.0001 |
| Biomasa | 21 | 14.81 | 0.0001 |
| Carbono | 21 | 14.81 | 0.0001 |
g.l.: grados de libertad, F: Valor de distribución de Fisher, P: significancia estadística
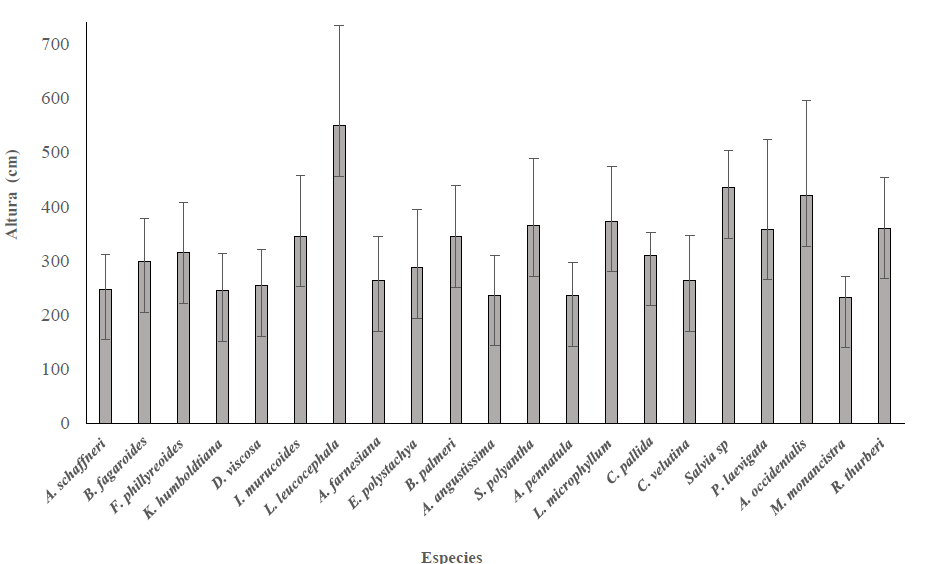
Figura 5 Altura promedio del total de individuos evaluados en el BTC del Municipio de Querétaro, Qro.
Respecto a diámetro del fuste basal y DAP, la especie que sobresale es: P. laevigata diferenciándose del resto de las especies en el grosor de su tallo. Por otra parte, D. viscosa es diferente de las demás al ser una especie abundante que presenta menores diámetros en su fuste (Tabla 4).
Tabla 4 Análisis múltiple de medias de Tukey (α = 0.05) de variables de crecimiento, arquitectura y capacidad de captura de carbono del total de individuos evaluados del BTC del municipio de Querétaro, Qro.
| Especie | Altura (cm) | Altura de copa (cm) | Diámetro basal (cm) | DAP (cm) | Tipo de fuste | Tipo de copa | Cantidad de carbono (kg / árbol) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Media | Tukey | Media | Tukey | Media | Tukey | Media | Tukey | Media | Tukey | Media | Tukey | Media | Tukey | |
| A. schaffneri | 247.1 | HIJ | 114.0 | BCDE | 19.3 | ABC | 25.7 | ABC | 2.0 | ABCD | 1.5 | BCDE | 5.2 | ABCDE |
| B. fagaroides | 299.0 | DEFGHIJ | 133.1 | BCDE | 19.8 | ABC | 21.0 | BCD | 1.7 | CD | 1.5 | BCDE | 3.1 | DE |
| F. phillyreoides | 314.7 | CDEFGHIJ | 134.7 | BCDE | 22.5 | ABC | 24.5 | ABCD | 2.4 | ABC | 2.0 | ABC | 5.4 | ABC |
| K. humboldtiana | 244.8 | HIJ | 117.4 | BCDE | 17.0 | ABC | 15.9 | BCD | 2.5 | AB | 2.1 | ABC | 4.0 | BCDE |
| D. viscosa | 254.1 | GHIJ | 109.0 | CDE | 8.8 | C | 8.3 | D | 2.1 | ABCD | 1.9 | ABCD | 4.0 | BCDE |
| I. murucoides | 345.4 | BCDEFGHI | 140.1 | BCDE | 26.3 | AB | 24.1 | ABCD | 1.9 | ABCD | 1.7 | BCDE | 3.7 | CDE |
| L. leucocephala | 546.9 | A | 265.0 | A | 20.9 | ABC | 20.6 | BCD | 2.0 | ABCD | 1.1 | E | 6.0 | AB |
| A. farnesiana | 262.9 | FGHUJ | 108.4 | CDE | 18.3 | ABC | 19.5 | BCD | 2.5 | AB | 1.9 | ABCD | 5.2 | ABCDE |
| E. polystachya | 287.3 | DEFGHIJ | 117.4 | BCDE | 14.7 | BC | 13.9 | CD | 2.1 | ABCD | 1.6 | BCDE | 4.5 | ABCDE |
| B. palmeri | 349.1 | BCDEFGHI | 139.0 | BCDE | 17.8 | ABC | 17.7 | AB | 1.4 | D | 1.2 | DE | 3.0 | E |
| A. angustissima | 238.0 | HIJ | 106.1 | CDE | 12.9 | BC | 18.9 | BCD | 2.1 | ABCD | 1.5 | BCDE | 4.7 | ABCDE |
| S. polyantha | 364.9 | BCDEF | 172.6 | BCD | 20.9 | ABC | 17.3 | BCD | 1.9 | ABCD | 1.4 | CDE | 5.0 | ABCDE |
| A. pennatula | 235.6 | IJ | 81.3 | E | 13.9 | BC | 13.9 | CD | 2.3 | ABC | 2.0 | ABC | 4.2 | ABCDE |
| L. microphyllum | 373.8 | BCDE | 186.0 | ABC | 17.4 | ABC | 18.7 | AB | 1.8 | BCD | 1.2 | E | 5.3 | ABC |
| C. pallida | 346.6 | BCDEFGH | 133.8 | BCDE | 19.3 | ABC | 19.3 | BCD | 2.3 | ABC | 1.9 | ABC | 5.0 | ABCD |
| C. velutina | 263.8 | EFGHIJ | 94.4 | BCD | 20.1 | ABC | 23.5 | ABCD | 1.8 | BCD | 1.8 | BCDE | 5.3 | ABCD |
| Salvia sp | 435.6 | B | 191.6 | AB | 23.4 | ABC | 22.5 | ABCD | 2.7 | A | 2.0 | ABC | 5.3 | ABC |
| P. laevigata | 358.7 | BCDEFG | 133.4 | BCDE | 30.7 | A | 38.6 | A | 2.1 | ABCD | 1.8 | BCDE | 6.4 | A |
| A. occidentalis | 420.6 | BC | 186.7 | ABC | 20.9 | ABC | 16.8 | BCD | 1.5 | D | 1.4 | CDE | 4.1 | BCDE |
| M. monancistra | 233.6 | J | 101.8 | DE | 18.1 | ABC | 17.8 | AB | 2.5 | AB | 2.2 | AB | 4.3 | ABCDE |
| R. thurberi | 317.5 | CDEFGHIJ | 87.5 | E | 15.8 | BC | 15.4 | CD | 2.5 | A | 2.5 | A | 5.3 | ABCD |
| E. coralloides | 395.0 | BCD | 144 | BCDE | 22.5 | ABC | 31.0 | AB | 1.5 | D | 1.5 | BCDE | 4.2 | BCDE |
Carbono almacenado
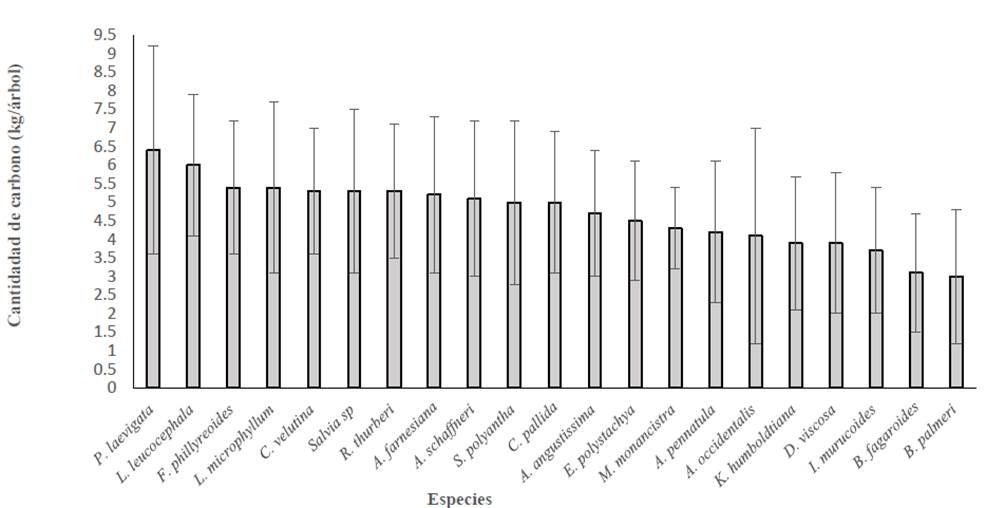
En las ecuaciones para la estimación del carbono almacenado se incluyeron las variables climáticas, así como variables de crecimiento (altura y diámetro) y la densidad de madera (Tabla 1, Tabla 2). Debido a lo anterior, con los resultados obtenidos se presumen relaciones positivas entre los atributos de los árboles y la captura de carbono; los coeficientes de determinación para dichas relaciones entre las variables de crecimiento y captura de carbono fueron: altura-contenido de carbono (r2 = 0.19), diámetro basal-contenido de carbono (r2 = 0.33) y DAP-contenido de carbono (r2 = 0.37). Se observó que P. laevigata aunada a L. leucocephala presentó la mayor cantidad de carbono almacenado con valores superiores en cobertura de copa y DAP, mientras B. fagaroides y B. palmeri fueron especies con menor carbono almacenado (Tabla 4, Fig. 6).
Discusión
El bosque tropical caducifolio del municipio de Querétaro se caracteriza por la existencia de una mayor abundancia de especies que dominan la estructura de dichos bosques. Según los resultados del índice de diversidad de Simpson (Shannon y Weaver, 1949), las especies con mayor presencia fueron: A. schaffneri, la cual, puede ser producto de su tolerancia a condiciones adversas y su capacidad para remplazar a otras especies (Flores y Yeaton, 2000). Por otro lado, la mayoría de las especies de estudio están presentes en todos los sitios según el resultado del índice de Shannon. El número de especies presentes en el municipio de Querétaro es similar al encontrado por Zacarías-Eslava, Cornejo-Tenorio, Cortés-Flores, González-Castañeda e Ibarra-Manríquez (2011) en la localidad de Cerro el Águila, en el estado de Michoacán. Estos autores encontraron 23 especies, de las cuales, pocas son iguales a las evaluadas en el presente estudio, como el caso de I. murucoides, F. phillyreoides, C. vetulina; sin embargo, los resultados publicados para la localidad Cerro del Águila difieren de lo estimado en el presente estudio respecto al índice de Simpson, al tener como resultado un valor 0.12 contra 0.77 del municipio de Querétaro. Por otro lado, en un estudio realizado en el estado de Chiapas (Rocha-Loredo, Ramírez-Marcial y González-Espinosa, 2010), se identificaron entre 13 y 18 especies en el BTC las cuales representan un número menor a las encontradas en este estudio (22 especies). Las diferencias en abundancia y riqueza de especies de los BTC en México pueden ser un indicador del estado del bosque, la presencia de un número superior de especies presume una mayor magnitud de contribución a algunos procesos del ecosistema (Balvanera et al., 2006; Cardinale et al., 2011; Quijas, Schmid y Balvanera, 2010), lo que ofrece servicios ambientales asociados a la regulación climática y al almacenamiento de carbono (Ledesma-Colunga, Cambrón-Sandoval, Suzán-Azpiri, Guevara-Escoba y Luna-Soria, 2015; Potvin y Gotelli, 2008; Ruiz-Jaen y Potvin, 2010). Con base en lo anterior y en los resultados de abundancia y riqueza de especies, el BTC del municipio de Querétaro se puede considerar en buen estado de conservación.
La composición de la vegetación típica del bosque tropical caducifolio del municipio de Querétaro es muy similar a la encontrada en el cerro El Águila en Michoacán, en este último sitio, I. murucoides y F. phillyreoides presentan los valores más elvados de importancia ecológica en comparación con el resto de las especies (Zacarías-Eslava et al., 2011). En el presente estudio, se observó un comportamineto similar, ya que algunas especies presetaron elavados valores de importancia ecológica como: A. schaffneri, B. fagaroides, F. phillyreoides, K. humboldtiana, D. viscosa e I. murucoides, en relación al resto de las especies evaluadas (16 especies restantes).
Si bien, históricamente la selección de mejores especies e individuos se basa en caracteres productivos y de crecimiento, dentro de la vegetación del bosque tropical caducifolio las especies presentan algunos atributos y usos diferentes a los productivos como lo son: medicinal, artesanal, forraje, ceremonial, etc. (Malda, Jiménez y Martínez, 2009; Sánchez et al., 2011), por lo cual conservar y aumentar la diversidad del bosque tropical caducifolio es primordial. Como ejemplo de lo anterior, se puede mencionar a E. coralloides, que es una especie que presenta poca abundancia, pero que debe ser prioritaria en planes de reforestación o conservación, ya que se considera una especie multipropósito al tener gran importancia ecológica y dentro de la medicina tradicional por la producción de alcaloides en sus tejidos (García-Mateos, Soto-Hernández y Kelly, 1998; García-Mateos, Soto-Hernández, Martínez-Vázquez y Villegas-Montero, 1999; Soto-Hernández y Miguel-Chavez, 2006). Por otra parte, la presencia de especies como A. pennatula, A. farnesiana, Eysenhardtia polystachya y I. murucoides, son conocidas como “especies típicas” de un estadio de sucesión estable del bosque tropical caducifolio (Labat, 1995; Rzedowski, 1978), da una idea del estado de conservación del bosque. Otro ejemplo es Leucaena leucocephala, especie con mayor altura, amplias copas y una mayor capacidad de almacenar carbono, entre otros atributos; sumado a lo anterior, se recomienda que estos árboles sean usados en sistemas silvo-pastoriles (Fiares, Olivera, Azevedo, Moura y Andrade, 2016) y en programas de restauración ecológica, por presentar rápido crecimiento, ayudar al establecimiento de otras especies y tener el potencial de aumentar la materia orgánica en suelos (Ceccon, Sánchez y Powers, 2015; Stewart y Dunsdon, 1994). Otra de las especies identificadas fue D. viscosa, que se caracterizó por ser muy abundante en los sitios de monitoreo del presente estudio; por sus atributos de arquitectura, velocidad de elongación y resistencia a la sequía, se recomienda su uso en reforestación (Asbjornsen, Vogt y Ashton, 2004).
Carbono almacenado
En la estimación del carbono almacenado, además de la densidad de la madera, se incluyeron las variables climáticas, que permitieron tener mayor precisión en la ecuación diámetro-altura del árbol; la densidad de la madera se presenta como una variable linealmente dependiente del estrés bioclimático (Chave et al., 2005; 2014). Se observó que P. laevigata presentó la mayor cantidad de carbono almacenado, con mayores copas y mayores diámetros a la altura del pecho. Complementariamente, se estimó que P. laevigata está asociado a una mayor diversidad de especies, lo cual hace posible que sea promotor del aumento en la cantidad de carbono (Yerena-Yamallel et al., 2015).
Acacia schaffneri presenta buena capacidad de almacenamiento de carbono en comparación con otras especies. Este resultado concuerda con lo observado por Yerena-Yamallel, Jiménez-Pérez, Aguirre-Calderón y Treviño-Garza (2011), en el matorral Tamaulipeco, estos autores encontraron que A. Schaffneri almacena más carbono con respecto a seis especies propias de la vegetación. Por otro lado, aunque A. farnesiana tiene poca presencia en el ecosistema, su carbono almacenado es elevado en comparación con el de otras especies típicas del bosque tropical caducifolio encontradas en este estudio. El uso de esta especie se ha recomendado en métodos de manejo de fitoremediación de suelos, en virtud de que los tallos pueden acumular metales pesados, por ello, es muy recomendable reforestar con estos árboles (Landeros-Márquez et al., 2017; Yerena-Yamallel et al., 2011). Adicionalmente, se ha propuesto para el control de la erosión en las zonas semiáridas (Vásquez-Méndez et al., 2010).
Las especies del bosque tropical caducifolio A. schaffneri, B. fagaroides, E. polystachya y K. humboldtiana han sido recomendadas en proyectos de reforestación ecológica a mediano y largo plazo; algunos autores han pronosticado en el futuro un incremento en sus áreas de distribución en el centro de México en un contexto de cambio climático (Gelviz-Gelvez, Pavon, Illoldi-Rangel y Ballesteros-Barrea, 2015). Leucaena leucocephala es otra especie que presenta buenos atributos para ser utilizada en programas de restauración ecológica y reforestación, al ser una especie medianamente abundante y con un valor de importancia ecológica intermedio, alcanza mayor altura, mayor carbono almacenado y ha sido recomendada y usada en sistemas silvopastoriles (Fiares et al., 2016).
Otra especie importante que presentó poca densidad (0.083) y bajo almacenamiento de carbono (4.2 kg/árbol)) es A. pennatula; Sin embargo, en otros estudios se ha observado su sobresaliente producción de biomasa dentro de 27 especies de la zona seca de América central (Stewart y Dundson, 1994). De la misma forma, Mimosa monancistra presentó baja densidad (0.01) y fijación de carbono (4.1 kg/árbol), contrario a otros estudios; esta especie ha sido recomendada por presentar una elevada capacidad de eliminar el pireno (BaP) de suelos contaminados (Álvarez-Bernal Contreras-Ramos, Marsch y Dendooven, 2007). Dodonea viscosa puede ser una especie importante utilizándola como pionera en programas de reforestación, debido a que es típica de sucesión temprana porque se caracteriza por un rápido crecimiento además llega a producir hojas aún bajo condiciones de sequía (Asbjornsen et al., 2004).
Otras especies recomendadas para su uso en programas de reforestación son aquellas con poca presencia como R. thurberi y E. coralloides; de esta última solo se encontraron dos individuos durante el periodo de muestreo; sin embargo, dicha especie produce alcaloides en sus tejidos, con aplicabilidad en medicina tradicional (García-Mateos et al., 1998, 1999; Soto-Hernández y Miguel-Chavez, 2006).
Se observaron 22 especies de arbustos y árboles dentro las que destacan por su abundancia: A. Schaffneri, B. fagaroides, F. phillyreoides, K. humboldtiana, D. viscosa e I. murucoides; con mayor grado de importancia: K. humboldtiana, F. phillyreoides, B. fagaroides, I. murucoides y A. schaffneri. Por otra parte, con mayor altura y capacidad de fijación de carbono se registraron especies como P. laevigata y L. leucocephala y las menos abundantes R. thurberi y E. coralloides.
Conclusiones
Se estimaron diferencias significativas (P ≥ 0.05) entre especies del BTC del municipio de Querétaro, para el total de variables evaluadas.
Se observaron 22 especies de arbustos y árboles, siendo las más abundantes: Acacia schaffneri, Bursera fagaroides, Forestiera phillyreoides, Karwinskia humboldtiana, Dodonaea viscosa e Ipomea murucoides.
Se estimó que especies como Prosopis laevigata y Leucaena leucocephala, presentan la mayor capacidad de captura de carbono en el presente estudio.

