











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
Los proyectos de mitigación al cambio climático tienen como objetivo reducir los gases de efecto invernadero [GEI], como el bióxido de carbono [CO2)], ya sea para disminuir su producción mediante el uso de energías renovables, incrementando la eficiencia, o a través de su captura o secuestro (e.g., ayudando a fijar carbono con proyectos forestales). El secuestro de CO2 consiste en utilizar el proceso natural de la fotosíntesis para capturar el carbono (C) de la atmósfera y retenerlo en forma de biomasa de las plantas y suelo, además de transferirlo a otros compartimentos de almacenamiento, de tal modo que no pueda ser re-emitido a la atmosfera. Debido a que los bosques retienen más C del que liberan, se les ha reconocido como “sumideros” de C. El secuestro de C en los ecosistemas terrestres ha provocado gran interés a escala mundial, debido a que la mayoría de los suelos se encuentran por debajo de la capacidad máxima de almacenamiento de C (Lal, 2004). Al plantar árboles (particularmente en bosques tropicales y templados) se pueden remover grandes cantidades de CO2 del aire (hasta 15 t ha-1año-1, en bosques tropicales) en un tiempo relativamente corto, lo que, además, acarrea beneficios socio-ambientales (Lal, 2011).
Dado el potencial de secuestro de C de los bosques en crecimiento y la conveniencia de mantener el C almacenado en los ecosistemas terrestres para no agravar el problema de emisiones de GEI, se han promovido recursos finacieros mediante mecanismos como REDD+ (Reducción de Emisiones por Deforestación y Degradación Forestal) de la Convención Marco de las Naciones Unidas sobre el Cambio Climático (CMNUCC) y Mecanismos de Desarrollo Limpio (MDL) del Protocolo de Kioto (1997); así como en practicas de manejo sostenible del bosque y en el establecimiento de plantaciones. Llevar a cabo proyectos como los mencionados requiere contar con métodos de medición confiables que cumplan con las exigencias establecidas por el Panel Intergubernamental de Cambio Climático [IPCC] y que sean costo-eficientes para medir el almacenamiento y la captura de C. Si se considera que los inventarios de carbono contabilizan el C fijado en los ecosistemas al momento de las mediciones, la pregunta que prevalece es ¿cuál es la capacidad máxima o en qué medida los compartimentos de los ecosistemas terrestres y costeros de México (biomasa aérea, material leñoso en pie, hojarasca-mantillo, material leñoso caído, biomasa subterránea y suelo) pueden contribuir como sumideros de CO2? (Vesterdal, Ritter y Gundersen, 2002). Una forma de abordar la pregunta es a través de un enfoque de estudio de largo plazo, que permita entender el desarrollo natural de la vegetación en ambientes perturbados por actividades antropógenicas y causas naturales. Los procesos de sucesión ecológica de la vegetación pueden tomar décadas o cientos de años y el uso de cronosecuencias permite la evaluación de estos periodos largos de desarrollo (Walker, Wardle, Bardgett y Clarkson, 2010).
Diferentes investigaciones indican que las cronosecuencias predicen un aumento en la riqueza de especies, cobertura, estructura, densidad y biomasa de las plantas, en ciertas etapas seriales a través del tiempo (Chazdon et al., 2007), así como en los contenidos de C en el suelo debido a la relación suelo-vegetación en los bosques tropicales húmedos y secos secundarios (Walker et al., 2010; Ahirwal y Maiti, 2017). A partir de lo anterior, se planteó el objetivo de conocer las trayectorias sucesionales de los contenidos de C en la biomasa área, para lo cual se recopiló, revisó y se sintetizó la información disponible en los estudios de la vegetación con enfoque de cronosecuencias en la República Mexicana. Los resultados encontrados muestran patrones generales de los contenidos de C en la biomasa aérea de la vegetación terrestre, pero no se hallaron datos de ecosistemas costeros, ni se intentaron modelar los patrones temporales del C en la biomasa aérea.
Materiales y métodos
Se efectuó una revisión de literatura para localizar publicaciones en internet con información relacionada con los cinco almacenes y contenidos de C en ecosistemas terrestres y costeros de México. Los principales almacenes de C en los ecosistemas terrestres son biomasa aérea viva (árboles, arbustos y hierbas), biomasa subterránea viva (raíces), biomasa muerta en pie (árboles y arbustos muertos y tocones), biomasa muerta en piso (troncos, ramas y ramillas sobre la superficie), mantillo (capa de hojarasca y capa de fermentación o material descompuesto) y suelo (profundidad de 30 cm) (IPCC, 2003) (Fig. 1).
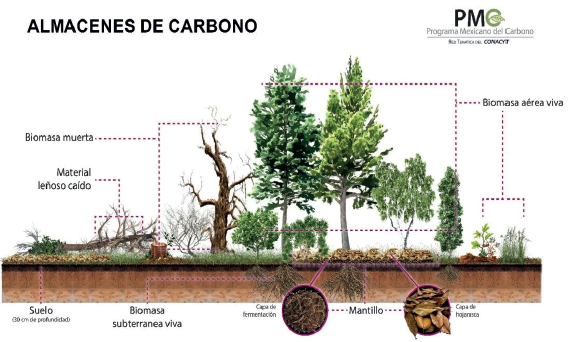
Fuente: Programa Mexicano del Carbono (PMC), adaptada de IPCC (2003)
Figura 1 Almacenes de carbono en las comunidades vegetales: 1) biomasa viva sobre el suelo, 2) biomasa subterránea [raíces], 3) biomasa muerta en pie, 4) biomasa muerta en piso y mantillo y 5) suelo.
Se utilizaron los motores de búsqueda públicos disponibles como Science Direct, Google Académico, Redalyc, Researchgate y Scopus. Los documentos encontrados se clasificaron en publicaciones convencionales y literatura gris. Las publicaciones convencionales se refieren a artículos científicos y libros indizados revisados por pares. La literatura gris son investigaciones plasmadas en tesis, informes y resúmenes en extensos publicados como resultado de reuniones científicas. La búsqueda se realizó mediante el uso de una serie de palabras clave con operadores lógicos para seleccionar la literatura relevante: cronosecuencias, índices de sitio, caracterización de la vegetación, etapas de sucesión, estimación de biomasa o C de los diferentes compartimentos, contenidos de C orgánico, materia orgánica, densidad aparente, propiedades físicas y químicas del suelo, fertilidad y caracterización de suelos, entre otros. La búsqueda se enfocó en trabajos de investigación realizados en el territorio nacional y sin restricción del periodo de publicación.
Los documentos resultantes se almacenaron en formato digital y se elaboraron dos archivos de Microsoft Excel® para sintetizar y sistematizar la información de cada documento. En el primer archivo se realizó un breve resumen de las diferentes publicaciones bajo el siguiente orden: identificador numérico secuencial del documento, tipo de documento (artículo, libro, capítulo de libro, tesis, tesina, informe técnico de proyecto, memoria de reunión científica, folleto o borrador), cita bibliográfica completa, título del artículo, autor (es), año de publicación, revista o editorial, resumen, coordenadas geográficas de los sitios, entidad federativa donde se realizó el trabajo, clima predominante, tipo de vegetación de acuerdo al Instituto Nacional de Estadística y Geografía [Inegi] y compartimento de biomasa con datos de C. El segundo archivo se integró con el mismo identificador numérico secuencial que el primer archivo para tener un campo vinculante entre ambos archivos y con los datos de contenido de C o biomasa, para posteriormente normalizarlos a toneladas de C por hectárea. Los contenidos de C se organizaron con base en su edad cronológica.
La captura de los datos a partir de los documentos localizados se realizó de forma manual; sin embargo, en algunos documentos los datos se presentaron en gráficos, por lo que se recurrió al software libre GetData Graph Digitizer para obtener datos numéricos aproximados mediante digitalización. Los datos se agruparon conforme a los grupos de vegetación propuestos por Challenger y Soberón (2008): bosques tropicales perennifolios (selvas altas y medianas perennifolias y subperennifolias del trópico húmedo); bosques tropicales caducifolios (selvas bajas y medianas caducifolias y subcaducifolias y selvas espinosas); bosques mesófilos de montaña (comunidades de gran diversidad en su estructura y composición, que comparten ambientes templados muy húmedos); bosques templados de coníferas y latifoliadas (bosques de pino, oyamel, ayarín, cedro, táscate, de encinos y mixtos de pino y encino); matorrales xerófilos (diversas comunidades vegetales dominantes en los climas áridos y semiáridos, i.e., matorrales rosetófilos, desérticos, crasicaules, submontanos y subtropicales, mezquitales y vegetación gipsófila y halófila); pastizales (pastizal natural, pradera de alta montaña, sabana y pastizales gipsófilos y halófilos) y humedales (manglar, bosque y selva de galería). No se encontraron datos para cronosecuencias de humedales.
Resultados y discusión
En el presente trabajo se identificaron 90 estudios de investigación en México relacionados con los contenidos de C con el enfoque de cronosecuencias. Geográficamente, 59% de los estudios se ubicaron en sur del país (incluido Veracruz), 22% se localizaron en las entidades de la zona centro y 19% en los estados del norte de la República Mexicana. Noventa y tres por ciento de los trabajos se circunscriben a alguna entidad federativa, mientras que seis trabajos presentaron datos a escala regional (Mejía, 2006; Urquiza, Dolman y Peres, 2007; Eaton y Lawerence, 2008; Návar, 2008; Yerena et al., 2014; Kauffman, Hernández, Jesús, Heider y Contreras, 2015). La revisión incluyó literatura convencional y gris. La mayor producción científica se concentra en los trabajos de artículos científicos. En esta revisión se incluyeron revistas indizadas indizadas o revisadas por pares (72), seguidas de trabajos de tesis que incluye los niveles de licenciatura, maestría y doctorado (16), en tanto que se identificaron dos informes (literatura gris). Las revistas Madera y Bosques, Revista Mexicana de Ciencias Forestales, Agrociencia y Revista Chapingo Serie Ciencias Forestales y del Ambiente fueron las editoriales con mayor número de artículos dedicados a la investigación de C en cronosecuencias. Los contenidos de C en cronosecuencias se publicaron por compartimento como sugiere el IPCC (2003). El compartimento más investigado fue el de biomasa viva sobre el suelo (56 estudios), seguido de la biomasa muerta en piso y mantillo (40). La revisión permitió conocer que solo seis investigaciones incluyeron datos de los cinco compartimentos (Jaramillo, Ahedo-Hernández y Kauffman, 2003a, Jaramillo, Kauffman, Rentería-Rodríguez, Cummings y Ellingson 2003b; Roncal, Soto, Castellanos, Ramírez y de Jong, 2008; Orihuela et al., 2013; Aryal, de Jong, Ochoa- Gaona, Esparza-Olguín y Mendoza-Vega, 2014; Masuhara et al., 2015).
En México, históricamente se ha hecho un esfuerzo creciente por investigar los contenidos de C en los cinco almacenes de los ecosistemas terrestres. Resulta importante señalar la variabilidad de enfoques con los que se han investigado los contenidos de C en cronosecuencias. El enfoque dado en las primeras investigaciones se perfiló medir la producción de biomasa y su relación con los nutrientes del suelo (Williams, 1983; Martínez y Sarukhán, 1993; Day et al., 1996; Hughes, Kauffman y Jaramillo, 1999). A partir de 1999, se mantuvo la idea de mediciones de producción de biomasa, pero bajo el método de ecuaciones alométricas. Los estudios desde 2004 sugieren temas de investigación diversos tales como medición de flujos de emisión y captura de CO2 (Návar, 2008), captura de C en suelo y vegetación (Vargas, Allen y Allen, 2009a; Vargas, Trumbore y Allen, 2009b; Casanova, Caamal, Petit, Solorio y Castillo, 2010; Figueroa, Ángeles, Velázquez y De los Santos, 2010; Gamboa et al., 2010, Mendoza-Ponce y Galicia, 2010; López-Hernández et al., 2013; Reyes, Méndez, Nájera y Cerano, 2013; Rivas, 2013; Rivera et al., 2013, Ramírez, 2015; Chávez-Aguilar et al., 2016; Huechacona, 2016; Mendoza-Morales, González-Sansón, y Aguilar-Betancourt, 2016; Chávez-Pascual, Rodríguez-Ortiz, Enríquez del Valle, Velasco-Velasco, y Gómez-Cárdenas, 2017). A partir de 2010, los estudios se dirigen hacia las estimaciones de contenidos de C para pagos por servicios ambientales a los poseedores de los recursos naturales (Moreno et al., 2010; Soto, Anzueto, Mendoza, Jiménez y de Jong, 2010; Aguirre y Jiménez, 2011; Castañeda, Vargas y Gómez, 2012; Espinoza, Krishnamurthy, Vázquez y Torres, 2012; Gómez, Monterroso, Tinoco y Etchevers, 2012; Gutiérrez, Méndez, Flores, Ramírez y Gutiérrez, 2012; Murray-Nuñez et al., 2012; Puc-Kauil, Ángeles, Plascencia y García, 2013; Razo- Zárate, Gordillo-Martínez, Rodríguez-Laguna, Maycotte-Morales y Acevedo-Sandoval, 2013; López-Reyes et al., 2016; Salvador et al., 2017) y hacia evaluaciones de impacto y mitigación al cambio climático (Pavón, Moreno y Ramírez-Bautista, 2012; Anguiano, Aguirre y Palma, 2013; Douterlungne, 2013; Soto y Armijo, 2014: Yerena et al., 2014). En los últimos años, las investigaciones se han orientado a la dinámica de cambio de uso de suelo y vegetación por deforestación y degradación forestal (Lawrence et al., 2013; Salazar, 2013; Aquino, 2014; Aryal et al., 2014; Puc-Kahuil, 2014, Romo, Navarro, De los Santos, Hernández y López, 2014; Anaya y Huber- Sannwald, 2015; Arévalo, 2015, Aryal, de Jong, Ochoa- Gaona, Mendoza-Vega, y Esparza-Olguín, 2015, Galicia, Saynes y Campo, 2015; Kauffman et al., 2015, Quintero- Gradilla, García-Oliva, Cuevas-Guzmán, Jardel-Pérez y Martínez-Yrízar, 2015; Yerena et al., 2015).
Contenidos de carbono en la biomasa viva sobre el suelo
Los contenidos de C en cronosecuencias encontrados en la literatura convencional y gris para México se presentan por tipos vegetación y condición sucesional primaria y secundaria, así como por agroecósistemas (Instituto Nacional de Estadística y Geografía [Inegi], 2009). Las claves de Inegi sin prefijo “VS” se refieren a la vegetación primaria, en donde la vegetación no presenta alteración. La vegetación secundaria es aquella que presenta indicios de que la vegetación nativa ha sido eliminada o perturbada sustancialmente. Las figuras 2 y 3 muestran los contenidos de C en la biomasa aérea de la vegetación de las zonas cálido-húmedas de México, que se definen como aquellas en donde se tienen temperaturas mínimas con promedio anual superior a 22 °C y precipitaciones superiores a 750 mm (Challenger y Soberón, 2008).
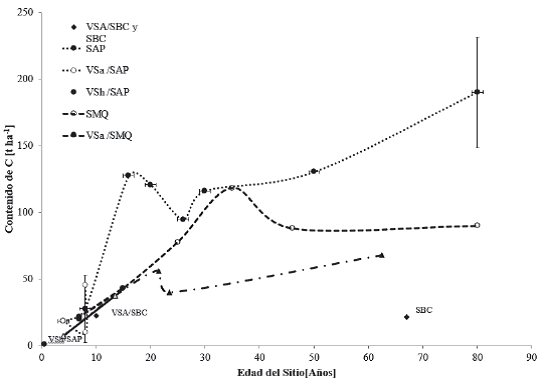
Figura 2 Contenido de carbono en cronosecuencias para vegetación secundaria arbórea de selva baja caducifolia (VSA/SBC) y vegetación primaria de selva baja caducifolia (SBC); vegetación primaria de selva alta perennifolia (SAP); vegetación secundaria arbustiva de selva alta perennifolia (VSa/SAP); y vegetación secundaria herbácea de selva alta perennifolia (VSh/SAP).
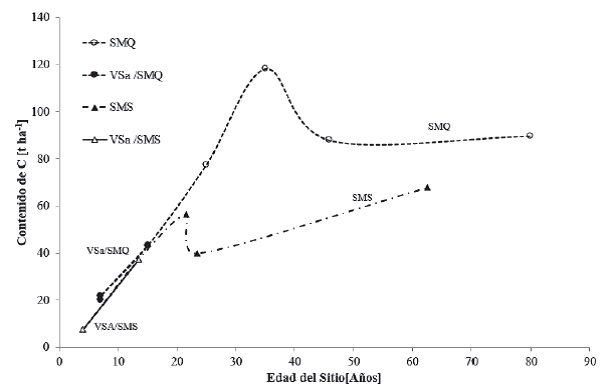
Figura 3 Contenido de carbono en cronosecuencias para vegetación primaria de selva mediana subperennifolia (SMQ); vegetación secundaria arbustiva de selva mediana subperennifolia (VSa/SMQ); vegetación primaria de selva mediana subcaducifolia (SMS); vegetación secundaria arbustiva de selva mediana subcaducifolia (VSa/SMS).
Los bosques cultivados (BC) son aquellos que se han establecido mediante la plantación de diferentes especies arbóreas, sobre todo en aquellas áreas que presentan una perturbación debido a las actividades humanas, mientras que los bosques inducidos (BI) son producto de reforestaciones sin un propósito comercial inicial. Estas poblaciones se pueden considerar como bosques artificiales, ya que son consecuencia de una reforestación con árboles de distintos géneros; por lo general, de especies exóticas. Los fines de estas plantaciones son recreativo, ornamental y forestal, además de conservar el medio ambiente y evitar la erosión del suelo (Inegi, 2009).
Los BC en los climas cálidos están representados por plantaciones forestales comerciales con especies de rápido crecimiento como eucalipto (Eucaliptus spp.) (Seppänen, 2002), hule (Hevea brasiliensis Müell. Arg.) (López-Reyes et al., 2016), melina (Gmelina arborea) y cedro rojo (Cedrela odorata L.) para las reforestaciones o bosques inducidos. Los proyectos más importantes se encuentran ubicados en el sureste de México (sur de Veracruz, Tabasco, norte de Chiapas, Campeche y Oaxaca), así como en las costas del Pacífico (Guerrero, Nayarit y sur de Sinaloa) (Seppänen, 2002).
Los bosques cultivados y los bosques inducidos los agrupa Inegi (2009) como parte de los agroecosistemas, cuyos datos de C en cronosecuencias para las zonas cálidohúmedas se observan en la figura 4.
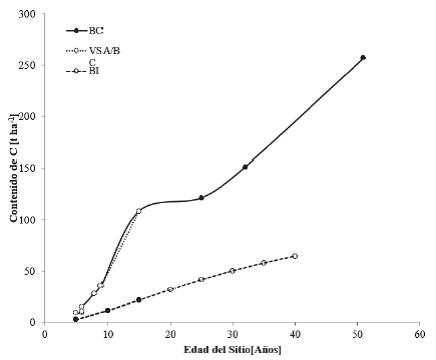
Figura 4 Contenidos de carbono en cronosecuencias para vegetación primaria de bosque cultivado de ciclo permanente (BC); vegetación secundaria arbórea de bosque cultivado de ciclo permanente (VSA/BC); vegetación primaria de bosque inducido (BI) y vegetación secundaria arbórea de bosque inducido (VSA/BI).
Los sistemas agroforestales (SAF) pueden ser una combinación entre cultivos agrícolas, pastizales y árboles forestales (Beer et al., 2003). Los contenidos de C en cronosecuencias para los tipos de vegetación donde se realizan actividades agropecuarias y forestales de las regiones cálido-húmedas se indican en la figura 5.

Agricultura de temporal anual (TA), pastizal natural (PN) y pastizal inducido (PI) de las zonas cálido-húmedas de México. Las barras verticales representan la desviación estándar, calculada a partir del promedio de los datos encontrados.
Las selvas caducifolias en México se desarrollan en climas trópico húmedo y subhúmedo, en condiciones de anegación total del suelo en la temporada de lluvias pero que se seca durante el estiaje, y constituyen el límite térmico e hídrico de los tipos de vegetación de las zonas cálido-húmedas. Se encuentran a lo largo de la costa del Pacífico (hasta el sur de Sonora en su límite norte), a lo largo de las faldas bajas y los cañones de la vertiente del Pacífico de la Sierra Madre Occidental, en las planicies, llanuras y otras zonas bajas e inundables de la Península de Yucatán y el sur de Veracruz y Tabasco. Se presentan en zonas con promedio de temperatura anual superior a 20 °C y precipitación anual de 1200 mm como máximo, aunque usualmente son del orden de 800 mm, con una temporada seca que puede durar hasta 7 u 8 meses (Challenger y Soberón, 2008).
Los contenidos de C en cronosecuencias para la vegetación de selva baja caducifolia (SBC), se muestra en la figura 6. Esta selva presenta baja altura de sus componentes arbóreos (normalmente, de 4 m a 10 m y, muy eventualmente, de hasta 15 m o poco más). El estrato herbáceo es bastante reducido y solo se puede apreciar después de que ha empezado claramente la época de lluvias y retoñan o germinan las especies herbáceas. Este tipo de vegetación es un importante centro de diversidad y endemismo de diferentes especies de Bursera y Fabaceae (Inegi, 2009).
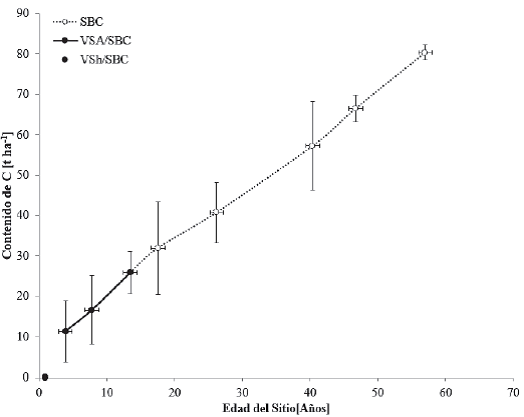
Las barras verticales representan la desviación estándar, calculada a partir del promedio de los datos encontrados.
Figura 6 Contenidos de carbono en cronosecuencias para vegetación primaria de selva baja caducifolia (SBC), vegetación secundaria arbórea de selva baja caducifolia (VSA/SBC) y vegetación secundaria herbácea de selva baja caducifolia (VSh/SBC) de las zonas tropicales de México.
Las selvas medianas caducifolias (SMC) se desarrollan en suelos que presentan mejores condiciones de humedad que los de la SBC. Challenger y Soberón (2008) consideraron que la vegetación de las SMC es resultado de siglos de explotación desde la época de los mayas, lo que le da un aspecto secundario. Los contenidos de C en cronosecuencias para este tipo de vegetación se observan en la figura 7.
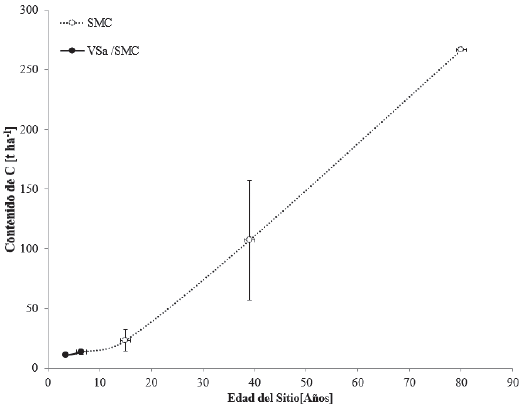
Las barras verticales representan la desviación estándar, calculada a partir del promedio de los datos encontrados.
Figura 7 Contenidos de carbono en cronosecuencias para vegetación primaria de selva mediana caducifolia (SMC) y vegetación secundaria arbustiva de selva mediana caducifolia (VSa/SMC) de la zonas cálido-húmedas de México.
Los contenidos de C en cronosecuencias para la selva mediana superennifolia (SMQ) se observan en la figura 8. En México, la selva mediana superennifolia se desarrolla en climas cálido-húmedos y subhúmedos: Aw para las porciones más secas, Am para las más húmedas y Cw en menor proporción (García, 1988). Con temperaturas típicas entre 20 °C y 28 °C y precipitación total anual de 1000 mm a 1600 mm, se ubica entre 0 m y 1300 m snm. Los árboles presentan alturas que van de 25 m a 35 m y diámetro a la altura del pecho menor que los de la selva alta perennifolia, aun cuando se trate de la misma especie. Es posible que esto se deba al tipo de suelo y a la profundidad. En este tipo de selva, se distinguen tres estratos arbóreos: de 4 m a12 m, de 12m a 22 m y de 22 m a 35 m (Inegi, 2009).
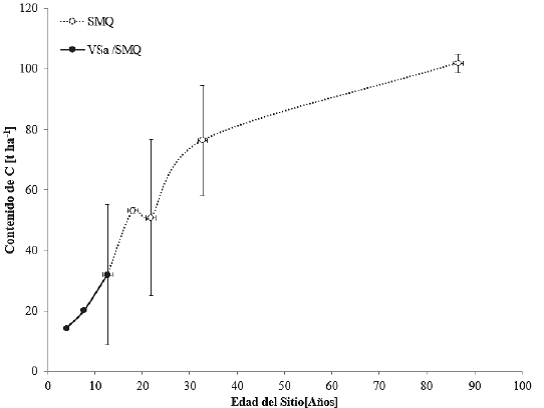
Las barras verticales representan la desviación estándar, calculada a partir del promedio de los datos encontrados.
Figura 8 Contenidos de carbono en cronosecuencias para vegetación primaria de selva mediana superennifolia (SMQ) y vegetación secundaria arbustiva de selva mediana superennifolia (VSa/SMQ).
En las áreas tropicales de México se han establecido plantaciones de bambú (Bambusa oldhamii Munro) para recuperar terrenos deforestados y obtener productos forestales, al mismo tiempo de darle el enfoque de captura de C (Castañeda et al., 2012). Por otra parte, se han establecido sistemas silvopastoriles con la utilización de especies leñosas forrajeras (Leucaena leucocephala (Lam) de Wit. y Guazuma ulmifolia Lam) como alternativa para el suministro de alimentos para el ganado en la época seca en zonas tropicales del sureste de México, además de darle el enfoque de captura de C (Casanova et al., 2010). Ambas se consideran dentro de la categoría de bosques cultivados. Los contenidos de C del BC en las zonas tropicales se muestran en la figura 9.
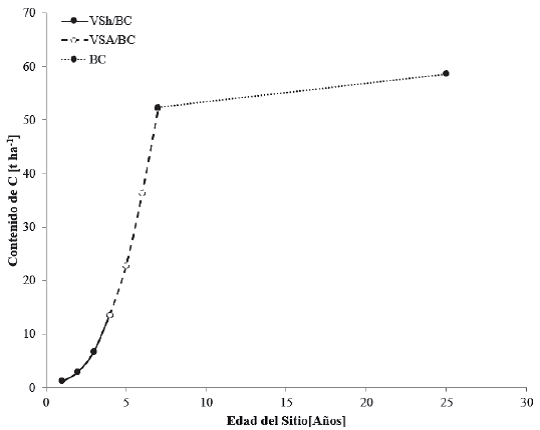
Figura 9 Contenidos de carbono en cronosecuencias para bosques cultivados (BC) y vegetación secundaria herbácea (VSh/BC) y arbórea (VSA/BC) de bosques cultivados.
En las zonas tropicales, la selva baja espinosa, incluyendo al matorral espinoso tamaulipeco (MET), se desarrolla en condiciones más secas que las correspondientes al bosque tropical caducifolio, pero, a su vez, más húmedas que las propias de los matorrales xerófilos. A diferencia del bosque tropical caducifolio que se restringe a cerros o laderas con suelos someros, la selva baja espinosa se desarrolla en suelos profundos (Rzedowski, 1996; Inegi, 2009). Los contenidos de C en cronosecuencias encontrados en este trabajo para el MET se muestran en la figura 10.
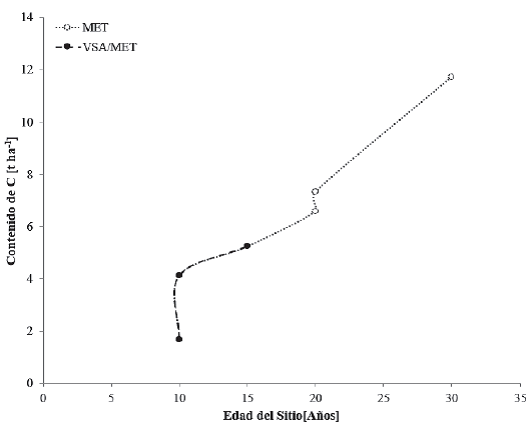
Figura 10 Contenidos de carbono en cronosecuencias para vegetación primaria de matorral espinoso tamaulipeco (MET) y vegetación secundaria arbórea de matorral espinoso tamaulipeco (VSA/MET).
Los bosques templados de coníferas y latifoliadas de México se encuentran, en su mayoría, en las zonas montañosas a lo largo de la Sierra Madre Occidental (la zona de mayor concentración de ecosistemas boscosos del país), las sierras Madre Oriental y del Sur de Chiapas, el Eje Neovolcánico, la Sierra Norte de Oaxaca y los Altos de Chiapas, así como en distintas serranías y montañas aisladas en el Altiplano y entremezclados en las planicies tropicales. De manera general, estos bosques se subdividen entre los de clima templado subhúmedo -que predominan en extensión- y los de clima templado húmedo. Los primeros se dividen en bosques de coníferas (de pino, abeto u oyamel, ayarín, cedro y táscate), bosques de latifoliadas (en los cuales los encinos son dominantes) y bosques mixtos de pino y encino, en distintas proporciones. Los bosques de clima templado húmedo son casi exclusivamente bosques mesófilos de montaña, aunque algunos bosques de oyamel pueden encontrarse en estas condiciones.
Los contenidos de C de la biomasa aérea en cronosecuencias asociados a bosques de pino (BP) se presentan en la figura 11. El Género Pinus se considera como uno de los principales dominantes en la vegetación primaria de los BP, aunque varias especies son pioneras predominantes tras las perturbaciones naturales y antrópicas (Challenger y Soberón, 2008).
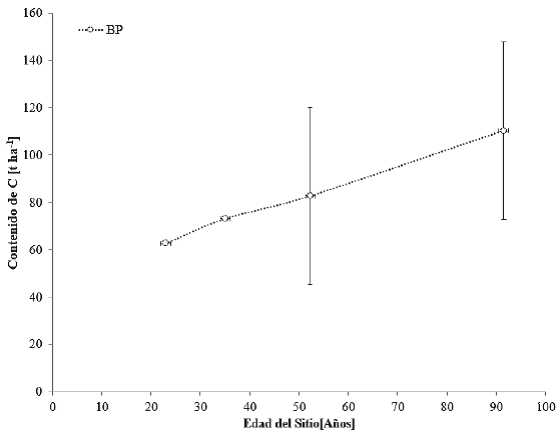
Las barras verticales representan la desviación estándar, calculada a partir del promedio de los datos encontrados.
Figura 11 Contenidos de carbono en cronosecuencias de la vegetación primaria de los bosques de pino (BP).
Los pinares primarios presentan un estrato inferior relativamente pobre en arbustos, pero con abundantes gramíneas. Esta condición se relaciona con los frecuentes incendios y la tala inmoderada. La fisonomía del estrato superior de los BP es característica y las diferentes especies de pino que los definen presentan alturas que van de 15 m a 30 m en promedio (Inegi, 2009). Estas diferencias se reflejan en la variabilidad de contenidos de C a partir de los 50 años (Fig. 11). Los bosques de pino, que con frecuencia se encuentran asociados con encinares y otras especies, son los de mayor importancia económica en la industria forestal del país, por lo que prácticamente todos soportan actividades forestales como producción de madera, resinación, obtención de pulpa para celulosa, postería y recolección de frutos y semillas. Los bosques de pino-encino (BPQ) y los de encino-pino (BQP) se consideran fases de transición en el desarrollo de bosques de pino o encino puros; sin embargo, Challenger y Soberón (2008) afirman que muchos de ellos se consideran vegetación clímax en muchas zonas de México. La recopilación de datos del contenido de C en cronosecuencias en BPQ para vegetación primaria y vegetación secundaria arbórea se observan en la figura 12.
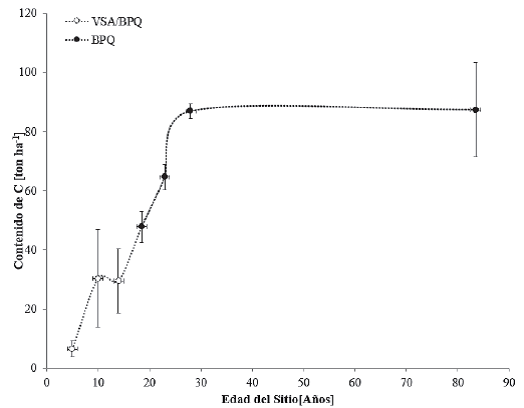
Las barras verticales representan la desviación estándar, calculada a partir del promedio de los datos encontrados
Figura 12 Contenidos de carbono en cronosecuencias de la vegetación primaria de los bosques de pino-encino (BPQ) y vegetación secundaria arbórea de pino-encino (VSA/BPQ).
Las especies que más se cultivan en los bosques cultivados de las zonas templadas son: pino (Pinus spp.), eucalipto (Eucaliptus spp.), cedro (Cupressus spp.), casuarina (Casuarina sp.), pirul (Schinus molle), álamo o chopo (Populus spp.), fresno (Fraxinus sp.) y aile (Alnus sp.) (Inegi, 2009). Las especies con datos sobre contenido de C en cronosecuencias para el BC de climas templados son Pinus greggii Engelm (Chávez-Pascual et al., 2017) y Pinus patula (Romo et al., 2014; Chávez-Aguilar et al.,2016) (Fig. 13).
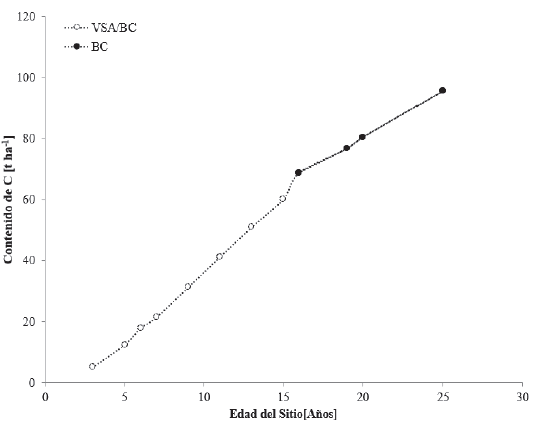
Figura 13 Contenidos de carbono en cronosecuencias de la vegetación primaria de bosque cultivados (BC) y vegetación secundaria arbórea de bosque cultivado (VSA/BC).
En los bosques mesófilos de montaña es notable la mezcla de elementos arbóreos con alturas de 10 m a 25 m o aún mayores. Es un bosque denso y la mayoría de sus componentes son de hoja perenne, pero también se encuentran árboles caducifolios que en alguna época del año tiran sus hojas. Es común la presencia de plantas trepado En los bosques mesófilos de montaña es notable la mezcla de elementos arbóreos con alturas de 10 m a 25 m o aún mayores. Es un bosque denso y la mayoría de sus componentes son de hoja perenne, pero también se encuentran árboles caducifolios que en alguna época del año tiran sus hojas. Es común la presencia de plantas trepadoras y epífitas debido a la alta humedad atmosférica y abundantes lluvias. Generalmente, se encuentran entre 800 m y 2400 m (Rzedowski, 1996; Inegi, 2009). Los contenidos de C asociados a bosque mesófilo se muestran en la figura 14.

Síntesis del contenido de carbono de la biomasa aérea en cronosecuencias
La Tabla 1 muestra los contenidos de C en cronosecuencias de la biomasa aérea para diferentes grupos de vegetación de México. Se observa un patrón de incremento en el tiempo en los contenidos de C para todos los grupos de vegetación. Las cantidades de C de la biomasa aérea son variables entre grupos. Los bosques tropicales perennifolios (Grupo de vegetación 1, Tabla 1) de entre 11 años y 20 años (64.077 t C ha-1 ± 48.564 t C ha-1) logran un incremento del almacenamiento de C hasta tres veces mayor que los mismos sitios de entre 1 año y 10 años (20.886 t C ha-1 ± 8.334 t C ha-1); es decir, en promedio capturan 4.31 t C ha-1 año-1 ± 4.02 t C ha-1 año-1. De la misma manera, los bosques templados de coníferas y latifoliadas (Grupo de vegetación 4, Tabla 1) de 11 años a 20 años (53.61 t C ha-1 ± 16.238 t C ha-1) incrementan 2.77 veces más sus contenidos de C en la biomasa con respecto a los sitios de la misma vegetación a la edad de 1 año a 10 años (19.298 t C ha-1 ± 7.536 t C ha-1). Se estima una tasa de captura de C en los bosques templados de coníferas y latifoliadas de México de 3.431 t C ha-1 año-1 ± 0.8702 t C ha-1 año-1, en sitios de 10 años a 20 años de edad.
Tabla 1 Contenido de carbono de la biomasa aérea (t C ha-1) por grupo de vegetación a diferentes edades.
| Vegetación | Edad del sitio (años) | ||||||
| Grupo | 1-10 | 11-20 | 21-30 | 31-40 | 41-50 | 51-60 | 61-95 |
| 1 | 020.8860 (008.334) |
064.0770 (048.564) |
067.4650 (076.915) |
076.2430 (075.563) |
108.9500 (030.053) |
162.0000 (133.980) |
087.950 (125.883) |
| 2 | 016.1320 (001.809) |
022.8490 (019.074) |
034.4300 | 082.1660 | 061.8250 | 080.2950 | 266.6000 |
| 3 | 015.9350 (000.458) |
037.8800 | - | 093.6300 | - | - | 295.5900 |
| 4 | 019.2980 (007.536) |
053.6100 (016.238) |
082.0500 (016.509) |
073.1800 | - | 082.7630 | 098.8460 (016.198) |
| 6 | 005.5880 (006.438) |
007.3500 (005.303) |
- | 014.0500 | - | - | - |
Se presentan valores promedio. Los datos en paréntesis son la desviación estándar (n = 4).
Los bosques mesófilos de montaña (Grupo de vegetación 3, Tabla 1) presentan la mayor cantidad de C almacenado en la biomasa, junto a los bosques tropicales caducifolios (Grupo de vegetación 3, Tabla 1): 295.59 t C ha-1 y 266.6 t C ha-1, respectivamente. Los pastizales alcanzan la menor cantidad de C en la biomasa.
La captura de C varía en función de las condiciones climáticas de la zona. La tasa anual total de captura de C en la biomasa aérea y dentro del suelo aumenta en las zonas boreales de 0.4 t C ha-1 año-1 a 1.2 t C ha-1 año-1; en las templadas, de 1.5 t C ha-1 año-1 a 4.5 t C ha-1 año-1; y en las tropicales, de 4 t C ha-1 año-1 a 8 t C ha-1 año-1 (Dixon, 1995). En tanto que Yepes, del Valle, Jaramillo y Orrego (2010), con base en un estudio realizado en la región de Porce, Antioquia, Colombia, hallaron diferencias significativas al comparar el contenido de la biomasa aérea en bosques con edades, en años, entre 7 y 11, 20 y 25 y 40 y 43, con el bosque primario; además, se observó mayor acumulación de la biomasa aérea total en bosques con edades de 20 años a 25 años.
Con base en los datos presentados en la tabla 1y en lo encontrado a escala internacional, en México se estima que la tasa de captura de C en los bosques templados de coníferas y latifoliadas es de 3.431 t C ha-1 año-1 ± 0.8702 t C ha-1 año-1, mientras que los bosques tropicales perennifolios capturan en promedio 4.31 t C ha-1 año-1 ± 4.02 t C ha-1 año-1. Ambas tasas ocurren en los sitios de 10 años a 20 años de edad y dentro de la etapa de crecimiento donde triplican los contenidos de C con respecto a los sitios de 1 año a 10 años, lo que se aproxima a lo registrado a escala mundial.
La rápida acumulación de biomasa se debe a que los bosques que tienen crecimiento neto son capaces de capturar más CO2 del que emiten a través de la respiración, y la velocidad de captura de C es directamente proporcional a dicho crecimiento. Por otra parte, los bosques maduros con poco crecimiento mantienen solo el C fijado y, además, experimentan una pérdida neta de biomasa por la mortalidad; Así mismo, ante la decadencia de la masa forestal, se convierten en emisores de CO2 (Kyrklund, 1991). Aunque Lewis et al. (2009) mostraron resultados de estudios en parcelas permanentes en bosques tropicales en América y África, que indican que los bosques intactos han funcionado como sumideros de C en las últimas décadas, en las que han presentado un aumento en la biomasa de una magnitud casi similar a las emisiones por deforestación (0.6 t C ha-1 en bosques amazónicos o 1.3 G t C ha-1 aproximadamente, si se consideran todos los bosques tropicales intactos).
Los bosques en crecimiento se refieren a los bosques secundarios que provienen, por un lado, de la modificación de bosques maduros producto de las actividades que abarcan desde la tradicional roza, tumba y quema hasta la agricultura y la ganadería intensiva que emplean especies exóticas, agroquímicos y maquinaria pesada. Y por otro lado, pese al pujante desarrollo agropecuario, existen eventos como la pérdida de la productividad, la urbanización y la migración en el ámbito rural, que han promovido el abandono de las zonas agropecuarias (Food and Agriculture Organization [FAO], 2005). Este proceso de abandono ha generado un importante aumento de bosques secundarios (Wright y Muller-Landau, 2006).
Los bosques secundarios y degradados en México ocupan 64% de la cobertura forestal de las zonas tropicales (FAO, 2010). La importancia de los bosques secundarios ante el cambio climático radica en su potencial como sumidero de C, por lo que es fundamental conocer la cantidad de biomasa que pueden almacenar y, a partir de esta, construir presupuestos globales de C, así como las políticas locales que propicien la ruta de su captura y almacenamiento (Becknell, Kucek y Powers, 2012).
La sucesión vegetal secundaria es un proceso ecológico por el cual se modifica la cobertura de los bosques secundarios y, conociendo su evolución, puede lograse conocer la cantidad de C capturado (Clark, 2007); es por ello que conocer la trayectoria sucesional es importante para planificar el uso, manejo y conservación de los ecosistemas. Las cronosecuencias son apropiadas para el estudio sucesional de la vegetación cuando existe evidencia de que los sitios de bosque secundario con diferentes edades siguen la misma trayectoria; es decir, existe un supuesto importante que cumplir, el cual establece que todos los sitios que conforman la cronosecuencia deberían diferir solamente en la edad (Fig. 15); esto es, que todos los sitios deberían compartir la misma historia de desarrollo y desarrollarse bajo las mismas circunstancias bióticas y abióticas (Johnson y Miyanishi, 2008). La violación a los supuestos implícitos en el uso de cronosecuencias limita su correcta aplicación y conducen a falsas conclusiones sobre los patrones ecológicos (Walker et al., 2010); sin embargo, la mayoría de los estudios han pasado por alto dicha restricción (Chazdon et al. 2007, Johnson y Miyanishi, 2008).
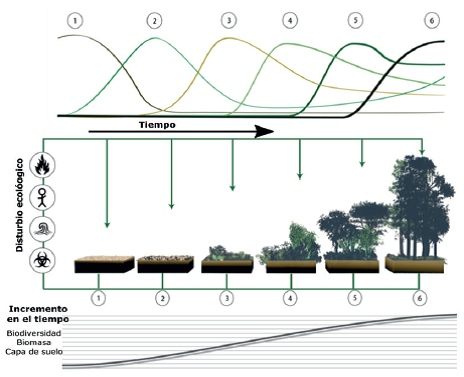
Fuente: adaptado de Luca Martin-Frey [CC BY 3.0]
Figura 15 Sucesión de la vegetación en seis etapas cronológicas: 1) roca intemperizada, 2) musgo y hierbas anuales, 3) pastos perennes, 4) arbustivas leñosas, 5) árboles de rápido crecimiento, 6) bosque clímax.
Si bien las cronosecuencias pueden llevar a plantear hipótesis sobre la forma de las trayectorias sucesionales y las tasas de cambio de los contenidos de C (Johnson y Miyanishi 2008, van Breugel, Martínez-Ramos y Bongers, 2006), no es posible asegurar que todas las sucesiones secundarias tengan una trayectoria similar, ya que en cada proceso de sucesión la estructura y composición varían con las condiciones de clima y suelo, intensidad y tiempo del uso anterior, cercanía a fuentes de semilla, capacidad de regeneración natural y desarrollo de las especies de interés económico. De ahí que existan áreas con alto potencial para la captura de C y áreas que solo permiten la recuperación parcial de la vegetación natural y algunas de sus funciones y servicios.
Por lo tanto, algunas de las trayectorias sobre la cantidad de C almacenado tenderán a un estado de vegetación cercano (Fig. 16, patrón A) similar al inicial (Fig. 16, patrón B) o distinto (Fig. 16, patrón C), en función del microclima, propiedades fisicoquímicas y biológicas del suelo, uso del suelo y prácticas de manejo prevalecientes en el lugar. El nuevo equilibrio en los contenidos de C puede ser similar, mayor o menor que el estado inicial de la vegetación, mientras que la permanencia temporal del C almacenado puede variar desde menos de un año hasta centurias (Ussiri y Lal, 2005).
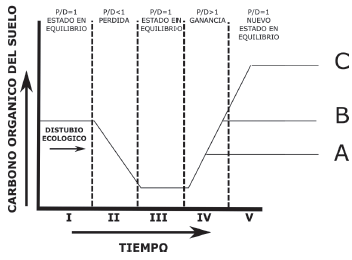
Figura 16 Modelo conceptual del contenido de C orgánico del suelo por producción y acumulación posterior a un disturbio o perturbación ecológica (Fuente: Adaptado de Johnson, Levine y Kern (1995) e IPCC (2005). Un estado en equilibrio (I) ocurre cuando las entradas de C vía producción de biomasa (P) es igual que las pérdidas de C vía descomposición (D), (i.e., P/D = 1). Después de una perturbación D frecuéntemente supera a P, lo que resulta en pérdida de C (II), hasta que un nuevo estado con bajo contenido de C se enriquece (III). Con la ejecución de buenas prácticas que favorecen la acumulación de C, donde P supera a D, ocurre re-acumulación (IV) de C hasta llegar a un nuevo estado enriquecido (V). Los nuevos estados (A, B, C) dependen de las prácticas adoptadas para recuperar carbono.
La revisión de contenidos de C en cronosecuencias para el compartimento de biomasa aérea mostrados en este trabajo solo atendió a 21 tipos de vegetación. Si se considera que la más reciente clasificación de los tipos de vegetación contenida en la cartografía de uso de suelo y vegetación digital escala 1:250 000 reconoce 50 tipos (Inegi, 2009), sin tomar en cuenta las condiciones de sucesión ecológica (vegetación primaria o secundaria) y pese a que es muy probable que existan investigaciones no detectadas en este trabajo, se sugiere realizar una investigación más exhaustiva de los documentos impresos de las instituciones dedicadas al estudio de la vegetación, ya que probablemente esos documentos no se encuentren digitalizados y disponibles electrónicamente. Para contar con una versión más aproximada de la capacidad de almacenamiento de C de los ecosistemas terrestres de México se requiere complementar la información de los contenidos de C en el almacén de la biomasa aérea viva, con los de biomasa subterránea (raíces), biomasa muerta en pie, biomasa muerta en piso, y mantillo y suelo.

