











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
El origen de la aceleración del cambio climático se ha atribuido a las actividades antropogénicas, especialmente al incremento en el uso combustibles fósiles (Intergovernmental Panel on Climate Change [IPCC], 2013); la emisión de gases de efecto invernadero (GEI) aumenta en promedio 1 Gt año-1 y las emisiones de CO2eq alcanzan casi 50 Gt año-1, esto ha permitido un aumento de temperatura de 0.05 °C en los últimos 15 años (IPCC, 2014). México se ubica entre los primeros diez países con emisiones altas de CO2, con 2% del total (Martínez y Fernández, 2004). La Organización de las Naciones Unidas para la Alimentación y la Agricultura [FAO] (2010) ha demostrado que los bosques, a través del procesos de la fotosíntesis, son los mayores reservorios de carbono del mundo almacenando en su biomasa 289 Gt y varios autores (Acosta, Vargas, Velásquez y Etchevers, 2002; Avendaño, Acosta, Carrillo y Etchevers, 2009; Aguirre y Jiménez, 2011) demuestran que los ecosistemas forestales son una alternativa biótica para almacenar carbono y de esa manera reducir la cantidad de CO2 atmosférico a través del proceso de la fotosíntesis, siendo esta una de las mejores formas de mitigar los efectos del cambio climático.
La mejor forma para estimar la biomasa vegetal es a través del método directo (Méndez, Luckie, Capó y Nájera, 2011), del cual se derivan ecuaciones alométricas (Chave et al., 2005; Návar, 2009). Para tener mejores estimaciones de biomasa es preferible probar diferentes ecuaciones (Fonseca, Alice y Rey, 2009), lo más común es usar combinaciones de variables dendrométricas fáciles de medir como diámetro normal y altura (Chave et al., 2005; Méndez et al., 2011); aunque otros autores (Návar, González, Graciano, Dale y Parresol, 2004; De los Ríos y Návar, 2010; Douterlungne, Herrera, Ferguson, Siddique y Soto, 2013) han demostrado que el diámetro basal del árbol es un buen estimador de biomasa y puede ser calculado directamente a partir del diámetro normal.
La importancia de contar con ecuaciones alométricas de biomasa es que a través de ellas es posible estimar de forma indirecta la cantidad de fijación de carbono en árboles individuales y extrapolar a cierta área y, con ello, evaluar la contribución de las especies en el amortiguamiento de los GEI, así como aportar información para fortalecer la política ambiental y gestión silvícola. Pinus halepensis Mill. es una especie endémica del Mediterráneo Occidental, se distribuye en regiones semiáridas y sub-húmedas, en altitudes que van de 0 m snm a 2600 m snm y con precipitación anual de 350 mm a 700 mm (Fady, Semerci y Vendramin, 2003). Por la característica que presenta de tener un rápido crecimiento, en 1929 fue introducida a México con motivos de restauración de ecosistemas (Cedeño y Pérez, 2007) y actualmente su uso se ha expandido en el centro y noreste de México. Por la importancia y gran uso de esta especie en el norte de México y, por otra parte, la escasez de ecuaciones de volumen y biomasa en esta especie, es necesario contar con ecuaciones alométricas para cuantificar almacenes de carbono.
Objetivos
Seleccionar la ecuación alométrica que mejor estime el volumen y la biomasa aérea en árboles individuales de P. halepensis en la sierra de Zapalinamé, Coahuila México.
Materiales y métodos
Área de estudio
Esta investigación se realizó en una plantación de P. halepensis de aproximadamente 40 años de edad, la cual fue establecida en varias etapas desde 1960 al 1978 en los terrenos de la Universidad Autónoma Agraria Antonio Narro, dentro de la sierra de Zapalinamé, al sureste de Saltillo Coahuila, México (25°20’41.75”N y 101°01’30.63”O; Datum WGS84). La plantación se estableció en hileras, siguiendo curvas de nivel con una distancia de aproximadamente 6 m entre árboles. La densidad fue 296 árboles ha-1 y es una de las plantaciones más grandes del país, con más de 1000 ha. El área se ubica a una altitud de 1800 m, posee una pendiente de 10%, exposición oeste y existen dos tipos de suelo (Litosol y Xerosol cálcico). La precipitación varía desde 250 mm hasta 400 mm, el clima es árido semicálido (Bsohw) y la temperatura oscila entre 18 °C y 22 °C (García, 1998).
Cuantificación de biomasa y volumen
El estudio se realizó con información de 40 árboles de P. halepensis que fueron seleccionados durante la ampliación de la carretera Saltillo-Zacatecas, en 2015. Antes de ser derribados, los árboles fueron medidos en diámetro a la base (Db), diámetro normal (Dn) diámetro de copa (Dc) y altura total (H). Después del derribo, cada árbol se troceó a longitud de no más de 150 cm, para separarlo en dos componentes: 1) hojas y ramillas (ramillas < 5 cm diámetro) y 2) fuste y ramas (ramas > 5 cm diámetro).
A cada componente se le determinó en campo el peso fresco total con báscula TecnoCour (México), con capacidad de 980.7 N (100 kg) y 4.9 N (0.5 kg) de resolución. El peso de muestras pequeñas (< 98.1 N; 10 kg) se obtuvo con una báscula Nuevo León (México) con capacidad de 980.7 N (100 kg) y 9.8 N (0.1 kg) de precisión. De cada árbol se obtuvieron cinco muestras de entre 19.6 N (2 kg; para hojas y ramillas) y 68.6 N (7 kg; para fuste y ramas), las cuales fueron pesadas en fresco para determinar el contenido de humedad. Las muestras completas se secaron en estufa Blue M. modelo 246F (U.S.A.), a temperatura de 70 °C hasta obtener peso constante. La biomasa se calculó a partir de la diferencia entre el contenido de humedad y peso fresco del componente (Picard, Saint- André y Henry, 2012) definiéndose como: biomasa de hojas y ramillas (Bhr), biomasa de fuste y ramas (Bfr) y por suma de ambas, biomasa total (Bt). El volumen (Vol) de cada troza del fuste y de ramas se calculó a partir de la fórmula de Smalian y, por suma, se obtuvo el volumen total del árbol en metros cúbicos.
Ajuste y selección de ecuaciones para cuantificar biomasa y volumen
Para cuantificar biomasa aérea y volumen de P. halepensis se probaron cinco ecuaciones alométricas (Tabla 1) propuestas por Picard, Saint-André y Henry (2012), obteniendo sus coeficientes de regresión por el método de mínimos cuadrados. La selección del mejor modelo estadístico se obtuvo acorde con Chave et al. (2005), usando el Criterio de Información Akaike (AIC, por sus siglas en inglés) y error estándar del modelo (Sxy). Además se consideraron otros estadísticos e indicadores de capacidad predictiva de los modelos como el Criterio de Información Bayesiano (BIC, por sus siglas en inglés), el coeficiente de determinación ajustado (R2 aj.) y el coeficiente de variación (CV), considerando la significancia estadística de los coeficientes de regresión (p ≤ 0.01) (Douterlungne et al., 2013; Cuenca, Jadán, Cueva y Aguirre, 2014). Para corregir el sesgo causado por la transformación logarítmica en la variable de respuesta, se obtuvo el factor de corrección ponderado (Sprugel, 1983). Los análisis estadísticos se realizaron usando el programa R (R Development Core Team, 2015).
Tabla 1 Ecuaciones alométricas probadas para cuantificar biomasa aérea y volumen de fuste y ramas en Pinus halepensis Mill. en la sierra de Zapalinamé, Coahuila, México.
| No. | Ecuación | Tipo de ajuste | Varianza |
| 1 | ln(y) = β0 + β1ln(Db) + ε | Lineal simple | Var(ε) = σ2 |
| 2 | ln(y) = β0 + β1ln(Db2H) + ε | Lineal simple | Var(ε) = σ2 |
| 3 | ln(y) = β0 + β1ln(Db) + β2ln(H) + ε | Múltiple | Var(ε) = σ2 |
| 4 | Y = β0 + β1Db2H + ε | Lineal ponderada | Var(ε) = ∝ Db2c |
| 5 | y / Db=β0/Db2 + β1H + ε | Lineal | Var(ε) = σ2 |
Donde y = biomasa (de cada componente en kg) y volumen (m3); Db = diámetro a la base (cm); H = altura total (m); β0, β1 y β2 = coeficientes de regresión; ln = logaritmo natural; ε = error; σ = varianza; ∝ y c = coeficientes de ponderación.
Resultados
De un total de 40 árboles, 67.5% mostraron bifurcación de fuste a menos de 1.3 m de altura, este alto porcentaje fue base para considerarlos como un solo árbol. El diámetro basal de los árboles promedió 46.86 cm ± 10.75 cm, 13.5 cm más que el promedio del diámetro normal. La altura de los árboles varió desde 7.12 m hasta 13.86 m (19%), pero el volumen desde 0.130 m3 hasta un máximo de 1.338 m3 (53%). La biomasa total (seca) mínima fue de 102.1 kg (Db = 26.9 cm) y la máxima de 1352.3 kg (Db = 75.4 cm) (Tabla 2). Es importante mencionar que se obtuvieron 3508 trozas, midiendo un total de 7016 diámetros, de los cuales 85% varió de 5 cm a 10 cm; el resto osciló entre 15 cm y 75 cm de diámetro.
Tabla 2 Estadísticas descriptivas de árboles usados para generar ecuaciones de biomasa y volumen de P. halepensis Mill., en la Sierra de Zapalinamé, Coahuila, México.
| Estadístico | Db | Dn ¶ | H | Bhr | Bfr | Bt | Vol |
| Promedio | 46.86 | 33.36 | 9.83 | 175.84 | 349.05 | 524.89 | 0.592 |
| Mínimo | 26.90 | 30.00 | 7.12 | 23.41 | 78.65 | 102.06 | 0.130 |
| Máximo | 75.45 | 39.50 | 13.86 | 487.67 | 864.70 | 1352.37 | 1.338 |
| Desv. Estandar | 10.75 | 3.10 | 1.86 | 84.70 | 183.07 | 257.61 | 0.313 |
| Varianza | 10.75 | 9.63 | 1.87 | 83.91 | 183.07 | 258.26 | 0.310 |
| Coef. de variación | 22.94 | 9.30 | 18.92 | 48.17 | 52.45 | 49.08 | 52.87 |
Donde: Db = diámetro a la base (cm); Dn = diámetro normal; H = altura (m); Bhr = biomasa en hojas y ramillas (kg); Bfr = biomasa en fuste y ramas (kg); Bt = biomasa total (kg); Vol = volumen (m3). ¶ = Dn de 13 árboles con bifurcación de fuste < 1.3 m de altura.
El porcentaje de biomasa de fuste y ramas de P. halepensis incrementó 1.2% por cada unidad de altura total; y como consecuencia, la biomasa de hojas y ramillas disminuyó 1.3% con la altura del árbol (Fig. 1A). La relación Bfr/Bhr indicó que en P. halepensis la biomasa de fuste y ramas fue en promedio 2.02 veces más que la biomasa de hojas (Fig. 1B). La densidad de la madera de P. halepensis (volumen de fuste y ramas/biomasa de fuste y ramas) fue en promedio 0.607 t/m3 (Fig. 1C) y fue menos variable en árboles con diámetro basal menor a 45 cm, en comparación con árboles con diámetro mayor.
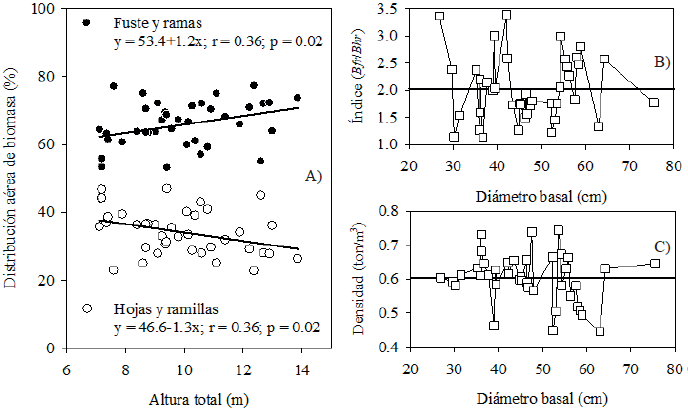
Figura 1 Distribución porcentual de biomasa aérea en función de altura total (A), relación Bfr/Bhr (B) y densidad de la madera de P. halepensis Mill. en función del diámetro basal (C).
Ecuaciones para la estimación de la biomasa y volumen
Para ahorrar espacio solo se muestran las ecuaciones de mejor ajuste (Tabla 3). La ecuación 2 (Tabla 1) es la expresión lineal del modelo alométrico y = e a x b y es una variación del modelo de potencia (y = ax b), usando “x” como una variable combinada diámetro y altura. Esta fue la que mejor predijo la biomasa de fuste y ramas, total y volumen, siendo en este orden de menor a mayor ajuste del modelo (R2 de 0.74 a 0.82).
Tabla 3 Coeficientes de regresión y estadísticos de ajuste de ecuaciones alométricas para estimar biomasa aérea y volumen de P. halepensis Mill. en la Sierra de Zapalinamé, Coahuila, México.
| No. | Modelo | R 2 aj. | S xy | CV | Valor de p | AIC | BIC | FC |
| 1 | Ln(Bhr)=Ln-2.5619(±1.48)+1.9910(±0.38)Ln(Db) | 0.70 | 46.49 | 26.70 | 1.27e -12 | 16.27 | 21.34 | 1.03 |
| 3 | Ln(Brf)=-2.6779(±1.49)+0.8463(±0.15)Ln(Db 2 H) | 0.74 | 92.81 | 26.59 | 9.02e -14 | 15.39 | 20.46 | 1.03 |
| 3 | Ln(Bt)=-1.8737(±1.34)+0.8077(±0.13)Ln(Db 2 H) | 0.77 | 124.74 | 23.84 | 1.36e -14 | 6.62 | 11.69 | 1.03 |
| 3 | Ln(Vol)=-9.7582(±1.33)+0.9161(±0.13)Ln(Db 2 H) | 0.82 | 0.13 | 22.63 | 2.36e -16 | 6.26 | 11.33 | 1.03 |
Donde: ln = logaritmo natural; Bhr = biomasa en hojas y ramillas (kg); Bfr = biomasa en fuste y ramas (kg); Bt = biomasa total (kg); Vol = volumen (m3); R2 aj = coeficiente de determinación ajustado; S xy = error estándar (kg); CV = coeficiente de variación (%); valor de p = significancia estadística; AIC = Criterio de Información Akaike; BIC = Criterio de Información Bayesiano; FC = factor de corrección; ± = intervalo de confianza de 95% del coeficiente de regresión.
Aunque los ajustes de los modelos de este estudio no fueron muy altos, en ningún caso se encontraron datos estadísticamente aberrantes. Todos los modelos fueron estadísticamente significativos (p < 0.0001); la variación más alta de los modelos se observó en biomasa de ramas (CV = 26.7%) y la menor en volumen (22.63%). El factor de corrección, además de corregir el sesgo, mejoró las estimaciones de biomasa de todos los componentes y de volumen en 3%, que ha sido el valor promedio obtenido en varios estudios similares.
La estimación de biomasa de hojas y ramillas, fuste y ramas, biomasa total y volumen fueron representados con el mejor modelo (Fig. 2A, 2B, 2C, 2D). Al comparar la precisión de la estimación de biomasa y volumen, se encontró que en general los datos observados versus datos estimados de los árboles, que midieron entre 34 cm y 60 cm de diámetro basal, se distribuyeron por encima y por debajo de la recta y = x, (relación 1:1). En árboles con diámetro basal fuera de estos límites, la ecuación sobreestimó y subestimó (área sombreada) respectivamente, tanto la biomasa como el volumen de P. halepensis (Fig. 2E, 2F, 2G, 2H). Las distancias verticales más grandes (errores en la predicción) corresponden básicamente a cuatro árboles cuyos diámetros basales fueron: 36.7 cm, 42.1 cm, 58.9 cm y 63 cm.

Figura 2 Biomasa estimada en: (A) hojas y ramillas, (B) fuste y ramas, (C) total; (D) volumen y (E-H) comparación de la capacidad predictiva del modelo en P. halepensis Mill., en la Sierra de Zapalinamé, Coahuila, México.
El error promedio porcentual más alto (32%) ocurrió en la biomasa de hojas y ramillas en la categoría de diámetro de 25 cm a 30 cm (Fig. 3A). En esa categoría el error promedio para la biomasa de fuste y total fue de 11.3% y 13.8%, respectivamente (Fig. 3B, 3C). Los errores más bajos se registraron entre las categorías de 30 cm y 60 cm de Db, promediando 2.1% para biomasa de hojas y ramillas (Fig. 3A), 3.0% para fuste y ramas (Fig. 3B) y 0.9% para la biomasa total (Fig. 3C). La ecuación 2 (Tabla 1) fue la que mejor estimó biomasa y se ajustó mejor al volumen de P. halepensis (Tabla 3). Su error porcentual en las diferentes categorías de diámetro basal fue homogéneo (Fig. 3D), promediando 4.7% (línea blanca del gráfico).
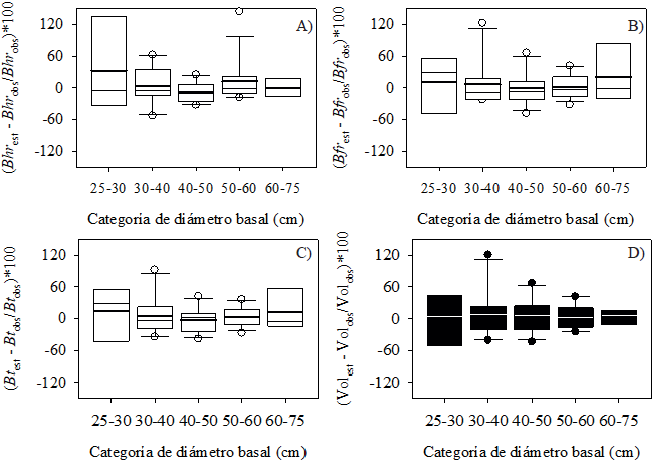
Figura 3 Error porcentual en biomasa de hojas y ramillas (A), de fuste y ramas (B), total (C) y en volumen (D) en P. halepensis Mill., en la sierra de Zapalinamé, Coahuila, México.
El análisis de una muestra de 122 árboles de P. halepensis procedentes de la misma área de estudio mostró alta correlación (r = 0.97) entre Dn y Db (Fig.4A) y paralelismo (misma pendiente, b = 1.00) según la ecuación y = x. Dicha ecuación se desplaza hacia un sentido positivo solamente 7.31 unidades de Db con respecto a la relación 1:1. Es decir, para uso de las ecuaciones de este estudio y especie, la transformación de Dn a Db puede hacerse eficientemente por medio de la ecuación señalada (Fig. 4A). La altura total de P. halepensis también puede ser estimada eficientemente mediante un modelo lineal (Fig. 4B). La relación Db/H (Fig. 4C) y Db/Dn (Fig. 4D) siguieron una tendencia logarítmica negativa.

Discusión
En P. halepensis la mayor cantidad de biomasa aérea se distribuyó en el fuste y ramas (64%) con respecto a biomasa total, el resto (hojas y ramillas) representó en promedio 36%.
Este patrón fue similar al encontrado para P. devoniana y P. pseudostrobus Lind., con 60% y 58% en fuste y 40% y 42% en hojas y ramas (Méndez et al., 2011). En Pinus montezumae Lamb., el mayor porcentaje de biomasa se presentó en fuste (77%) y en ramas y hojas promedió 9% (Carrillo, Acosta, Flores, Juárez y Bonilla, 2014). Por su parte Rodríguez et al. (2012) registraron que la distribución de biomasa en P. patula Schl. et Cham. fue 92.9%, 4.7% y 2.4% para fuste, ramas y follaje, respectivamente.
Al estimar la biomasa de P. halepensis, usando diámetros normales de 25 cm, 35 cm y 45 cm, y compararla con la biomasa aérea de otras especies, se encontró que en general la mayoría de las especies almacenaron más biomasa que P. halepensis (Fig. 5A). P. arizonica Engelm., para esos mismos diámetros normales, almacenó 2.0, 2.2 y 2.4 veces más, representando en promedio 130 326 kg y 644 kg de biomasa y hasta de 3.2 veces más en P. montezumae Lamb. para el diámetro de 45 cm (Carrillo et al., 2014). La magnitud del error (Sxy) de estimación de biomasa no fue totalmente dependiente de la dimensión de la variable “y” (Fig. 5B), sino que dependió del tipo (simple, múltiple, etc.) y del número de variables independientes consideradas en el modelo. Los ajustes de los modelos (R2) encontrados en la literatura (Fig. 5C) son altos (R2 > 0.91) y mayores a los de P. halepensis y no son proporcionales al tamaño de muestra (Fig. 5D).

Figura 5 Comparación de la biomasa aérea (seca y fresca) de P. halepensis Mill. con la biomasa seca de diferentes especies (A), estadísticos: errorde estimación (B), ajuste (C) y número de observaciones (D) de ecuaciones usadas para estimar biomasa aérea de las especies del lado izquierdo.
En árboles de Abies religiosa (Kunth) Schltdl. & Cham. la biomasa en fuste fue casi 85% del total, mientras que en ramas y hojas fue 7% y 8%, respectivamente (Avendaño et al., 2009). A pesar de que las hojas representan la menor cantidad de biomasa con respecto al total en casi todas las especies, algunos autores (Rodríguez et al., 2012) sugieren que esta debe ser estimada con precisión, especialmente por su relación con la fotosíntesis y productividad primaria neta. Específicamente para P. halepensis, López et al. (2013) demostraron que las hojas fueron el componente más activo, cuyas variaciones dependieron de la precipitación y temperatura y de los sitios en donde ocurre el mayor flujo de carbono de la vegetación al suelo.
En este, como en varios estudios, se ha demostrado que la biomasa total es el componente mejor estimado (Návar, 2009; Fonseca et al., 2009; Méndez et al., 2011; Cuenca et al., 2014). En contraste, la biomasa de ramas presenta la mayor variación y es en este componente en donde ocurre el menor ajuste, independientemente de la especie y el género (Aguirre y Jiménez, 2011).
La ecuación con la variable combinada (diámetroaltura) que mejor predijo biomasa de P. halepensis en este estudio se ha utilizado también para estimar el volumen de esta especie (Návar y Domínguez, 1997) y en la mayoría de otras especies (Muñoz et al., 2012). La mayoría de las ecuaciones alométricas se han desarrollado con la variable diámetro normal (Picard et al., 2012; Cuenca et al., 2014), aunque por las características de la especie y lo práctico de medir algunas variables de árbol, varios autores han usado diámetro basal (Návar et al., 2004; De los Ríos y Návar, 2010; Douterlungne et al., 2013).
Cabe destacar que el peso seco de especies registrado por Brown (1997), Návar (2009) y Carrillo et al. (2014) (Fig. 4A) es aún mayor que el peso verde de P. halepensis de este estudio, variando desde 1.2 (P. arizonica) hasta 1.8 veces más (P. montezumae), pero en promedio 1.2 veces más que P. halepensis. El peso verde de P. halepensis registrado aquí es similar al peso seco de P. pseudostrobus registrado por Aguirre y Jiménez (2011). Diferencias de almacenes de biomasa específicas e interespecíficas han sido documentadas por Návar (2009) y Chave, Riéra y Dubois (2001), demostrándose que estas son dependientes de las condiciones de crecimiento (Méndez et al., 2011) y de la calidad de sitio (Návar, 2009), de la misma especie y de la densidad del rodal (Aguirre y Jiménez, 2011).
El número de árboles con que se han desarrollado las ecuaciones de biomasa ha sido variable, desde 15 (Carrillo et al., 2014) hasta 384 (Návar, 2009) (Fig. 4B). Particularmente en México, más de 50% de los estudios de ecuaciones alométricas para cuantificar biomasa se han construido usando desde 10 hasta 40 árboles.
Algunos autores, para mejorar la estimación de biomasa, han sugerido incluir en el modelo la densidad de la madera (Chave et al., 2005; Picard et al., 2012) o usar modelos múltiples y polinomiales (Chave et al., 2005) con densidad, área del tronco, diámetro y altura. El valor de densidad básica de la madera de P. halepensis de este estudio (0.607 t/m3) es muy similar al obtenido por Centre de la Propietat Forestal (2004) para esta misma especie (0.610 t/m3).
Conclusiones
La mejor predicción de biomasa aérea y volumen de P. halepensis se logró con la ecuación que incluyó la variable combinada diámetro y altura en forma logarítmica, mientras que el diámetro por sí solo mostró estimaciones sesgadas. Los errores más altos (subestimación) de biomasa aérea de P. halepensis ocurrieron en las categorías de diámetro inferiores, lo cual se debió a la alta variación del diámetro-altura en edades tempranas. La biomasa aérea de P. halepensis fue hasta 2.5 veces menor que la de otras especies de coníferas que no se ramifican desde la base del árbol. Ochenta y cinco por ciento de las ramas de P. halepensis variaron entre 5 cm y 10 cm de diámetro, contrario a lo que se ha registrado para la mayoría de las coníferas. La particularidad de la especie se reflejó también en menor ajuste de los modelos alométricos en comparación con otras especies de coníferas.

