











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
La vegetación que se desarrolla debajo del dosel arbóreo, comúnmente llamada sotobosque, tiene entre sus principales elementos estructurales a las briofitas, los líquenes, los helechos, las plántulas arbóreas, las herbáceas y los arbustos (Antos, 2017). En el bosque templado, el sotobosque cuenta con una importante diversidad de especies vegetales y juega un rol importante en la dinámica del ecosistema (Bartels y Chen, 2010).
El sotobosque es esencial para procesos del ciclo de nutrientes, la trayectoria sucesional o el régimen de fuego es el hábitat o fuente de alimento para numerosos vertebrados e invertebrados (Gilliam, 2007). En este, algunas especies crean condiciones de interferencia en las plántulas arbóreas que varían en competencia, alelopatía, inhibición micorrícica y obstrucción física (Royo y Carson, 2006).
El hábitat en el sotobosque es heterogéneo y dinámico, con un ambiente variable que depende de las características del dosel y de los recursos en el medio (Augusto, Dupouey y Ranger, 2003; Hart y Chen, 2006). En los bosques de coníferas, el sotobosque presenta regularmente una elevada humedad, con poca luz, viento y escasos nutrientes en el suelo. En dichas comunidades la tolerancia hacia los suelos ácidos es amplia (Candan, Broquen y Pellegrini, 2006; Rzedowski, 2006).
Con respecto a la composición florística, el sotobosque de las coníferas presenta una amplia riqueza de especies. En los bosques de Pinus hartwegii, los componentes dominantes son las gramíneas de géneros como Festuca, Calamagrostis y Muhlenbergia. Eventualmente se encuentran especies del género Lupinus como resultado de la ocurrencia de incendios (Rzedowski, 2006). En el bosque de Abies religiosa, por el contrario, el sotobosque es poco abundante en gramíneas. En ambos bosques, los representantes de la familia Asteraceae son numerosos (Encina-Domínguez, Encina-Domínguez, Mata-Rocha y Valdes-Reyna, 2008).
En el área de protección de flora y fauna Nevado de Toluca (Apffnt), los bosques de pino y oyamel ocupan 66% de la superficie forestal (Endara, 2010), donde 7815 ha son bosques densos de pino y 13 040 ha bosques densos de oyamel (Franco, Regil, González y Nava, 2006). En la estructura espacial los bosques densos de Pinus hartwegii y Abies religiosa presentan tres estratos verticales que llegan a los 30 m y 48 m respectivamente. La densidad promedio de individuos en el bosque de pino es de 336 ha-1 y de 637 ha-1 (≥ 2.5 cm DAP) en el bosque de oyamel (Endara, Franco, Nava, Valdez y Fredericksen, 2012).
Objetivos
En el Nevado de Toluca existen extensas regiones dominadas por especies arbóreas nativas, que definen la fisonomía de la comunidad (Mastretta et al., 2014), por ello es necesario caracterizar su composición vegetal y promover la conservación de sus especies. En el presente trabajo se planteó, como objetivo central, determinar la composición y estructura de herbáceas y arbustos en áreas de bosque denso de Pinus hartwegii y Abies religiosa del Nevado de Toluca. Adicionalmente se cuantificó la densidad de la regeneración arbórea.
Materiales y métodos
El Apffnt se localiza en el Estado de México, hacia el suroeste del valle de Toluca y abarca parte de los municipios de Almoloya de Juárez, Amanalco de Becerra, Calimaya, Coatepec Harinas, Temascaltepec, Tenango del Valle, Toluca, Villa Guerrero, Villa Victoria y Zinacantepec (Gobierno del Estado de México [GEM], 1999). Forma parte del sistema volcánico transmexicano y alcanza una altitud de 4660 m snm. En el área predomina el tipo de suelo Andosol y en menor proporción Feozem, Regosol, Cambisol y Litosol. De los 3700 m snm en adelante predomina el clima frío y, entre los 2800 m y los 3700 m snm, el clima semifrío subhúmedo con lluvias en verano. En el Apffnt la riqueza de coníferas comprende especies de pino: Pinus hartwegii Lindl., P. montezumae Lamb., P. pseudostrobus Lindl., entre otros, así como oyamel (Abies religiosa Kunth Schltdl. & Cham.), Juniperus flaccida Schlechtendal y Cupressus lusitánica Mill. Además, se desarrollan bosques de Quercus laurina Humb et Bonpl. y Alnus jorullensis Humboldt, Bonpland & Kunth, así como zonas de pastizal, zacatonal alpino o páramo de altura (Comisión Nacional de Áreas Naturales Protegidas, 2013). El muestreo de la vegetación se realizó en los meses de agosto a octubre de 2015, en las localidades de Agua Blanca y Santa María del Monte (Zinacantepec) (Fig. 1). Las zonas de bosque denso de Abies religiosa y Pinus hartwegii se diferenciaron previamente con la ayuda de imágenes de satélite multiespectrales. En el área, se considera bosque denso a aquella superficie que cubre entre 80% y 100% de cobertura de copa, identificado sobre un cuadrante cartografiable de 100 m2 (Regil, Franco, Nava y Ordóñez, 2015).

En cada sitio de muestreo se establecieron 10 cuadrantes de 10 m ×10 m (1000 m2) en un gradiente continuo y en cada uno se registraron altitud, pendiente, coordenadas geográficas y tipo de perturbación asociada. Para confirmar la representatividad del muestreo, se realizó una curva de acumulación de especies en cada comunidad vegetal (Fig. 2) y, para analizar la composición, se realizó un método discriminativo por altura; en las especies mayores a 50 cm, se obtuvieron los valores relativos en cobertura, densidad, frecuencia y valor de importancia (Mostacedo y Fredericksen, 2000). De las especies menores a 50 cm se estimó la cobertura y abundancia con base en la escala Braun-Blanquet (Braun-Blanquet, 1979).
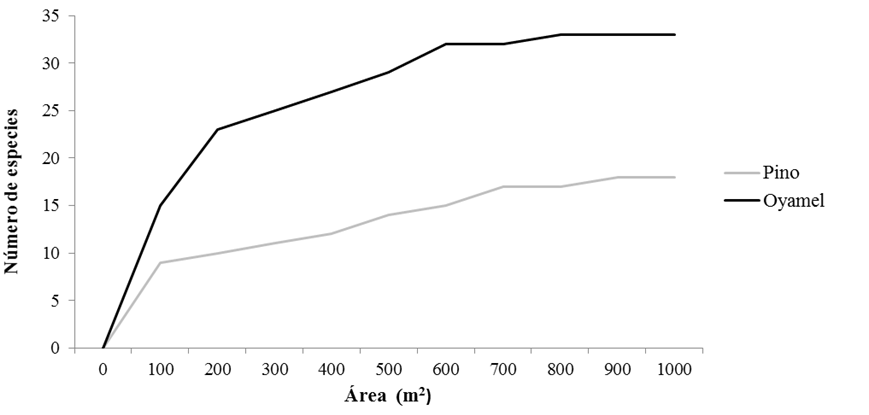
Por la elevada densidad de gramíneas (mayores a 50 cm) en el bosque de pino, el muestreo se estandarizó al método Braun-Blanquet. Finalmente, para evaluar la densidad de regeneración arbórea, se cuantificó el número de plántulas (< 30 cm de altura), brinzales (≥ 30 cm < 1.5 m altura) y latizales (≥ 1.5 m < 2.5 cm de DAP) (Valdez, 2002).
De cada especie vegetal registrada, se recolectaron ejemplares por triplicado para su posterior herborización y determinación taxonómica con claves especializadas. En el Herbario-Hortorio (Chapa) del Campus Montecillo del Colegio de Postgraduados, se depositaron duplicados de los ejemplares recolectados y se confirmó la determinación de especímenes. La agrupación de familias se realizó de acuerdo con el sistema Angiosperm Phylogeny Group[APG IV] (2016) y para las abreviaturas de los autores de las especies se utilizó el Índice Internacional de Nombres de Plantas (The International Plant Names Index [IPNI], 2017). El nombre común de las especies, se obtuvo por consulta a las personas de las localidades adyacentes a las zonas de muestreo y se complementó con bibliografía especializada (Rzedowski, Rzedowski y colaboradores, 2005; Cornejo-Tenorio e Ibarra-Manríquez, 2008). La agrupación de especies en gremios ecológicos se realizó de forma preliminar con base en las características ecológicas y foliares de la especie (Givnish, 1988; Grime, 1989).
Resultados
Composición florística
En el bosque de Abies religiosa, la flora del sotobosque cuenta con un total de 33 especies, 25 géneros y 14 familias agrupadas en 24 herbáceas, ocho arbustos y un árbol (Tabla 1).
Tabla 1 Flora vascular del sotobosque en el bosque denso de Abies religiosa del Nevado de Toluca.
| FAMILIA/ Nombre científico | Nombre común | Forma de vida | Gremio ecológico |
|---|---|---|---|
| APIACEAE | |||
| Osmorhiza aff. mexicana Griseb. | S/D | Herbácea | Heliófita |
| ASTERACEAE | |||
| Ageratina aff. parayana 2 (Espinosa) B. L. Turner | Raíz de serpiente | Herbácea | Esciófita |
| Ageratina pazcuarensis (Kunth) R. M. King & H. Rob. | Hierba del ángel | Herbácea | Esciófita |
| Ageratina rivalis (Greenm.) R. M. King & H. Rob. | S/D | Arbusto | Heliófita |
| Bidens triplinervia 1 Kunth | Acahual cimarrón | Herbácea | Heliófita |
| Cirsium ehrenbergii 12 Sch. Bip. | Cardo santo | Herbácea | Esciófita |
| Piqueria pilosa 2 Kunth | Cardón pelón | Herbácea | Esciófita |
| Roldana angulifolia (DC.) H. Rob. & Brettell | Hoja ancha | Arbusto | Esciófita |
| Roldana lineolata 12 (DC.) H. Rob. & Brettell | S/D | Herbácea | Esciófita |
| Roldana platanifolia 2 (Benth.) H. Rob. & Brettell | Hierba del zopilote, mano de león | Herbácea | Esciófita |
| Roldana reticulata 2 (DC.) H. Rob. & Brettell | S/D | Herbácea | Esciófita |
| Senecio callosus Sch. Bip. | Hoja de flecha | Herbácea | Esciófita |
| Senecio cinerarioides 12 Kunth | Jara, Jarilla blanca | Arbusto | Heliófita |
| Sigesbeckia jorullensis 1 Kunth | Flor de araña | Herbácea | Esciófita |
| Stevia monardifolia 2 Kunth | S/D | Herbácea | Esciófita |
| CAPRIFOLIACEAE | |||
| Symphoricarpos microphyllus Kunth | Perlilla, Perlitas, Escobilla | Arbusto | Heliófita |
| CARYOPHYLLACEAE | |||
| Cerastium nutans Raf. | S/D | Herbácea | Esciófita |
| Stellaria cuspidata Willd. | S/D | Herbácea | Esciófita |
| DRYOPTERIDACEAE | |||
| Plecosorus speciosissimus (A. Braun ex Kunze) T. Moore | S/D | Herbácea | Esciófita |
| GERANIACEAE | |||
| Geranium seemannii Peyr. | Mano de león | Herbácea | Esciófita |
| LAMIACEAE | |||
| Salvia gracilis Benth. | S/D | Herbácea | Esciófita |
| Salvia lavanduloides Kunth | Tepechia | Herbácea | Esciófita |
| Salvia aff. microphylla Kunth | Bandera mexicana | Arbusto | Esciófita |
| ONAGRACEAE | |||
| Fuchsia microphylla Kunth | Aretillo | Arbusto | Esciófita |
| PINACEAE | |||
| Abies religiosa (Kunth) Schltdl. & Cham. | Oyamel, abeto | Árbol | Esciófita |
| POACEAE | |||
| Deschampsia sp. | S/D | Herbácea | Esciófita |
| Trisetum irazuense Hitchc. | S/D | Herbácea | Esciófita |
| ROSACEAE | |||
| Acaena elongata 1 L. | Pegarropa, cadillo | Arbusto | Esciófita |
| Alchemilla procumbens 1 Rose | Chinilla, pata de león | Herbácea | Esciófita |
| RUBIACEAE | |||
| Didymaea alsinoides (Schltdl. & Cham.) Standl. | Ocoxóchitl | Herbácea | Esciófita |
| Galium aschenbornii S. Schauer | Cuajaleche, pegarropa | Herbácea | Esciófita |
| SCROPHULARIACEAE | |||
| Sibthorpia repens (L.) Kuntze | Pepita | Herbácea | Esciófita |
| SOLANACEAE | |||
| Cestrum anagyris Dunal | Hierba del zopilote | Arbusto | Esciófita |
“1” especies asociadas a disturbio, “2” especies endémicas del país, “S/D” sin dato.
El bosque de Pinus hartwegii se caracteriza por un sotobosque con 18 especies, 16 géneros y nueve familias, que se agrupan en 15 herbáceas, dos arbustos y un árbol (Tabla 2).
Tabla 2 Flora vascular del sotobosque en el bosque denso de Pinus hartwegii del Nevado de Toluca.
| FAMILIA/ Nombre científico | Nombre común | Forma de vida | Gremio ecológico |
|---|---|---|---|
| ASTERACEAE | |||
| Cirsium jorullense 2 Spreng. | Cardo santo | Herbácea | Esciófita |
| Pseudognaphalium liebmannii (Sch. Bip. ex Klatt) | Gordolobo, hierbabuena, altamisa | Herbácea | Esciófita |
| Senecio cinerarioides 12 Kunth | Jara, Jarilla blanca | Arbusto | Heliófita |
| Senecio iodanthus 2 Greenm. | S/D | Herbácea | Esciófita |
| Senecio roseus 2 Sch. Bip. | Sierrilla | Herbácea | Esciófita |
| Stevia incognita Grashoff | Yoloquilde | Herbácea | Esciófita |
| CARYOPHYLLACEAE | |||
| Arenaria bryoides D. F. K. Schltdl. | S/D | Herbácea | Esciófita |
| ERICACEAE | |||
| Vaccinium caespitosum Michx. | Arándanos, Borrachos | Arbusto | Esciófita |
| FABACEAE | |||
| Lupinus montanus 1 Kunth | Garbancillo, matapiojo | Herbácea | Heliófita |
| OXALIDACEAE | |||
| Oxalis alpina Rose | Xocoyotl | Herbácea | Esciófita |
| PINACEAE | |||
| Pinus hartwegii Lindl. | Ocote | Árbol | Heliófita |
| POACEAE | |||
| Agrostis tolucensis Kunth | S/D | Herbácea | Heliófita |
| Calamagrostis tolucensis (Kunth) Trin. ex Steud. | Zacatón | Herbácea | Heliófita |
| Cinna poiformis (Kunth) Scribn. & Merr. | S/D | Herbácea | Heliófita |
| Festuca tolucensis Kunth | Zacatón | Herbácea | Heliófita |
| Trisetum spicatum (L.) K. Richt. | S/D | Herbácea | Heliófita |
| ROSACEAE | |||
| Alchemilla procumbens 1 Rose | Chinilla, pata de león | Herbácea | Esciófita |
| SCROPHULARIACEAE | |||
| Penstemon gentianoides Poir. | Jarritos, campanita morada | Herbácea | Esciófita |
“1” especies asociadas a disturbio, “2” especies endémicas del país, “S/D” sin dato.
En cuanto a la regeneración arbórea de las áreas evaluadas se encontró que esta es exclusiva de las especies de oyamel y pino, las plántulas fueron dominantes en densidad, con 87.2% en Abies y 47% en Pinus, seguidas por brinzales (12.2% Abies y 41.2% Pinus) y latizales (0.6% Abies y 11.8% Pinus).
Estructura del sotobosque
En el sotobosque de Abies religiosa, las herbáceas: Stevia monardifolia, Salvia gracilis y Sigesbeckia jorullensis son dominantes en la altura mayor a 50 cm, en este nivel también predominan los arbustos Roldana angulifolia y Cestrum anagyris. En las herbáceas menores a 50 cm, Stevia monardifolia, Roldana platanifolia y Sibthorpia repens tienen la mayor cobertura y abundancia, mientras que los arbustos Roldana angulifolia, Acaena elongata y Fuchsia microphylla son dominantes (Tabla 3 y Fig. 3).
Tabla 3 Especies dominantes del sotobosque en el bosque denso de Abies religiosa*.
| Estrato ≥ 50 cm | |||||||||||
| Valor de importancia | |||||||||||
| Densidad relativa | Cobertura relativa | Frecuencia relativa | VIR | ||||||||
| Herbáceas | |||||||||||
| Stevia monardifolia | 34.4 | 46.6 | 33.3 | 38.1 | |||||||
| Salvia gracilis | 39.1 | 20.9 | 8.3 | 22.8 | |||||||
| Sigesbeckia jorullensis | 17.2 | 17.7 | 25 | 20 | |||||||
| Arbustos | |||||||||||
| Roldana angulifolia | 94.6 | 93.1 | 40 | 75.9 | |||||||
| Cestrum anagyris | 4.6 | 6.3 | 40 | 17 | |||||||
| Estrato ≤ 50 cm | |||||||||||
| Escala Braun-Blanquet | |||||||||||
| Cuadrante | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |
| Herbáceas | |||||||||||
| Stevia monardifolia | 2 | 4 | 2 | 4 | 3 | 2 | 3 | 3 | 3 | 2 | |
| Roldana platanifolia | 3 | 3 | 4 | 5 | 3 | 2 | 2 | 2 | 2 | + | |
| Sibthorpia repens | 3 | 2 | 1 | 3 | + | 3 | 3 | 3 | 2 | 1 | |
| Ageratina pazcuarensis | - | + | - | 3 | 3 | 2 | 4 | 3 | 2 | 3 | |
| Ageratina aff. parayana | + | - | - | 2 | + | + | 3 | + | 4 | 4 | |
| Sigesbeckia jorullensis | + | - | - | 2 | + | 2 | + | 3 | 2 | + | |
| Trisetum irazuense | + | 1 | + | + | + | 1 | + | + | + | + | |
| Piqueria pilosa | + | - | - | - | 2 | + | 2 | 2 | 1 | - | |
| Arbustos | |||||||||||
| Roldana angulifolia | 4 | 4 | 3 | 3 | 2 | 2 | 4 | 4 | 4 | 2 | |
| Acaena elongata | - | 1 | 1 | 1 | 2 | 3 | 3 | 2 | 3 | + | |
| Fuchsia microphylla | 2 | 2 | 2 | 3 | 1 | 1 | + | 1 | - | + | |
| Cestrum anagyris | - | 2 | 2 | + | + | 2 | + | 2 | + | 2 | |
* Se indica el valor de importancia o la estimación de cobertura y abundancia.
Escala Braun-Blanquet: cobertura de la especie; (-) ausencia, (+) pocos individuos con baja cobertura, (1) <5%, (2) 5 - 25%, (3) 25 - 50%, (4) 50 - 75%. Los resultados señalados en la tabla consideran a las especies dominantes, las especies restantes se omitieron por ser poco informativas.
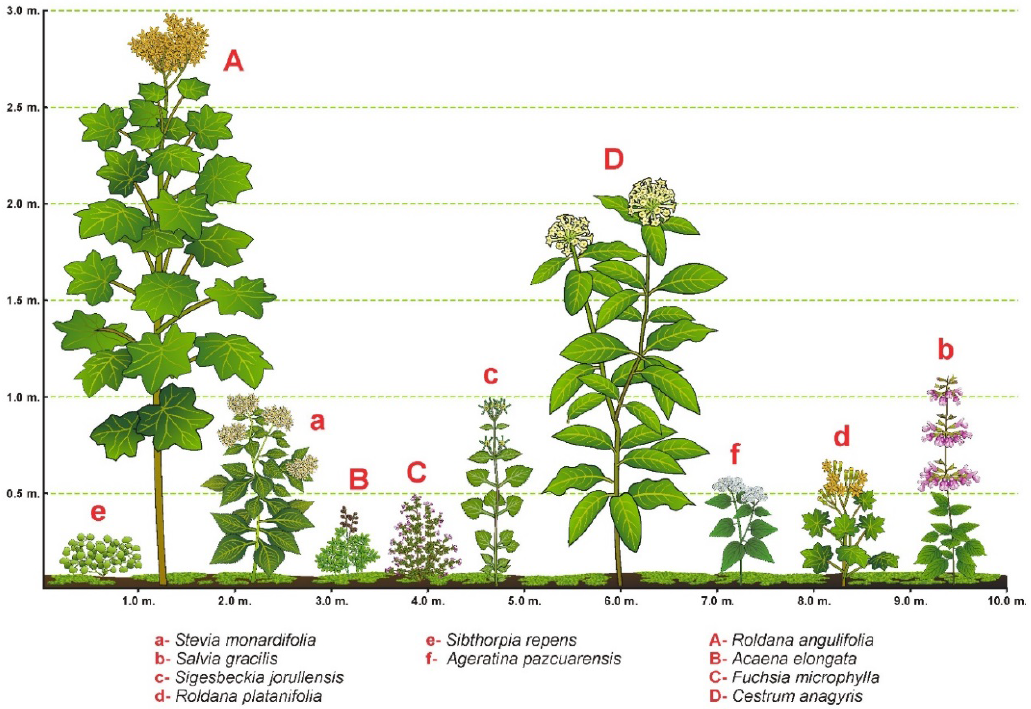
Figura 3 Estructura del sotobosque en el bosque de Abies religiosa, se presenta la altura y cobertura aproximada de las especies dominantes.
Con respecto al sotobosque en el bosque denso de Pinus hartwegii estudiado, las herbáceas dominantes en la altura mayor a 50 cm son Calamagrostis tolucensis, Agrostis tolucensis, Penstemon gentianoides y Lupinus montanus, en esta categoría, la única especie arbustiva dominante es Senecio cinerarioides. En el nivel menor a 50 cm Alchemilla procumbens y Penstemon gentianoides son las herbáceas dominantes (Tabla 4 y Fig. 4).
Tabla 4 Especies dominantes del sotobosque en el bosque denso de Pinus hartwegii*.
| Estrato ≥ 50 cm | |||||||||||
| Valor de importancia | |||||||||||
| Densidad relativa | Cobertura relativa | Frecuencia relativa | VIR | ||||||||
| Herbáceas | |||||||||||
| Penstemon gentianoides | 61.3 | 60 | 42.9 | 54.7 | |||||||
| Lupinus montanus | 35.2 | 37.7 | 47.6 | 40.2 | |||||||
| Arbustos | |||||||||||
| Senecio cinerarioides | 100 | 100 | 100 | 100 | |||||||
| Estrato ≤ 50 cm | |||||||||||
| Escala Braun-Blanquet | |||||||||||
| Herbáceas | Cuadrante | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
| Calamagrostis tolucensis | 4 | 4 | 3 | 4 | 5 | 4 | 5 | 4 | 5 | 4 | |
| Alchemilla procumbens | 3 | 3 | 2 | 2 | 3 | 3 | 2 | 2 | 2 | 2 | |
| Penstemon gentianoides | 2 | 3 | 2 | 2 | 2 | 2 | 3 | 2 | 2 | 2 | |
| Agrostis tolucensis | 3 | 1 | 4 | 3 | 2 | 2 | 1 | 2 | 2 | + | |
| Lupinus montanus | + | + | + | + | + | + | + | 2 | + | 2 | |
| Oxalis alpina | - | + | + | + | + | + | + | + | + | + | |
* Se indica el valor de importancia o la estimación de cobertura y abundancia.
“0” especies mayores a 50 cm. Escala Braun-Blanquet: cobertura de la especie; (-) ausencia, (+) pocos individuos con baja cobertura, (1) <5%, (2) 5 - 25%, (3) 25 - 50%, (4) 50 - 75%, (5) 75% - 100%. Los resultados señalados en la tabla consideran a las especies dominantes, las especies restantes se omitieron por ser poco informativas.
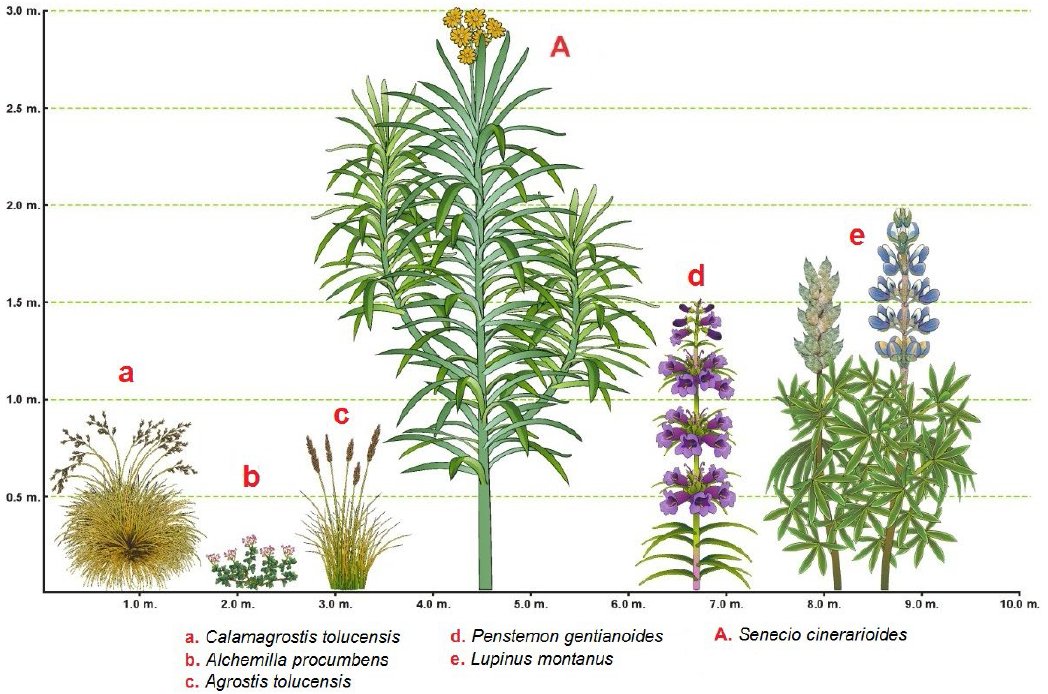
Discusión
Composición florística
La vegetación del sotobosque en la comunidad de Abies religiosa posee una mayor riqueza de especies que la comunidad de Pinus hartwegii. Este dato concuerda con el registro de Villers, García y López (1998) en el Nevado de Toluca y con lo registrado por Vázquez (2014) en el Cofre de Perote, donde los bosques de Abies religiosa son más diversos y preceden en distribución altitudinal a los bosques de Pinus hartwegii.
La riqueza florística del bosque denso de Abies religiosa en el Nevado de Toluca es menor a la encontrada por Ávila-Akerberg (2010) en la cuenca del río Magdalena (46 especies: 26 herbáceas, 15 arbustos y cinco árboles), pero mayor a la registrada por Vázquez (2014) en Veracruz con 27 especies (18 herbáceas, siete arbustos y dos árboles). Es posible que la riqueza de especies en el sotobosque de la especie esté determinada por múltiples factores, entre ellos la pendiente o la orientación (Gracia, Montané, Piqué y Retana, 2007; Huo, Feng y Su, 2014). Se trata de comunidades que se distribuyen principalmente en barrancas, cañadas o laderas, donde la pendiente llega a superar 40% y la humedad es elevada (Sánchez-González y López-Mata, 2003).
La riqueza de especies en el bosque denso de Pinus hartwegii en el Nevado de Toluca es menor al registro de Ávila-Akerberg (2010) en la cuenca del río Magdalena (47 especies) y a lo encontrado por Vázquez (2014) en el Cofre de Perote con 21 especies. En la cuenca del río Magdalena la flora cuenta con 42 herbáceas, cuatro arbustos y un árbol, mientras que en el Cofre de Perote existen 17 herbáceas, tres arbustos y un árbol. En los bosques de esta especie la riqueza florística suele estar condicionada a su distribución altitudinal. Hacia el límite superior de la vegetación arbórea, factores como la temperatura, la precipitación, la radiación solar y los nutrientes del suelo reducen el crecimiento de las plantas (Sharma, Suyal, Gairola y Ghildiyal, 2009). Además de estos factores, otros como el nivel de adaptación o tolerancia al ambiente de las especies, el tipo o estructura del dosel en el que se desarrollan y la disponibilidad de recursos, determinan la composición y estructura del sotobosque (North, Oakley, Fiegener, Gray y Barbour, 2005; Barbier, Gosselin y Balandier, 2008; Fuxai, Fousseni, Chungang, Huaijiang y Xiuhai, 2014).
En el sotobosque de ambas comunidades evaluadas la forma de vida que predomina es la herbácea, seguida de los arbustos y los árboles, resultado que avala el registro de Villaseñor y Ortiz (2014) para las plantas con flores del país. Así mismo la familia que reúne el mayor número de especies y endemismos es Asteraceae. En México, dicha familia representa la mayor riqueza florística y un elevado porcentaje de endemismos (Villaseñor, 2003; Villaseñor, Maeda, Colín y Ortiz, 2005). En el sotobosque de Abies religiosa, el género más representativo fue Roldana y en el sotobosque de Pinus hartwegii fue Senecio.
Además de la familia Asteraceae, en el sotobosque de Pinus hartwegii la familia Poaceae presenta un nivel de importancia significativo. En el ambiente la abundancia de gramíneas está asociada frecuentemente a la rápida adaptación o expansión que presentan y que les permite aprovechar los recursos del medio. Estas especies crecen en condiciones de deficiencia de drenaje, inundaciones periódicas o suelos con exceso de sales solubles, lo que las convierte en especies competidoras (Grime, 1989; Rzedowski, 2006). Es importante señalar que en ambos ecosistemas la mayor parte de las especies encontradas son de origen nativo, situación prevaleciente en gran parte del territorio nacional (Villaseñor, 2016).
Las especies tolerantes a la sombra (esciófitas) dominan la composición vegetal de ambas comunidades evaluadas, en el sotobosque de oyamel esta dominancia es justificable por la densidad de individuos que presenta el dosel (637 ha-1). Esta densidad es significativamente menor en el bosque de pino (336 ha-1) (Endara et al., 2012). En el bosque de oyamel, la estructura arbórea presenta copas simétricas o piramidales que se extienden e impiden el paso de luz hacia el sotobosque (Narave y Taylor, 1997), estas características explican el ambiente de sombra y humedad que se genera y que resulta ideal para las especies tolerantes. Por el contrario, en el bosque de Pinus hartwegii la estructura del dosel es irregular, con ramas cortas y extendidas (Narave y Taylor, 1997) que permiten que los rayos solares lleguen al suelo (Rzedowski, 2006). Esta condición promueve la dominancia en cobertura y abundancia de las especies heliófitas, que son, en su mayoría gramíneas y que presentan una gran afinidad a los altos niveles de luz (Grime, 1989). En las plantas, el grado de tolerancia a la sombra define su relación con otros organismos y con el medio. La sombra influye en los elementos bióticos y abióticos del bosque, determinando su estructura y diversidad (Blanco-García, Sáenz-Romero, Martorell, Alvarado-Sosa y Lindig-Cisneros, 2011; Valladares, Laanisto, Niinemets y Zavala, 2016).
En relación a la regeneración, este presenta un patrón de crecimiento poblacional en forma de “J” invertida, donde la densidad se concentra en las clases diamétricas inferiores y disminuye paulatinamente. Esta situación ha sido observada para otros bosques de Pinus hartwegii (Endara et al., 2012; Murrieta-Hernández, Pineda-López, Noa-Carrazana, Mata-Rosas, Zulueta-Rodríguez y Flores-Estévez, 2014) y Abies religiosa (Encina-Domínguez et al., 2008; Endara et al., 2012; Pineda-López, Ortega-Solis, Sánchez-Velásquez, Ortiz-Ceballos y Vázquez-Domínguez, 2013). La regeneración de una especie arbórea puede depender de múltiples factores, entre los que se encuentran el gremio ecológico (heliófita o esciófita), la tasa de crecimiento, mortalidad o depredación de las plántulas, la competencia inter e intraespecífica de los renuevos, así como variables ambientales, bióticas y antrópicas (Yamamoto, 2000; Villavicencio, Santiago, Godínez, Chávez y Toledo, 2012). La regeneración del bosque de pino depende especialmente de la luz (al ser una especie heliófita) y de la frecuencia de incendios superficiales, debido a que favorece la germinación de sus semillas (Ávila-Flores, González-Tagle, Jiménez-Pérez, Aguirre-Calderón, Treviño-Garza y Vargas-Larreta, 2012; Robles-Gutiérrez, Velázquez-Martínez, Rodríguez-Trejo, Reyes-Hernández y Etchevers-Barra, 2016). En cambio, el oyamel puede regenerarse bajo su propio dosel o con una mayor abundancia en los claros del bosque (Lara-González, Sánchez-Velásquez y Corral-Aguirre, 2009).
Estructura del sotobosque
El patrón de dominancia encontrado en el sotobosque de Abies religiosa es distinto al reporte de Villers et al. (1998) para la condición de bosque denso en la misma área de estudio, donde Roldana barba-johannis, Eupatorium mairetianum, Smilax sp. y Senecio callosus. fueron las especies dominantes. Dicho resultado concuerda con los reportes del sistema volcánico transmexicano (Sánchez-González, López-Mata y Granados-Sánchez, 2005), la Sierra Nevada (Sánchez-González y López-Mata, 2003) y el Cofre de Perote (Vázquez, 2014), donde algunas de las especies mencionadas constituyen un elemento importante de la flora.
En el sotobosque en el bosque denso de Pinus hartwegii estudiado la única especie arbustiva dominante es Senecio cinerarioides, mientras que en el estrato herbáceo predominan Alchemilla procumbens y Penstemon gentianoides. Esto concuerda con lo señalado por Villers et al. (1998), en la misma área de estudio, pero difiere de los hallazgos de Sánchez-González y López-Mata (2003), Santillana (2013) y Vázquez (2014) en distintas áreas, donde algunas de las especies dominantes son Acaena elongata, Baccharis conferta, Roldana angulifolia, Vaccinium geminiflorum y Bromus catharticus.
Los bosques analizados comparten, en el sotobosque, dos especies: Alchemilla procumbens y Senecio cinerarioides que están asociadas a perturbaciones. Además de estas especies existen Acaena elongata, Sigesbeckia jorullensis (en el oyamel) y Lupinus montanus (en el pino) que alcanzan una importancia significativa. En el ambiente, la presencia de algunas especies vegetales está relacionada con los eventos de perturbación o las prácticas de manejo que modifican la composición y la riqueza de la comunidad (North et al., 2005; Duguid, Frey, Ellum, Kelty y Ashton, 2013).
En el bosque de oyamel, las perturbaciones más frecuentes son la apertura de senderos y el pastoreo. En cambio, en el bosque de pino ocurren eventos de reforestación, ocoteo, construcción de zanjas de infiltración y pastoreo. De forma general, el pastoreo, la herbivoría o el fuego alteran el establecimiento, la sobrevivencia y el rendimiento de las plantas, sus efectos pueden ser positivos, negativos o generar una reacción directa e indirecta sobre la vegetación (Warner y Cushman, 2002; Husheer, Robertson, Coomes y Frampton, 2006; Mysterud, 2006). Las perturbaciones influyen también en el suelo, los nutrientes y las condiciones del agua (Pokhriyal, Chauhan y Todaria, 2012). Una perturbación en la composición vegetal, puede aumentar la riqueza de especies o reducirla, en ocasiones un nivel moderado de disturbio, genera una elevada riqueza (Royo, Collins, Adams, Kirschbaum y Carson, 2010). El daño de una perturbación depende de su extensión, intensidad o frecuencia (Augustine, Derner y. Milchunas, 2010), por ejemplo, en algunos casos la acción de los herbívoros aumenta la densidad o cobertura en las plantas y favorece la invasión de especies exóticas (Nai-Bregaglio, Pucheta y Cabido, 2002; Vázquez, 2002). En el bosque, el fuego es otro factor clave que modifica los depósitos de biomasa, altera el ciclo hidrológico y transforma la vegetación (Secretariat of Convention on Biological Diversity, 2001; Hoss, Lafon, Grissino-Mayer, Aldrich y DeWeese, 2008). Especialmente en el sotobosque evaluado de Pinus hartwegii, la presencia de Lupinus montanus indica quemas frecuentes y la abundancia de gramíneas favorece la propagación del fuego (Castañeda, Endara, Villers y Nava, 2015).
Conclusiones
La presente investigación contribuye al conocimiento de la riqueza, dominancia y estructura de especies del sotobosque, en bosques densos de Abies religiosa y Pinus hartwegii del APFFNT. En las comunidades evaluadas, el sotobosque presenta la mayor diversidad florística y alberga una gran cantidad de especies nativas que le confieren una fisonomía estructural única a cada ecosistema.
En el sotobosque de Abies religiosa la riqueza de especies es mayor a la del sotobosque de Pinus hartwegii. En esta comunidad las plantas de la familia Asteraceae (Roldana angulifolia, Stevia monardifolia, Roldana platanifolia) son dominantes en la composición florística y algunas como Sigesbeckia jorullensis están asociadas a disturbio. En el sotobosque de Pinus hartwegii la familia Asteraceae fue dominante por número de especies, sin embargo, en cuanto a cobertura y abundancia las especies Calamagrostis tolucensis y Agrostis tolucensis, de la familia Poaceae son predominantes. En esta comunidad la especie arbustiva más importante, por su abundancia y dominancia fue Senecio cinerarioides. La regeneración arbórea en el sotobosque de las comunidades evaluadas es exclusiva de Abies religiosa y de Pinus hartwegii. En ambos bosques por orden de importancia en clase y tamaño se registró un mayor número de plántulas que de brinzales y latizales.

