











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Tectona grandis (teca) en Costa Rica se ha utilizado en proyectos de reforestación con fines comerciales (Boley, Drew y Andrus, 2009), se caracteriza por tener una madera con excelentes propiedades físicas (Tewari, Sukumar, Kumar y Gadow, 2014), mecánicas (Aristide, Christophe, Sossou, Malahimi y Antoine, 2015) y atractivo estético (Boley et al., 2009), lo que propicia que tenga un valor económico susceptible en los mercados internacionales de comercialización maderera (Tewari et al., 2014). En el proceso de establecimiento de T. grandis es fundamental considerar las características edáficas, esta especie requiere de suelos con densidad y acidez baja, con niveles nutricionales elevados, drenajes profundos y poca pendiente (Khanduri, Lalnundanga y Vanlalremkimi, 2008; Zhou, Liang, Xu, Zhang y Huang., 2012). En Costa Rica es común encontrar plantaciones juveniles (menores a cinco años de edad) de T. grandis establecidas en sitios anteriormente dedicados a la ganadería (Boley et al., 2009), presentando niveles de compactación de suelo elevados que afectan directamente el crecimiento y calidad de las plantaciones (Khanduri et al., 2008; Vyas, Mehta, Dinakaran y Krishnayya, 2010; Fernández-Moya y Alvarado 2014).
Para contrarrestar la compactación se han desarrollado actividades como la labranza o mecanización de suelo (Aruga, Hiyamizu, Nakahata y Saito, 2013), que generan cambios en la estructura (Aruga et al., 2013) y en la densidad (Engler, Becker y Hoffmann, 2016), mejoran la aeración (Aruga et al., 2013) y generan variaciones en el movimiento superficial e interno del agua e inclusive en la disponibilidad de nutrientes y materia orgánica (Bottinelli, Hallaire, Goutal, Bonnaud y Ranger, 2014). Lo anterior estimula en el mediano plazo el crecimiento de T. grandis y con ello la reducción de los turnos de cosecha y el incremento en la calidad de la madera (Culshaw y Stokes, 1995).
Para evaluar los efectos de la labranza en el tiempo se han desarrollado múltiples pruebas como análisis de desarrollo radicular (Tewari et al., 2014), análisis nutricionales foliares (Fernández-Moya y Alvarado, 2014), pruebas de compactación, infiltración y estructura de suelo (Yoshifuji et al., 2009) y análisis de índice de área foliar (IAF) (Vyas et al., 2010; Zhang et al., 2011). Este último análisis es uno de los más efectivos por que cuantifica el nivel de crecimiento aéreo de un individuo (Aruga et al., 2013) y el efecto de condiciones ambientales del sitio (Vyas et al., 2010). Además, es una variable fisiológica clave para el desarrollo de modelos de crecimiento (Demarez, Duthoit, Baret, Weiss y Dedieu, 2008; Chianucci, Cutini, Corona y Puletti, 2014), fijación de carbono (Liu, Pattey y Admiral., 2013) y dinámica de desarrollo de especies y poblaciones (Poblete-Echeverría, Fuentes, Ortega-Farias, Gonzalez-Talice y Yuri, 2015; Vyas et al., 2010). El IAF se define como la relación del área de tejido foliar vivo por unidad horizontal de superficie (Marshall y Waring, 1986; Arias, 2004). Es un parámetro que tiende a ser utilizado a nivel individuo (y no extrapolado a una unidad de área) tanto en plantaciones como en bosque natural, debido es un indicador que muestra el grado de desarrollo foliar y salud de un organismo y puede ser relacionado con mayor simplicidad a otras variables fisiológicas (Demarez et al., 2008, Chianucci et al., 2014).
La medición del IAF se puede realizar con método directo o indirecto; el método directo consiste en cosechar el individuo o parte de su copa y, a partir de relaciones alométricas, masa seca de las hojas y área foliar específica (el área promedio de las hojas), se genera el IAF (Mason, Diepstraten, Pinjuv y Lasserre, 2012); es un método de alta precisión, pero destructivo y de un costo económico elevado (Mason et al., 2012; Leblanc y Fournier, 2014). En cambio, el método indirecto no altera al individuo de estudio, facilitando un monitoreo del IAF en el tiempo de una forma precisa y rápida, pero con la necesidad de crear factores de corrección en las mediciones de cada especie a estudiar (Vyas et al., 2010). Entre las metodologías indirectas de más empleadas se destaca la fotografía digital hemisférica. Esta se basa en la utilización de lentes de tipo “ojo de pez” que se incorporan a cámaras fotográficas y teléfonos móviles (recientemente se han desarrollado aplicaciones que permiten recrear el ángulo del lente) y permiten tomar una fotografía angular que es complementada con un software libre. Este software contiene modelos algorítmicos basados en la Ley de Beer-Lamberth que simplifican el procesamiento de la información (Vyas et al., 2010; Mason et al., 2012) y genera el IAF de forma precisa.
Entre los estudios desarrollados para evaluar el efecto de la labranza en el IAF de cultivos forestales se destaca el realizado por Aruga et al. (2013), quienes evaluaron los efectos de la labranza con más de tres especies forestales en Japón, encontrando aumentos de IAF en suelos mecanizados. Por su parte Engler et al. (2016), con plantaciones dendroenergéticas de Eucalipto, encontró aumento en la productividad de biomasa y de IAF de 10% a 15% en suelos labrados. Bottinelli et al. (2014) con especies de eucalipto y pino obtuvieron aumento en la homogeneidad de las plantaciones y mejoras en el desarrollo diamétrico a siete años de la labranza del suelo a 30 cm de profundidad. Sin embargo, de los estudios realizados no se han encontrado investigaciones puntuales con T. grandis que relacionen la labranza del suelo y el IAF como indicador de desarrollo.
Objetivo
Evaluar, después de tres años de establecido el experimento, los efectos de ocho tratamientos de labranza en el crecimiento diamétrico, altura total e IAF en una plantación de 144 meses de edad de T. grandis en la zona norte de Costa Rica. La hipótesis nula del trabajo es que el crecimiento de las plantaciones es independiente del mejoramiento de las propiedades físicas y químicas del suelo en plantaciones que fueron establecidas sin labranza inicial del suelo.
Materiales y métodos
Se evaluaron 48 individuos (seis individuos por tratamiento) de T. grandis de 144 meses de edad (cuando se realizaron las actividades de labranza la plantación tenía 72 meses de edad), con un espaciamiento de 3 m × 3 m, todos los individuos contaron con una poda a 2,5 m de altura a partir de la base, realizada a los 12 meses de edad.
Sitio de estudio
El sitio de estudio se ubicó en una plantación comercial con fines de producción de madera para exportación del Grupo de Reforestación Internacional en Pavón, Alajuela, Costa Rica (10°50’18,51” N; 84°39’40,12” O), a una altitud de 792 m, con una temperatura media anual de 29 °C y una precipitación anual promedio de 2700 mm (Instituto Meteorológico Nacional, 2016). El sitio no había tenido preparación previa del suelo al momento de la plantación, y anteriormente se había dedicado a actividades agrícolas y ganaderas.
Tratamientos
La operación de labranza se realizó en el año 2013 con un tractor marca Same de 65 caballos de fuerza y un arado de cincel. La velocidad del tractor al momento de la actividad fue constante en cada tratamiento, con el fin de obtener un subsolado homogéneo (Tabla 1). Los tratamientos consistieron en una combinación de dos profundidades de subsoleo (25 cm y 50 cm) y tres intensidades diferentes de labranza definidas por la cantidad de cinceles utilizados en el aditamento y la velocidad del tractor al momento de la actividad, formando un total de ocho tratamientos (Tabla 1). Adicionalmente, a todos los tratamientos exceptuando al testigo, se les adicionó una dosis de cal y magnesio (CO3)2 equivalente a 3 Mg ha-1 para tener un suelo con un porcentaje de saturación de acidez inferior a 10%. Los tratamientos se aplicaron en parcelas de 8 árboles × 8 árboles y se acomodaron aleatoriamente dentro de un bloque de 16 árboles × 32 árboles. Antes de la labranza, el suelo presentó una densidad promedio de 1,18 g cm-3 y una alta resistencia a la penetración de 1,08 kg cm-2, posterior a la labranza la densidad media bajó a 1,07 g cm-3 y la resistencia a la penetración a 0,80 kg cm-2.
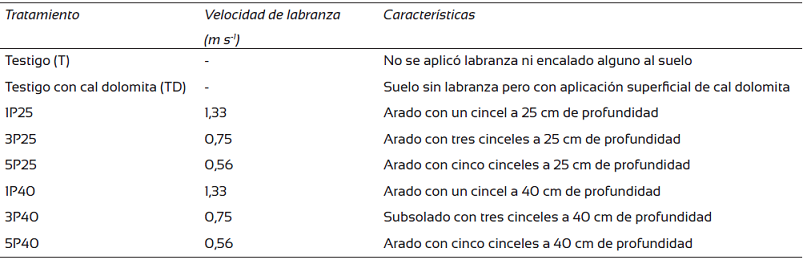
Mediciones alométricas
Se desarrollaron mediciones de altura total y diámetro tanto en el año 2013 como en el 2016; la medición diamétrica se realizó a una altura de 1,3 m sobre nivel del suelo con una cinta diamétrica marca Forestry Suppliers® graduada en milímetros. Mientras la altura total se evaluó con una vara telescópica de 15 m de altura máxima marca Hastings®.
Medición indirecta de IAF
Se consideró el índice de área de la planta (IAP, definido como la sumatoria del área foliar y leñosa aérea de la planta) como parámetro para obtener el IAF indirecto, generado a partir de imágenes fotográficas hemisféricas (obtenidas con la aplicación móvil). Las mediciones se desarrollaron en el mes de marzo de 2016 aplicando la metodología propuesta por Mougin et al. (2014) y Chianucci, Puletti, Giacomello, Cutini y Corona (2015), en clima totalmente nublado y sin presencia de viento.
Para la obtención de fotografías digitales hemisféricas se utilizó un teléfono móvil marca ASUS® Modelo Zenfone 5 con una cámara de 8MP (una resolución de 2024 pixeles × 2024 pixeles en formato RGB) y con la aplicación móvil Fisheye Pro versión 1.2 (R22 software) para Android. Todas las fotografías se tomaron niveladas (controlado con el uso un trípode) a una altura de 60 cm sobre nivel del suelo y con la dirección hacia el norte previamente marcada.
Las fotografías hemisféricas se procesaron con el software CAN_EYE versión 6.3.13 (Weiss et al., 2004), previamente se calibraron según el método descrito en el manual del CAN_EYE (versión 6.3.6) (Weiss, Baret, Smith, Jockheere y Coppin, 2004). Se limitó el grado de visión de las imágenes a 60° y con la opción de recorte en CAN_EYE, se eliminaron las copas de los árboles que rodeaban a cada individuo de estudio con el fin de prevenir las distorsiones y sesgos en el cálculo del IAP; igualmente se configuró un ángulo zenith de 2,5° y azimut de 5° (Chianucci et al. 2015), con un análisis de pixeles de tipo No mixed pixels (2 clases); el algoritmo seleccionado fue el IAF-2000 4 rings debido a que en estudios previos realizados por Chianucci et al. (2015) ha mostrado mayor precisión en especies tropicales.
Medición directa de IAF
En los árboles evaluados se tomaron muestras foliares siguiendo la metodología de Mason et al. (2012), las muestras se escanearon a color y con una calidad de 354 pixeles por centímetro (900 pixeles por pulgada), utilizando un escáner Epson® modelo TX-110. Se calculó el área foliar específica (AFE) con el software WINFOLIA® versión 2012 pro. Posteriormente, las hojas se secaron a 65 °C por 48 horas para determinar el peso seco y con este se determinó el IAF (Ec. 1).
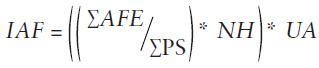 [1]
[1]
Donde,
∑AFE = sumatoria del área foliar específica (m2)
∑PS = sumatoria del peso seco de las hojas (g)
NH = número de hojas contabilizadas en la copa del árbol
UA = unidad de área del suelo (en este caso de 9 m2)
Análisis estadístico
La base de datos se sometió a pruebas de consistencia mediante la realización de gráficos Box Plot. Se evaluó si existían diferencias significativas en los valores de diámetro y en los de altura total de los períodos 2013 y 2016 y, posteriormente, se analizaron las diferencias en crecimiento obtenidas en dicho periodo. Además, se evaluó la existencia de diferencias entre el IAF directo y el indirecto (del año 2016) de cada uno de los tratamientos, para lo cual se aplicó un análisis de varianza de una vía (Andeva), con su respectiva comprobación de normalidad de los residuales y homogeneidad de las varianzas. En los tratamientos donde se obtuvieron diferencias, se procedió a realizar la prueba de comparación múltiple según Tukey (α = 0,05). Además, se realizó un análisis de correlación de Pearson para valorar el grado de relación entre la altura total (del 2016) y el IAF indirecto; según mencionan Piayda et al. (2015), conforme mayor sea la correlación de IAF y altura total, mayor es la efectividad del tratamiento de mecanización. Se generaron modelos de regresión para relacionar las variables IAF indirecto con IAP, para lo cual se siguió la metodología de Mason et al. (2012), que consiste en regresiones lineales (Ec. 2) en las que el IAP es la variable independiente y el IAF indirecto es la variable dependiente.
 [2]
[2]
Donde:
IAF = índice de área foliar
IAP = índice de área de la planta
a = ordenada al origen
b = pendiente
Estos análisis de regresión se realizaron con una significancia de 0,05 en el programa STATISTICA 9.0.
Resultados
Efectos de las actividades de labranza en las variables dasométricas
Los resultados no mostraron diferencias significativas en los valores del diámetro (Tabla 2) tanto del año 2013 (con diámetros que variaron de 8,19 cm a 9,16 cm), como del 2016 (de 10,80 cm a 12,50 cm); con un aumento diamétrico anual de 0,67 cm a 1,27 cm. Con la altura total en el año 2013 se mostraron diferencias significativas en las alturas con variaciones de 8,64 m a 9,39 m sin un tratamiento claramente dominante; sin embargo, para el año 2016 no se encontraron diferenciaciones considerables en las alturas (variación de 9,90 m a 12,31 m) con incrementos anuales de 0,20 m a 1,17 m.
Tabla 2 Valores promedio y desviación estándar del diámetro, altura total e incrementos de crecimiento en individuos de una plantación de T. grandis en la zona norte de Costa Rica.
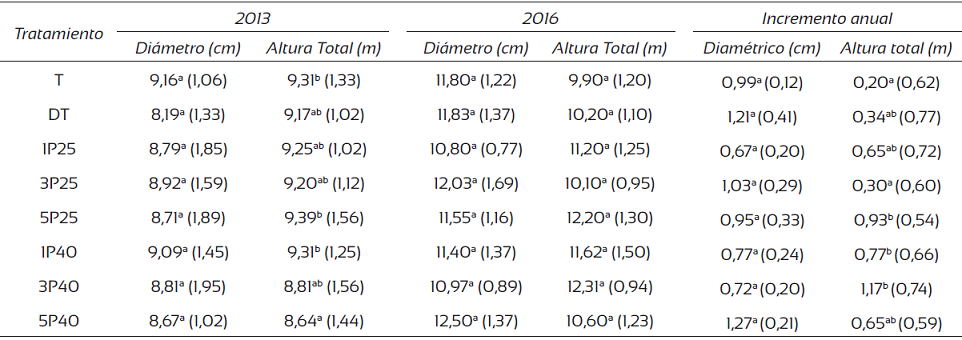
Nota: datos entre paréntesis corresponden a la desviación estándar; letras distintas en cada columna muestran diferencias significativas (Tukey, P < 0,05).
Efectos de la labranza en la estimación del IAF
Un primer resultado indica que se encontró diferencia entre el AFE del testigo (Tabla 3), que presentó un valor medio de 24,79 cm2, en comparación con los seis tratamientos de labranza que mostraron AFE significativamente mayores (en promedio de 57,17 cm2). Al analizar el IAF de método directo se encontraron variaciones de 3,51 m2 m-2 a 5,88 m2 m-2, mostrando dos agrupaciones con diferenciaciones significativas entre sí. El primer grupo conformado por los testigos (T y TD) que arrojaron valores inferiores a 3,75 m2 m-2 y una segunda agrupación conformada por los tratamientos de labranza que mostraron valores superiores a 4,87 m2 m-2, pero sin tener diferenciaciones significativas entre los tratamientos de labranza.
Tabla 3 Valores promedio y desviación del área foliar específica (AFE) e índice de área foliar (IAF) directo e indirecto de una plantación de T. grandis en la zona norte de Costa Rica.
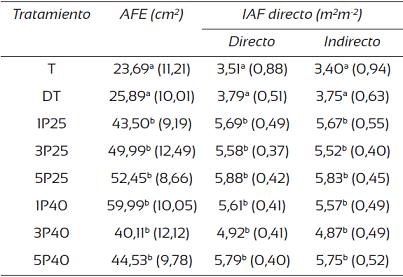
Nota: datos entre paréntesis corresponden a la desviación estándar; letras distintas en cada columna muestran diferencias significativas (Tukey, P < 0,05).
El IAF de método indirecto, presentó valores de 3,40 m2m-2 a 5,83 m2m-2, con una subestimación de los valores de IAF de 15% a 35% con respecto a los IAF directos, pero manteniendo la tendencia de agrupación de T y DT con los valores menores (3,30 m2m-2 a 3,75 m2m-2) y con el grupo de tratamientos con labranza de suelo (superiores a 4,87 m2m-2), que no mostraron diferencias entre sí.
Correlación del IAF y la altura total
Al relacionar los valores de IAF indirecto con la altura total (Fig. 1) se encontró una correlación inferior a 15% para los tratamientos T y TD (Fig. 1a y 1b), mostrando que la variabilidad en IAF de los árboles de ambos tratamientos es amplia. En cambio, los árboles ubicados en el suelo labrado mostraron relaciones superiores a 59%, mostrando una tendencia inversamente proporcional: conforme aumenta la altura total, el IAF decrece. Esta tendencia se muestra con mayor claridad en los tratamientos con labranza superficial: 1P25 (Fig. 1c), 3P25 (Fig. 1d) y 5P25 (Fig. 1e) con correlaciones entre 65% y 81%; en comparación con los tratamientos de labranza profunda 1P40 (Fig. 1f), 3P40 (Fig. 1g) y 5P40 (Fig. 1h), que mostraron correlaciones moderadas entre 59% y 75%. Sin embargo, es relevante destacar que aun con esta tendencia en las correlaciones, la diferenciación entre tratamientos no es significativa.
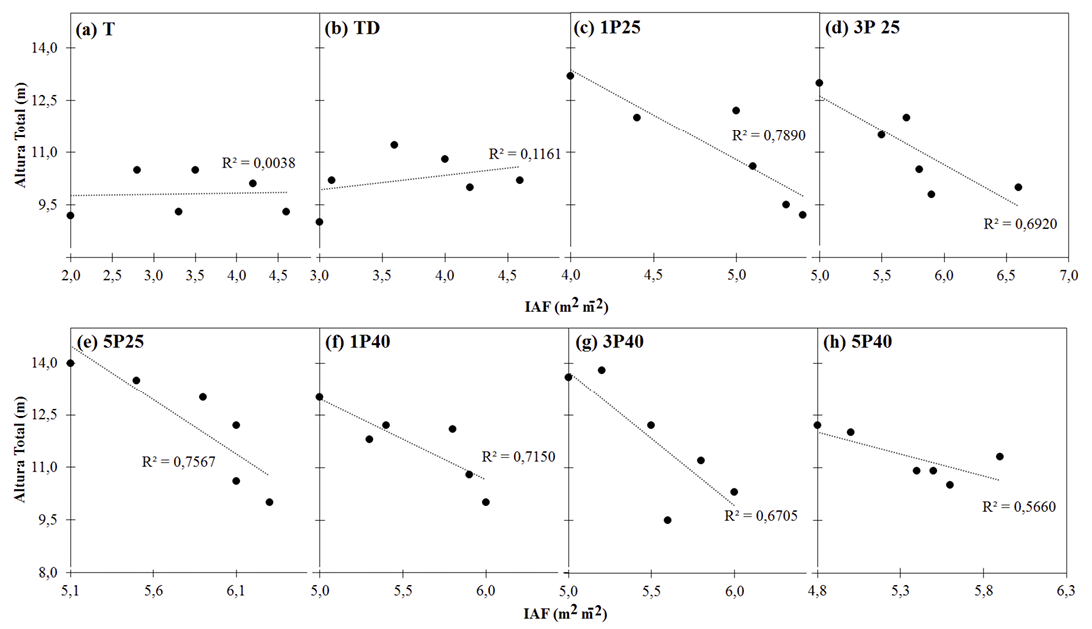
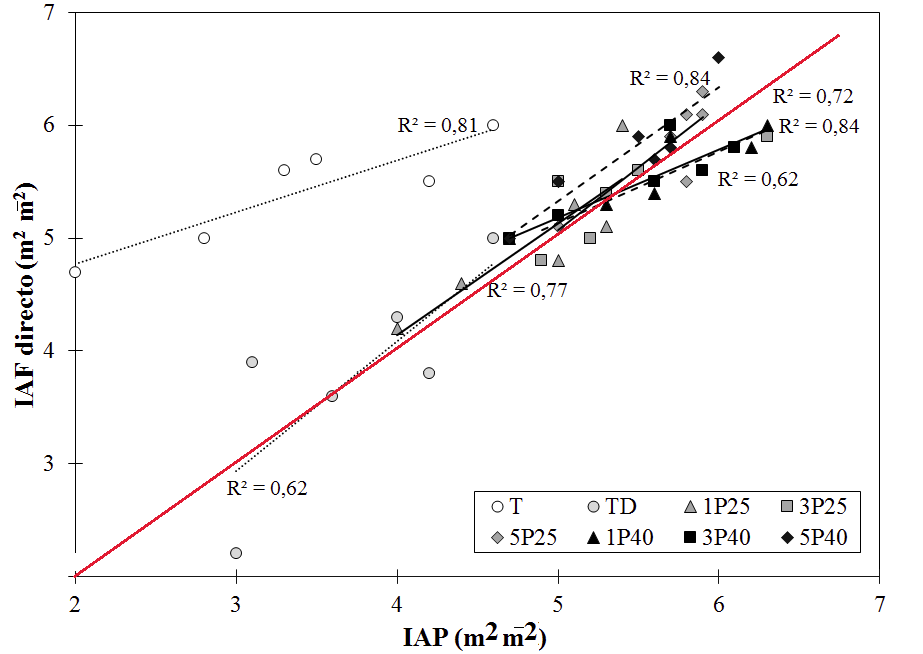
Modelos de regresión del IAF a partir de IAP
De manera general, los modelos de IAF indirecto mostraron la tendencia hacia un aumento lineal de IAF en función del IAP (Fig. 2). Los tratamientos 1P40 y 3P40 tendieron a sobrestimar el IAF cuando los valores de IAF eran superiores a 5,0 m2m-2 y a subestimarlo cuando los valores de IAP eran inferiores a 5,3 m2m-2; en cambio el comportamiento de los seis tratamientos restantes fue a subestimar el IAF entre 10% y 40% con respecto a los valores del IAP. Al evaluar los modelos considerados, se encontró que los modelos de T y TD (IAF=0,46·IAP+3,86 y IAF=0,46·IAP-0,54) mostraron la menor precisión, debido a que son los que más subestiman el IAF, presentan los coeficientes de determinación menores y tienen errores estándar superiores a 0,19 (Tabla 3); en cambio con los modelos de labranza a 25 cm de profundidad, se obtuvieron subestimaciones de IAF entre 5% y 15%, siendo las menores en todos los tratamientos evaluados, con coeficientes de determinación ente 0,72 y 0,84 y errores estándar inferiores a 0,16. En cambio, los modelos con labranza a 40 cm presentaron una tendencia a subestimar el IAF especialmente en árboles de porte pequeño y de sobreestimarlo en árboles de mayores dimensiones, con coeficientes de determinación que variaron de 0,62 a 0,84 y errores estándar intermedios que oscilaron entre 0,09 y 0,16.
Discusión
Los resultados obtenidos (Tabla 2) no evidenciaron efectos de la labranza en el crecimiento diamétrico, esto se debe entre otros aspectos a dos factores principales: 1) el crecimiento de T. grandis en etapa juvenil (de tres a ocho años de edad) se enfoca en el desarrollo apical, como estrategia de sobrevivencia de la especie para mantenerse en un sitio; por esto el crecimiento diamétrico se incrementa posterior al sexto año de vida (Khanduri et al., 2008). 2) No ha transcurrido el suficiente tiempo para ver reflejado el efecto de la mejora del suelo (que se evidenció con la disminución de resistencia a la penetración y de densidad del suelo) en el crecimiento en el diámetro de los individuos; esto último se debe a que es un proceso de readaptación de los árboles y de su sistema radicular a los suelos mejorados. Según Tausz, Grulke, Richter y Kikuta (2014), las plantas inician con aumento radicular y foliar como promotores de crecimiento apical en primera instancia y el crecimiento diamétrico se evidencia de 24 a 60 meses después de la labranza (dependiendo de la edad y la especie), por cual no es de esperar cambios significativos a los 36 meses.
En el caso de la altura total, los resultados (Tabla 2) mostraron una homogeneización en esta variable, producto de la labranza. Manchado y Silva (2001) mencionan que, en suelos compactados, el estrés fisiológico es elevado y limita el crecimiento por la falta de disponibilidad de nutrientes (mayor resistencia de liberación nutricional por parte del suelo) y por consiguiente la capacidad crecimiento radicular. En este contexto, la labranza genera una mejora en las condiciones que estimulan el desarrollo radicular, que permite mayor absorción de nutrientes y una reactivación apical, la cual resulta en un aumento de la altura. Bottinelli et al. (2014) mencionan, para dos especies de eucalipto, que la mejora del suelo proporcionó aumentos de 15% a 20% en el crecimiento apical y de 10% a 20% en el desarrollo foliar. Por su parte, Alvarado (2016) destaca que se pueden encontrar cambios significativos de alturas en árboles en terrenos con labranza 24 meses después de la actividad; este debe ser el tiempo necesario para una recuperación de la micro fauna liberadora de los nutrientes en el suelo, cuyas poblaciones se pueden ver mermadas por los cambios físicos y la cantidad de biomasa radicular viva. Es importante destacar que los árboles de los tratamientos testigo, presentaron las tasas de crecimientos de altura menores debido a que el individuo continúa en un estado de estrés y, según destaca Pancel (2016), conforme el árbol tenga una edad mayor el estrés aumentará, el crecimiento disminuirá y se corre riesgo de muerte del individuo.
Se considera que las diferencias de IAF entre las unidades testigo (T y DT) y los seis tratamientos de labranza se debieron a la estimulación generada por la mecanización. Bottinelli et al. (2014) mencionan que el desarrollo foliar es altamente susceptible a cambios en el suelo, esto debido a que un aumento o reducción del sistema radicular incide en la capacidad de absorción de agua y macronutrientes por parte de la planta que son precursores en la formación foliar. Aquellos suelos con mejoras en sus propiedades físicas, pueden resultar en un aumento del área foliar de 20% a 65%, como lo obtenido por Culshaw y Stoke (1995) con dos especies de eucalipto, que a 12 meses poslabranza presentaron aumentos del IAF de 10% a 30%; estos autores resaltan que un aumento de área foliar y por consiguiente en los procesos de fotosíntesis, generaron un estímulo en el crecimiento de 10% durante primeros 18 meses poslabranza. El aumento foliar según mencionan Culshaw y Stoke (1995) y Bottinelli et al. (2014) es de los primeros indicadores de los efectos fisiológicos de las actividades de labranza, que se pueden explicar en tres fases: (i) recuperación radicular (en el que los árboles empiezan a recuperar volumen radicular y repoblar áreas del suelo; generalmente esto tarda de 6 a 12 meses en árboles con menos de tres años de edad), (ii) regeneración foliar (aumento del IAF a 18 meses poslabranza y una estimulación del crecimiento longitudinal del individuo) y (iii) crecimiento total (estimulación del crecimiento radial y formación de nuevos ejes en el individuo).
Las mejoras en la labranza del suelo y la estimulación en el crecimiento se evidenciaron claramente en los análisis de correlación (Fig. 1). La homogeneidad en los individuos con labranza en comparación con los testigos (T y DT) se debe a que los testigos muestran evidencia de una compactación mixta (Carballas et al., 2015), que es resultado del uso anterior del suelo (ganadería). Por su parte, Srinivasa et al. (2017) mencionan que la labranza elimina el efecto de la compactación mixta generalmente en los primeros 40 cm de profundidad del suelo, permitiendo no solo homogeneizar la compactación del suelo sino aumentar la permeabilidad de la cal y magnesio y con ello mitigar los efectos de la acidez; además promueve la recuperación de colonias bacterianas fijadoras de nitrógeno y eliminación de otras coberturas herbáceas que pueden limitar el desarrollo de la especie (Meinzer et al., 2011).
Piayda et al. (2015) mencionan que, como producto de la mejora en las condiciones edáficas en las plantaciones, los árboles con mayor porte tienden a producir pocas hojas, pero con un AFE significativa mayor en comparación con los árboles de mediano y bajo porte, que presentarían una mayor cantidad de hojas pero de menor tamaño como respuesta a la competición por luminosidad y espacio. Este comportamiento no se da en condiciones de suelo con limitaciones tanto en propiedades físicas como químicas que generan en las plantaciones una alta variabilidad en altura, debido a que los individuos concentran su energía en adaptarse a la condición de estrés como medida de supervivencia (Pugnaire y Valladares, 1999; Pessarakli, 2014). Esto además incidió en los modelos de regresión (Tabla 4) que en los tratamientos testigo presentaron un error estándar mayor, menor precisión y por ende subestimación del IAF (Fig. 2) con respecto a los tratamientos de labranza. Estos, por la homogeneización de la plantación, mostraron modelos con menor error, mayor precisión y subestimaciones menores a 30%. Lo anterior es similar a lo encontrado por Mason et al. (2012), con árboles de pino, quienes justificaron que la mejoría en las condiciones de crecimiento se debieron a mejorías en las condiciones edáficas, que impactaron positivamente en el desarrollo de cada individuo en la plantación.
Tabla 4 Modelos de regresión del modelo IAF=a·IAP+b para determinar el IAF a partir del IAP de una plantación de T. grandis en la zona norte de Costa Rica una significancia de 0,05.

IAF = índice de área foliar; IAP = índice de área de la planta. Modelos de correlación realizados con un P de 0,05
La no presencia de diferencias significativas entre los tratamientos de labranza se puede explicar por el periodo de evaluación que no ha sido suficientemente amplio. Mapa (1995) menciona que los procesos de mejora del suelo en especies tropicales son efectivos a 72 meses de la labranza y las diferencias en los árboles se producen después de los 84 meses; esto debido a que es la edad en que alcanzan la madurez y las leves mejorías en suelo pueden ser aprovechadas para su adaptación. En otras palabras, la reestructuración del suelo poslabranza no se ha dado en su totalidad; Clemens et al. (2010) destacan que la formación de estructura y macroestructura y la incorporación de material orgánico en el suelo mecanizado son de lento desarrollo, debido a que es un proceso que depende de factores internos del suelo como son presencia de arcillas, limos y arcillas además de variables externas como precipitación y viento.
A pesar de que en este tipo de experimentos se esperan respuestas más inmediatas, la realidad y las experiencias de otros estudios indican que hay un período de reestructuración del suelo y, en este caso, del sistema radicular de los árboles, que requieren evaluaciones en períodos más prolongados. Definitivamente variables relacionadas con el área foliar son clave en la evaluación de este tipo de experimentos. No obstante, los aportes en el presente estudio ayudan en la toma de decisiones con respecto al manejo de las plantaciones establecidas. La principal recomendación a la luz de los resultados es que las acciones de corrección de los factores limitantes en el suelo, sean físicos, químicos o en combinación, deben formar parte de las labores previas al establecimiento de una plantación forestal.
Conclusiones
Los árboles de T. grandis, tres años después de la labranza, no presentaron diferencias significativas en el crecimiento diámetrico, debido principalmente al corto período de la evaluación; la especie estudiada centra su desarrollo en el crecimiento apical. En cambio, se presentó una homogeneización de la altura total entre los tratamientos debido a que esta es una variable de respuesta a corto plazo.
EL IAF mostró diferencias significativas entre los tratamientos, obteniendo que la mecanización aumentó hasta en 20% el IAF en comparación con los testigos (T y TD); que presentaron un retardo del crecimiento debido al estrés producto de la compactación del suelo, que limita la disponibilidad de nutrientes y de agua, retardando el desarrollo radicular.
La no diferenciación entre los tratamientos de labranza se debe a que el periodo de evaluación es aún muy corto para evidenciar sus efectos en el desarrollo de los árboles. Sería ideal realizar la medición 60 meses después de la labranza y correlacionarse con la densidad y resistencia a la penetración del suelo.

