











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
Pinus greggii Engelm. es una especie forestal que enfrenta diferentes niveles de riesgo y de perturbación en sus rodales nativos, pero también es una especie con un alto potencial para programas de restauración y establecimiento de plantaciones comerciales, ya que posee resistencia a la sequía (López y Muñoz, 1991) y a daños por heladas. La especie tiene altas posibilidades para emplearse en plantaciones en México (Donahue y López, 1996) e incluso ya se emplea en plantaciones comerciales en otros países como Sudáfrica, en localidades con baja precipitación donde otras especies de pino no han prosperado (Dvorak y Donahue, 1993).
El área de distribución natural de Pinus greggii se encuentra entre los 20º 13’ y los 25º 29’ de latitud Norte, con una discontinuidad de 360 km en la parte central. Las poblaciones del norte se conocen como P. greggii var. greggii y las del sur como P. greggii Engelm. var. australis Donahue y López. El intervalo altitudinal de la especie es de 1400 m a 2613 m snm. Las poblaciones del sur se encuentran a menor elevación, con mayor temperatura media y precipitación anual (aproximadamente: 1720 m, 17.5 °C y 1370 mm) que las poblaciones de la región norte (aproximadamente: 2450 m, 13 °C y 640 mm); por lo tanto, se puede afirmar que el intercambio genético entre las dos regiones es muy difícil debido a la separación entre las poblaciones (Ramírez, Vargas y López 2005).
Al llevar a cabo plantaciones es necesario conocer los orígenes o procedencias adecuadas para cada sitio. Los ensayos de procedencias son el procedimiento experimental más empleado para comparar la respuesta y la productividad de la semilla que proviene de diferentes poblaciones en sitios ambientales diferentes previamente caracterizados (Zobel y Talbert, 1988).
El patrón de crecimiento en altura es el determinante primario de las diferencias en altura anual y en altura final de los individuos (Zobel y Talbert, 1988). Las coníferas presentan un crecimiento monopódico, lo que origina un tallo principal constituido por secciones sucesivas de formación y alargamiento anual; debido a la ubicación apical y al alargamiento anual de estas nuevas secciones, se les ha denominado brote terminal o brote anual (Zavala, 1986). Lanner (1976) reconoce cuatro principales patrones de crecimiento anual en altura de las especies forestales, que clasifica en: crecimiento libre, crecimiento fijo, crecimiento fijo-libre y crecimiento fijo-fijo.
El inicio y la terminación del crecimiento en especies forestales están influenciados por un fenómeno fisiológico denominado letargo, el cual está asociado a fotoperiodos cortos y bajas temperaturas. El patrón general para las especies de clima templado establece que en otoño entran en quiescencia o pre-letargo debido a factores ambientales; en invierno se acentúa el letargo entrando en receso debido al ambiente interno de la yema (factores fisiológicos internos y factores genéticos). En este periodo se da una baja actividad mitótica y es cuando la planta presenta la mayor resistencia a las bajas temperaturas. A finales del invierno y principios de primavera las plantas entran de nuevo en una etapa de quiescencia post-letargo (Viveros y Vargas, 2007).
Con el propósito de estudiar la fenología en árboles se han realizado diversos tipos de estudios. Por ejemplo, en un ensayo de 20 procedencias de Pinus engelmannii Carr. establecido en Tecamac, Edo. de México se evaluó el nivel de variación geográfica en la supervivencia, el crecimiento anual en altura y la fenología del brote terminal (Rodríguez et al., 2000). De igual forma en España se estudió la fenología del crecimiento de Pinus sylvestris L. durante dos años consecutivos en procedencias españolas con la finalidad de identificar la procedencia apta para realizar plantaciones en la región de Aragüés del Puerto, Huesca (Alía, Galera, Agúndez y de Miguel, 1999). En México existe escasa información sobre el desarrollo del brote terminal de Pinus greggii, y la intención a futuro es realizar plantaciones de productividad en la Sierra de Arteaga, Coahuila. El ensayo de tres procedencias ayuda a identificar los árboles con crecimientos de su yema terminal dentro del periodo favorable evitando presentar daños importantes en las zonas meristemáticas que repercuten en el crecimiento final anual.
Objetivo
Evaluar el patrón de crecimiento del brote terminal en árboles de tres procedencias de Pinus greggii durante dos años plantados en Los Lirios, Arteaga, Coah.
Materiales y métodos
El ensayo de procedencias de Pinus greggii se localiza en el Campo Agrícola Experimental Sierra de Arteaga (Caesa) de la Universidad Autónoma Agraria Antonio Narro, en Los Lirios, Arteaga, Coah., ubicado a aproximadamente 45 km de la ciudad de Saltillo, Coah., (25° 23’ N a 25° 24’ N; 100° 36’ O a 100° 37’ O; 2,280 m snm) (Instituto Nacional de Estadística y Geografía [Inegi], 2000), con una temperatura media anual de 13.6 °C y una precipitación promedio anual de 521.2 mm. Los meses de mayor precipitación son de abril a septiembre (Conagua, 2000), con un tipo de clima Cb(x’)(wo)(e)g que corresponde a un clima templado con verano fresco y largo (García, 1987).
La plantación se llevó a cabo el 5 de agosto de 2005. Se plantaron 910 plantas en cepa común, distribuidas en marco real a 2.5 m entre plantas. La plantación tiene un diseño experimental de bloques completos al azar, con cinco procedencias, cuatro de ellas son del estado de Coahuila (Puerto Conejos, Santa Anita, Jamé y Los Lirios) y una de Nuevo León (El Tarillal), distribuidas en 13 bloques. En cada procedencia de cada bloque se usaron 14 plantas.
Con base en una evaluación previa realizada en diciembre de 2006, se identificaron tres procedencias, Puerto Conejos, El Tarillal y Los Lirios (Tabla 1), que contrastaban en su altura total, las cuales fueron seleccionadas para la medición del alargamiento del brote terminal, utilizando seis plantas por unidad experimental en cinco bloques.
Tabla 1 Localizacion geografica de las procedencias de Pinus greggii Engelm., evaluadas en el presente estudio, del ensayo establecido en el Caesa, Arteaga. Coah.

Fuente: Curiel, 2005.
Para registrar el alargamiento del brote principal (crecimiento en altura), en cada una de las plantas seleccionadas se marcó con pintura roja vinílica en la base de la yema principal, el 3 de marzo de 2007 y el 26 de enero de 2008. La marca fue considerada como la medida cero y a partir de ella se comenzó con el registro de datos de campo con diferencias entre cada medida de una a dos semanas. En el año 2007 el registro fue hasta el 13 de septiembre y en el año 2008 hasta el 22 de noviembre. Las lecturas se registraron con base en el número de días transcurridos a partir del 1 de enero de cada año. Para realizar las mediciones se utilizó una regla graduada en centímetros, con resolución de 1 mm, los datos de incrementos se registraron en milímetros.
Los incrementos acumulados se obtuvieron sumando las diferencias desde la medida cero hasta la última medición realizada. Los incrementos parciales son la diferencia entre un incremento respecto a la medición anterior o siguiente. Se realizaron análisis estadísticos simples para obtener la media y medidas de dispersión de los incrementos parciales y acumulados.
También se realizaron correlaciones simples (correlaciones de Pearson) para cada procedencia entre el crecimiento (parcial y acumulado) y las variables ambientales: temperatura mínima, temperatura máxima, temperatura media, precipitación y fotoperiodo. Debido a que los valores obtenidos de temperatura y precipitación de la estación meteorológica de San Antonio de las Alazanas, Arteaga, Coah., para los años 2007 y 2008 se registraron como promedio para cada mes, fue necesario obtener los crecimientos acumulados y parciales lo más cercano a cada mes, para hacer las correlaciones. En el caso del fotoperiodo el valor que se usó fue el correspondiente al día medio de cada mes, para la latitud del Caesa.
Asimismo, se realizaron análisis de varianza con los datos de cada fecha de evaluación por procedencia. Cuando hubo diferencias estadísticamente significativas se realizó la prueba de comparación de medias Tukey.
Los análisis de varianza se realizaron usando el siguiente modelo estadístico:
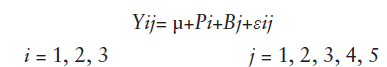
donde:
Yij = Valor de la observación en la i-ésima procedencia, en el j-ésimo bloque.
μ = Valor del efecto presente respecto a la media general.
Pi = Efecto de la i-ésima procedencia.
Bj = Efecto del j-ésimo bloque.
εij = Efecto de la interacción entre la i-ésima procedencia y el j-ésimo bloque (error experimental considerado).
Resultados y discusión
Crecimiento del brote terminal durante el año
El alargamiento del brote terminal en el año 2007 fue superior y comenzó para las tres procedencias alrededor del 11 de marzo (día 68) y en el año 2008 alrededor del 16 de febrero (día 47) (Fig. 1). El crecimiento total promedio de las tres procedencias en el año 2007 fue de 29.97 cm y en el año 2008 de 19.86 cm (Fig. 1), es decir, en el año 2008 las procedencias crecieron solamente dos terceras partes de lo que crecieron en 2007. La explicación más probable para este comportamiento es la diferencia en precipitación, ya que mientras en el año 2007 la precipitación promedio en Coahuila fue de 522.3 mm, en 2008 fue de 459 mm; además en 2008 se presentaron temperaturas máximas promedio superiores a las del año 2007 (Conagua, 2017).
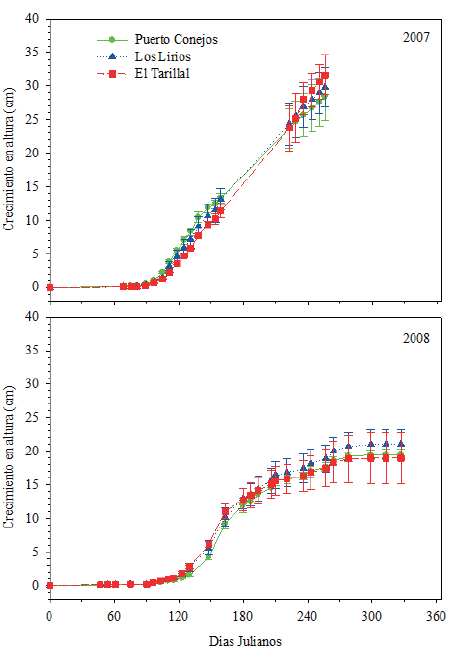
Fuente: Se muestran los valores del error estándar para cada valor promedio en la fecha de medición.
Figura 1 Crecimiento en altura (cm) durante los años 2007 y 2008 de tres procedencias de Pinus greggii Engelm., evaluadas en una plantacion experimental en el Caesa, Arteaga, Coahuila.
En el año 2007 los registros muestran que hasta el 13 septiembre continuó el crecimiento, lo que corresponde a un período de 131 días; sin embargo, durante ese año no se continuó la evaluación después de septiembre, por lo que no se tiene certeza del tiempo preciso de la finalización del crecimiento en ese año. En 2008 se tiene la certeza que las procedencias dejaron de crecer hasta el día 25 de octubre (día juliano 299) lo que corresponde a un periodo de crecimiento de 203 días. De manera que aun cuando en 2007 se presentó un menor período de crecimiento, la tasa de crecimiento fue mayor, lo que se aprecia en mayor pendiente del brote de elongación (Fig. 1).
En el año 2007 desde mayo hasta septiembre se observa un constante crecimiento para las tres procedencias, mientras que para 2008 la mayor tasa de crecimiento se presentó a partir de la de crecimiento fueron 0.28 cm día-1 en mayo en 2007 y 0.33 cm día-1 en junio en 2008. Mientras que las menores tasas de crecimiento registradas se presentan en febrero y octubre con valores alrededor de 0.01 cm día-1 y 0.03 cm día-1.
Los factores ambientales que mejor se asociaron con la tasa de crecimiento en cada una de las tres procedencias durante el periodo de evaluación fueron: en el año 2007 la temperatura mínima (0.79 < r < 0.84) seguida de la temperatura media (0.64 < r < 0.70); mientras que para el año 2008 fue el fotoperiodo (0.64 < r < 0.73) seguido de la temperatura mínima (0.64 < r < 0.69), todos ellos de forma positiva (Tabla 2).
Tabla 2 Correlacion de Pearson entre tasa de crecimiento y temperatura media mensual (TMe), temperatura mínima mensual (TMi) y fotoperiodo (FOT) para tres procedencias de Pinus greggii Engelm. establecidas en el Caesa, Arteaga, Coah.

a Valor de correlación de Pearson. b Probabilidad. c n=número de pares de valores empleados en la correlación.
Se obtuvieron correlaciones positivas que indican que al incrementarse el valor de la temperatura mínima y la cantidad de horas luz, aumenta la tasa de crecimiento, encontrando los mayores valores a mediados del año y, a medida que disminuye la temperatura y el fotoperiodo, también lo hace la tasa de crecimiento. Esta misma situación la describe Hocker (1984) para diversas especies, denotando que la temperatura y el fotoperiodo son determinantes para que las yemas invernales inicien y terminen su letargo. Además de que también tiene fuerte influencia sobre la tasa de crecimiento, dado que la cantidad de horas luz es mayor en el verano y menor en el invierno.
En el presente estudio se encontró que el crecimiento del brote terminal tuvo una duración aproximada de 200 días en las tres procedencias de Pinus greggii evaluadas. Si solamente se considera el periodo entre el momento que tiene 10% y 90% del crecimiento total, entonces se tendrían 145 días y 197 días para 2007 y 2008, respectivamente. Esto es notoriamente diferente con lo encontrado para P. patula en Texcoco, Mex., donde se registró crecimiento durante más de 300 días, desde finales del mes de febrero hasta finales del mes de diciembre (Gómez, 1993) y con lo hallado para P. sylvestris en Aragüés del Puerto, Huesca, España, donde se registró un periodo de crecimiento desde marzo hasta finales de junio, pero el período entre el momento que tiene 10% y 90% del crecimiento total del año fue de solamente 41 días (Alía et al., 1999).
Puede notarse que los estudios se realizaron en latitudes diferentes. Las procedencias reportadas de P. sylvestris son nativas de Europa y la plantación donde se evaluó es una latitud alrededor de los 42º Norte (Alía et al., 1999). Las procedencias de P. greggii var. greggii evaluadas en el presente estudio, así como el lugar de la plantación se encuentran a la latitud de los 25º Norte (Inegi, 2000). Mientras que la plantación de P. patula que se evaluó en Texcoco se encuentra alrededor de los 19º Norte (Gómez, 1993).
La latitud es un factor que explica en parte el resultado del número de días de crecimiento que tienen las plantas durante el año, porque influye en la temperatura y el fotoperiodo. De tal manera que afecta directamente el inicio y terminación del letargo del brote terminal (Hocker, 1984). A menor latitud, es decir más cerca del Ecuador, las temperaturas mínimas son menos restrictivas para el crecimiento de las plantas, mientras que en latitudes mayores, se presentan como un factor limitante para el crecimiento, ya que se presentan más temprano, al inicio del invierno o finales del otoño, y más tarde, a finales del invierno e inicios de la primavera. Asimismo, el fotoperiodo tiene mayor oscilación conforme aumenta la latitud, de manera que en el invierno son pocas horas luz y en el verano son muchas, comparadas con latitudes menores (Torres, 1995). Existen especies de árboles que prácticamente no dejan de crecer durante todo el año: un ejemplo son los árboles tropicales, ya que sus ciclos de crecimiento rítmicos pueden ser anuales, bianuales o irregulares (Tomlinson y Longman, 1983).
Esto hace entendible que las plantaciones forestales comerciales de rápido crecimiento que se realizan en el mundo se encuentran en muchos de los casos en los trópicos, con tasas de crecimiento muy elevadas, por ejemplo, en coníferas se presentan crecimientos en volumen de hasta 25 m3 ha-1 año-1 en Brasil y hasta 40 m3 ha-1 año-1 en latifoliadas en Indonesia (Dvorak y Hodge, 1998), donde la elongación del brote terminal o crecimiento en altura es uno de los principales elementos del volumen del fuste de los árboles.
Variación entre procedencias
Para el año 2007 el análisis de varianza mostró diferencias significativas (P < 0.05) entre las tres procedencias evaluadas en siete fechas de evaluación (días 89, 97, 105, 111, 118, 125, 131 y 138), que corresponde a los meses de marzo a mayo. Mientras que para el año 2008 no se presentaron diferencias estadísticamente significativas (P < 0.05) entre las tres procedencias en ninguna de las fechas evaluadas. Es probable que el no encontrar diferencias se deba al alto valor del error estándar (Fig. 1) lo cual se ve reflejado en el alto coeficiente de variación del análisis de varianza de cada fecha, que presentó valores superiores a 30%.
En el año 2007, los árboles de Puerto Conejos presentaron mayor crecimiento del brote terminal entre marzo y mayo. Por ejemplo, el día 89 la procedencia Puerto Conejos presentó un crecimiento de 0.48 cm, mientras que la de El Tarillal solamente registró 0.25 cm. En el día 147 Puerto Conejos presentó 11.81 cm y El Tarillal 9.47 cm (Tabla 3).
Tabla 3 Valores promedio del crecimiento acumulado en altura (cm) de tres procedencias de Pinus greggii Engelm. evaluadas en el año 2007 en una plantación experimental en el Caesa, Arteaga, Coah.

*valores promedio en una columna con la misma letra son estadísticamente iguales (P ≤ 0.05) de acuerdo con la Prueba de Tukey
La procedencia local (Los Lirios) para el año 2007 en ningún momento sobresalió de las otras dos procedencias evaluadas (Puerto Conejos y El Tarillal). En las fechas en que se presentaron diferencias significativas, en algunos casos, la procedencia de los Lirios fue igual que El Tarillal y en otras fechas fue la de menor crecimiento (días 111 y 125). Sin embargo, después del día 147 y hasta el final del año ya no hubo diferencias significativas entre las tres procedencias en el crecimiento acumulado del brote, al igual que en el año 2008 que en ninguna fecha de evaluación se presentaron diferencias estadísticamente significativas.
En otras plantaciones experimentales de P. greggii tampoco se encontraron diferencias entre las procedencias evaluadas. Ornelas (1997), al evaluar tres procedencias en el Caesa, una de ellas del Cañón de Jamé, otra de Los Lirios, ambas de la Sierra de Arteaga y una tercera del Ejido Cuauhtémoc, Saltillo, Coah., encontró que a los cuatro años siete meses de la plantación no hubo diferencias en el incremento en altura. En la misma plantación, Contreras (2005) y Cornejo, Bucio, Contreras, Valencia y Flores (2005) hallaron que a los 12.6 años no se presentan diferencias en altura entre las procedencias; sin embargo, al evaluar el crecimiento relativo en altura a uno, dos y 12.6 años se encuentra que la procedencia Los Lirios es superior a las otras dos procedencias. Asimismo, en un ensayo de nueve procedencias de P. greggii var greggii establecido en el Ejido 18 de Marzo, Galeana, N. L. no se encontraron diferencias para la variable altura entre las procedencias evaluadas se encontraban Puerto Conejos y Los Lirios (Hernández, 2005).
Sin embargo, en otras plantaciones experimentales de P. greggii si se han encontrado diferencias entre procedencias. Por ejemplo, en dos plantaciones establecidas en la Mixteca Alta de Oaxaca, donde se probaron 13 procedencias, seis de la var. greggii y siete de la var. australis, se encontraron deferencias entre procedencias en altura total y otras variables a los 2.5 años de plantado, de la var. greggii sobresale la procedencia Puerto Conejos, que supera a Los Lirios (Valencia, Velasco, Gómez, Ruiz y Capó, 2006). En otra plantación establecida en Patoltecoya, Puebla, se registraron diferencias entre procedencias en altura a los 2.9 años de plantado; es importante hacer notar que entre las procedencias del norte también sobresale Puerto Conejos por encima de Los Lirios (López, Vargas, Ramírez y López, 1999).
Fuera del país también se han realizado ensayos con P. greggii y se han encontrado diferencias entre procedencias, aun dentro de la var. greggii. En Brasil y Sudáfrica Los Lirios se ha manifestado como la de mejor crecimiento en altura (Dvorak, Kietzka y Donahue 1996), sin embargo en el ensayo de procedencias establecido en Liriossu, sitio de origen, dicha procedencia no es la más sobresaliente.
Lo anterior parece mostrar que en los ensayos que se establecen cerca del área de distribución de las procedencias es más difícil de detectar diferencias en algunas variables como es el crecimiento en altura; pero al establecer los ensayos en lugares más distantes, donde el ambiente es diferente al de su hábitat nativo, es más fácil detectar diferencias entre procedencias. En las especies con amplia distribución es más fácil que se presenten diferencias entre procedencias (Zobel y Talbert, 1988) y más aun si los ensayos se establecen fuera de su área de distribución.
Los estudios genéticos de P. greggii señalan que existen diferencias genéticas importantes entre procedencias y poca variación genética dentro de procedencias, debido en parte al aislamiento de las propias poblaciones (Ramírez, Vargas, Jasso, Carrillo y Guillen, 1997; Parraguirre, Vargas, Ramírez, Azpiroz y Jasso, 2002), por lo que debiera encontrarse diferencias entre procedencias para diversas variables. El no encontrar diferencias entre procedencias para la variable altura en muchos casos probablemente se debe a que es una variable que depende en gran medida del ambiente, es decir, es una variable de poco control genético como lo han descrito Zobel y Van Buijtenen (1989) en diversas especies. En México se conocen pocos estudios sobre heredabilidad en diferentes especies de coníferas, para Pinus greggii se registra una heredabilidad de 0.44 en altura y de 0.30 para longitud promedio de ciclos de crecimiento (López-Ayala, Vargas-Hernández, Ramírez-Herrera y López-Upton, 1999) mientras que para P. patula se documenta una heredabilidad de 0.14 en altura de los árboles (Valencia, Vargas, Molina y Jasso, 1996).
Conclusiones
El inicio y la terminación del crecimiento del brote terminal en los árboles de P. greggii del ensayo de procedencias están fuertemente influenciados por la temperatura mínima y el fotoperiodo.
El crecimiento del brote terminal en árboles de P. greggii, en las procedencias establecidas en la Sierra de Arteaga, Coahuila, en promedio inicia a los 96 días y termina hasta los 299 días, teniendo un periodo de 145 días y 197 días en que se presenta alrededor de 90% del crecimiento total para los años 2007 y 2008, respectivamente.
Existen diferencias entre procedencias en el crecimiento del brote terminal que solamente se detectaron en algunas fechas de evaluación del año 2007. Sin embargo, no se detectó variación dentro de procedencias a través del tiempo ni entre los años evaluados.

