










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkMadera y bosques
versión On-line ISSN 2448-7597versión impresa ISSN 1405-0471
Madera bosques vol.21 no.3 Xalapa sep./dic. 2015
Artículos científicos
Composición, diversidad y estructura de comunidades de árboles en un gradiente altitudinal de selvas subtropicales de montaña (Serranías de Zapla, Jujuy, Argentina)
Composition, diversity and structure of tree communities along an altitudinal gradient of subtropical forests of mountain (foothills of Zapla, Jujuy, Argentina)
Griet An Erica Cuyckens1*, Lucio Ricardo Malizia2 y Cecilia Blundo3
1 Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET). Centro de Estudios Territoriales Ambientales y Sociales (CETAS). Facultad de Ciencias Agrarias. Universidad Nacional de Jujuy. San Salvador de Jujuy, Argentina. * Autor de correspondencia. grietcuyckens@yahoo.com
2 Centro de Estudios Territoriales Ambientales y Sociales (CETAS). Facultad de Ciencias Agrarias. Universidad Nacional de Jujuy. San Salvador de Jujuy, Argentina.
3 Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET). Instituto de Ecología Regional (IER), Universidad Nacional de Tucumán, Argentina.
Manuscrito recibido el 18 de diciembre de 2014.
Aceptado el 1 de octubre de 2015.
Resumen
Los gradientes altitudinales están asociados con gradientes climáticos que promueven la diversidad de especies. Este estudio describe la composición, diversidad y estructura de comunidades de árboles a lo largo de un gradiente altitudinal en las selvas subtropicales de montaña (Yungas) en las Serranías de Zapla, Provincia de Jujuy, Argentina. Se establecieron tres parcelas de una hectárea cada una; una a 1150 m, una a 1600 m y una a 2050 metros sobre el nivel del mar. A su vez, cada parcela fue subdividida en 25 subparcelas, donde se midieron e identificaron todos los árboles ≥10 cm de diámetro a la altura del pecho. Se compararon la composición específica, número de especies, diversidad, densidad de individuos, área basal y dominancia de individuos por especie entre elevaciones. Se analizó el efecto de la distancia sobre el recambio de especies y se evaluó la relación entre abundancia de individuos por especie y frecuencia de ocurrencia de la especie. Se muestrearon 1375 individuos, pertenecientes a 46 especies. Se encontró un notable recambio en la composición de especies a lo largo del gradiente: solo tres especies estuvieron presentes en las tres elevaciones muestreadas, y la mayoría (29) se encontraron a una sola altitud. La riqueza específica y la diversidad disminuyeron con la altitud, la densidad de individuos no mostró un patrón claro, el área basal presentó un pico a los 1600 m y la dominancia fue más marcada a mayor altitud. La Selva Montana (1150 m) y el Bosque Montano (2050 m) se diferenciaron claramente en términos de composición específica. La similitud de especies se asoció negativamente con la distancia. La abundancia de individuos por especie se asoció positivamente con la frecuencia de ocurrencia de las especies. Este estudio ratifica que los árboles están ubicados en comunidades en pisos ecológicos altitudinales determinados por variables ambientales a nivel regional, y a su vez enfatiza la importancia de los factores locales en determinar la diversidad, composición y estructura de estas comunidades.
Palabras clave: abundancia, área basal, densidad, dominancia, frecuencia, riqueza, similitud, Yungas.
Abstract
Altitudinal gradients are associated with climatic gradients that promote species diversity. This study describes the composition, diversity and structure of trees along an altitudinal gradient in subtropical mountain forests (Yungas) in Serranías de Zapla, Jujuy province, Argentina. Three 1-ha plots at 1150 m, 1600 m and 2050 m above sea level were established. Each plot was subdivided into 25 subplots, where all trees ≥10 cm of diameter at breast height (dbh) were measured and identified as for species; number of species, diversity, and density of individuals, basal area, and dominance of individuals per species among elevations were compared. The effect of distance on species turnover was analyzed, and the relations between abundance of individuals per species and frequency of species occurrence was evaluated. We sampled 1375 individuals belonging to 46 species. A significant turnover in species composition along the gradient was found: only three species were present at all elevations, and most species (29) were found at a single altitude. Species richness and diversity decreased with altitude, density of individuals showed no clear pattern, basal area peaked at 1600 m, and dominance was more pronounced at higher altitudes. Low Montane Forest (1150 m) and High Montane Forest (2050 m) were clearly different in terms of species composition. Species similarity was negatively associated with distance within each altitudinal level. Abundance of individuals per species was positively associated with frequency of species occurrence. This study confirms the community organization of tree species in altitudinal ecological levels determined by environmental variables in Yungas forests, and emphasizes the importance of local factors in determining the diversity, composition and structure of these communities.
Keywords: abundance, basal area, density, dominance, frequency, richness, similarity, Yungas.
Introducción
Los factores que determinan la composición, diversidad y estructura de las comunidades vegetales cambian dependiendo de la escala y el contexto de estudio (Condit et al., 2002). En ambientes con marcados gradientes altitudinales, la elevación suele ser un factor importante que condiciona de modo directo a variables ambientales como temperatura y precipitación e indirectamente a variables poblacionales como crecimiento y mortalidad. Los disturbios (deslizamientos, ganado, fuego) también varían según el nivel altitudinal (Stephenson y Mantgem, 2005; Homeier et al., 2010; Girardin et al., 2010; Blundo et al., 2012; Anderson-Teixeira et al., 2013).
En general, hay una relación inversamente proporcional entre la riqueza de especies y altitud; más recientemente, Rahbek (2005) describió un patrón en forma de "joroba" con riquezas máximas a altitudes intermedias en gradientes que localmente abarcan todos los niveles de altitud. En Las Yungas bolivianas Bach et al. (2003) encontraron una disminución en la diversidad en la comunidad vegetal relacionada no solo con el incremento de la altitud, sino también con los tipos de suelo. Por su parte, Blundo et al. (2012) encontraron una fuerte asociación entre la temperatura y el estrés hídrico (condicionados por la elevación) con la composición de comunidades de árboles en las Yungas argentinas, con una influencia adicional de factores locales (topografía y presencia de ganado) en la composición específica.
Son pocos los estudios que describen en detalle la composición, diversidad y estructura de las selvas subtropicales de montaña (Yungas) en Argentina. Este estudio describe cambios en la composición, diversidad y estructura de las comunidades de árboles en un gradiente de elevación (1000-2000) metros sobre el nivel del mar (snm), en las selvas sub-tropicales de montaña de las Serranías de Zapla, Provincia de Jujuy, noroeste de Argentina y compara los resultados encontrados con los estudios disponibles.
Objetivos
De modo general, este estudio describe cambios en la composición, diversidad y estructura de las comunidades de árboles en un gradiente de elevación (1150, 1600 y 2050) m snm, en las selvas subtropicales de montaña de las Serranías de Zapla, Provincia de Jujuy, noroeste de Argentina.
De modo específico, se plantean las siguientes preguntas:
1) Composición:
a) ¿Cómo cambia la composición de especies con la elevación?
b) ¿Qué similitud de especies existe entre comunidades a distintas elevaciones?
2) Diversidad:
a) ¿Cómo cambian la riqueza y la diversidad de especies con la elevación?
3) Estructura:
a) ¿Varían el área basal y la densidad de individuos por especie con la elevación?
b) ¿Son las especies de árboles más frecuentes también las más abundantes?
c) ¿Cómo cambia la dominancia según el nivel altitudinal?
Materiales y Métodos
Área de estudio
En Argentina, la ecorregión de Yungas forma una angosta franja de aproximadamente 100 km de ancho, que se extiende latitudinalmente desde la frontera con Bolivia hasta la provincia de Catamarca, 700 km al sur (Cabrera, 1976). En esta extensión se reconocen tres sectores latitudinales: norte, centro y sur; y entre estos bloques se encuentran áreas de bosques xerofíticos chaqueños (Brown y Ramadori, 1988). En sentido altitudinal, las Yungas se desarrollan aproximadamente entre los 400 m y los 2300 m snm, donde se reconocen tres pisos altitudinales: Selva Pedemontana (400-700) m snm, Selva Montana (700-1500/1700) m snm y Bosque Montano (1500/1700-2300) m snm (Cabrera, 1976; Grau y Brown, 1993). El clima de Yungas es templado y húmedo, con lluvias principalmente estivales (80%) y heladas durante el invierno (Bianchi, 1981). La Reserva Ecológica Municipal Serranías de Zapla (24°12' S - 65°4' O) se ubica en el Departamento Palpalá, en el sector norte de Yungas en Argentina (Brown y Ramadori, 1988) y abarca los pisos altitudinales de Selva Montana y Bosque Montano.
Trabajo de campo
Se establecieron tres parcelas rectangulares de 1 ha (500 m x 20 m) cada una en la Reserva Ecológica Municipal Serranías de Zapla, a tres altitudes: una en la Selva Montana a 1150 m snm (24° 11' 40.56" S; 65°8'25.26" O); una en "Transición" a 1600 m snm, entre la Selva Montana y el Bosque Montano (24°14'12.96" S; 65°4'30.84" O); y una en el Bosque Montano a 2050 m snm (24°14'5.22" S; 65°3'42.06" O). No es posible localizar una parcela de muestreo en selva pedemontana (400-700) m snm en las Serranías de Zapla ya que esta, por su topografía regional, no incluye áreas a menos de 1000 m de elevación. Incluir un área de pedemonte hubiera requerido alejarse latitudinalmente demasiado (unos 30 km a 40 km). Las parcelas fueron elegidas maximizando el estado de conservación del bosque; sin embargo, las tres parcelas estuvieron sometidas a pastoreo extensivo por ganado vacuno y presentaron evidencias de extracción de leña y madera, aunque en baja intensidad (tocones: 7 en la Selva Montana, 1 en Transición y 4 en el Bosque Montano). Las tres parcelas se ubicaron a corta distancia entre sí (<10 km), en una misma cuenca y con la misma exposición suroeste (235° ± 34°). Cada parcela fue dividida en 25 sub-parcelas cuadradas de 0.04 ha cada una (20 m x 20 m). Esto permitió medir distancias entre sub-parcelas; las distancias entre las parcelas fueron calculadas con base en sus coordenadas geográficas. Se midieron, marcaron e identificaron a nivel de especie todos los árboles con un diámetro a la altura del pecho (DAP; 1.3 m) igual o mayor a 10 cm. La identificación específica se hizo con base en Digilio y Legname (1966) y Legname (1982). Cuando no fue posible la identificación en campo, se colectó una muestra y se identificó en gabinete. Para la nomenclatura taxonómica se siguió a The Plant List (2013).
Análisis de datos
Composición
Se analizó la composición específica por nivel altitudinal, considerando la identidad de especies compartidas y exclusivas. Se evaluó el efecto de la abundancia de individuos sobre el número de especies (riqueza) mediante curvas de rarefacción realizadas con el programa R (R Development CoreTeam, 2012). El tamaño de las parcelas de muestreo (1 ha), que a su vez condiciona en número de individuos muestreados en cada una (Tabla 2), no parece haber afectado marcadamente el número de especies registradas en cada parcela (elevación). Esto se visualiza mediante el empleo de curvas de rarefacción, que llegan a una asíntota alrededor de los 300 individuos para las tres parcelas (Fig 1), lo que permite comparar de modo directo la riqueza de especies entre elevaciones.
Se calculó la diversidad entre parcelas (1 ha) y sub-parcelas (0.04 ha) con el índice de similitud de Sørensen; β = 2c / (2c+α+b), donde α es el número de especies presentes en una muestra, b es el número de especies presentes en una segunda muestra y c es el número de especies en común. El valor de este índice varía de 0 (ninguna especie en común) a 1 (todas las especies compartidas). Se calculó entre parcelas (1 ha) y entre cuadrados (0,04 ha) de una misma parcela para analizarlo en relación a la distancia geográfica dentro de cada parcela tomando como cero el inicio de la parcela. Luego se calculó la disimilitud (1-β) entre cuadrados y se correlacionó con la distancia (Colwell, 2000). Se evaluó la relación entre el la similitud y la distancia geográfica entre sub-parcelas dentro de una misma parcela empleando pruebas de Mantel (Rosenberg y Anderson, 2011).
Diversidad
Se computó el número de especies como riqueza especifica (S) por nivel altitudinal. Se calculó la diversidad α (diversidad local) por parcela mediante el índice de diversidad de Shannon (H) (Ricklefs, 1998). En este índice, la contribución de cada especie se pondera por su abundancia relativa. Mayores valores de H indican mayor diversidad.
Estructura
La estructura del bosque se analizó considerando la densidad absoluta (D) como el número de individuos presentes en 1 ha y se calculó el área basal (AB) por parcela como la suma de todos los DAP. Se calcularon la abundancia por especie, considerando el número de individuos por especie, sumando las tres elevaciones y la frecuencia absoluta (F) como el número de sub-parcelas (0.04 ha) donde aparece una especie, dividido por 75 (número total de sub-parcelas). Se evaluó una posible asociación entre abundancia y frecuencia con una correlación de Spearman, (Analytical Software, 2003). Se calculó el índice de dominancia de Simpson (λ), que manifiesta la probabilidad de que dos individuos tomados al azar de una muestra sean de la misma especie (calculado como: λ= 27n(n-1)/N(N-1), donde N es el número total de individuos de todas las especies y n es el número total de individuos de la especie en consideración) (Moreno, 2001). Al aumentar la dominancia en una comunidad, esta se hace menos diversa porque los individuos están menos equitativamente distribuidos entre especies.
Resultados
Composición
Se muestrearon un total de 1375 árboles >10 cm de DAP, pertenecientes a 46 especies (40 géneros y 30 familias). La mayoría de las especies (63%; 29 especies) se encontraron en un solo piso altitudinal: 20 en la Selva Montana, cuatro en la Transición y cinco en el Bosque Montano. Catorce especies (30.4%) se encontraron en más de una elevación y solamente tres especies (6.5%; Cedrela lilloi, Sambucus nigra subsp. peruviana y Blepharocalyx salicifolius) estuvieron presentes en las tres elevaciones muestreadas. Una sola especie (Solanum trichoneuron) se encontró a 1150 m snm y a 2050 m snm y no en la altitud intermedia (Tabla 1). La similitud de especies, medida como el índice de Sørensen, fue mayor entre la Transición y el Bosque Montano (0.3396), menor entre la Selva y el Bosque Montanos (0.1481) e intermedia entre la Selva Montana y la Transición (0.2817). El recambio de especies y la distancia geográfica entre sub-parcelas de una misma elevación se correlacionaron positivamente (Prueba de Mantel; 1150 m snm: r = 0.1765, p = 0.032; 1600 m snm: r = 0.4330, p = 0.001; 2050 m snm: r = 0.1402, p = 0.038) (Fig 2).
Diversidad
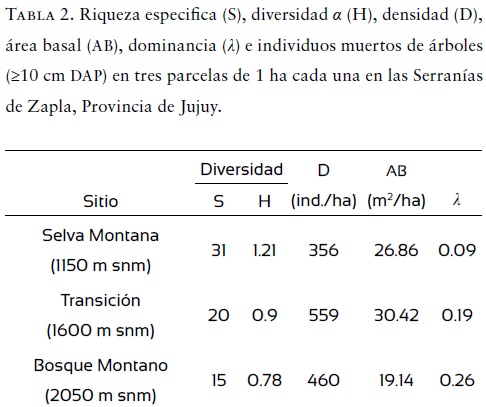
La parcela de Selva Montana presentó una riqueza especifica (S) significativamente mayor de especies que las parcelas ubicadas a elevaciones superiores (X2 = 6.09, p = 0.048) (Tabla 2). La diversidad α (H) disminuyó con la altitud (Tabla 2), pero no fue estadísticamente significativo (X2 = 0.96, p = 0.1).
Estructura
La densidad de individuos por hectárea varió significativamente entre elevaciones (X2 = 44.99, p < 0.0001), siendo máxima en la elevación intermedia (Tabla 2). El área basal fue similar entre las tres parcelas (X2 = 2.61, p = 0.27) (Tabla 2). Unas pocas especies de árboles fueron abundantes, mientras que la mayoría fueron poco abundantes o raras. La especie más abundante fue Allophylus edulis, con 233 individuos, seguida de Prunus tucumanensis, con 211 individuos y Blepharocalyx salicifolius, con 209 individuos. Ocho especies de las 46 encontradas (17.4%) estuvieron representadas por un solo individuo. La abundancia de individuos se correlacionó positivamente con la frecuencia de ocurrencia por especie (Spearman, r = 0.98; p < 0.0001) (Fig. 3). En relación con la dominancia de las especies a distintas elevaciones, esta fue más alta en la parcela de Bosque Montano a 2050 m snm (0.26), seguida por la parcela de Transición a 1600 m snm (0.19) y luego por la de la Selva Montana a 1150 m snm (0.09), pero este patrón no fue significativo (X2 = 0.96, p = 0.08) (Tabla 2).
Discusión
Composición
No se registraron especies exóticas en el estudio, lo que indica el estado de conservación relativamente bueno. Más al sur en las Yungas argentinas (provincia deTucumán) es común encontrar naranjos (Citrus aurantium) y Siempre Verde (Ligustrum sp.) como especies invasoras. Las especies exóticas pueden formar una amenaza en las Yungas (Ayup et al., 2014; Zamora Nasca et al., 2014), pero en el sector estudiado aún no.
Los resultados de este trabajo indican una clara diferencia en términos de composición de especies entre la Selva Montana (1150 m snm) y el Bosque Montano (2050 m snm). La comunidad a 1600 m snm representó una composición intermedia, pero se pareció más al Bosque Montano. En el Parque Provincial Potrero de Yala, distante 36 km de las Serranías de Zapla, la comunidad de árboles a 1600 m snm se parece más a la Selva Montana y el límite entre selva y bosque se ubica a los 1700 m snm (Carranza, 2005). Las diferencias altitudinales en los límites no son algo nuevo: en las Yungas argentinas, Cabrera (1976) establece el límite entre selva y bosque a los 1700 m snm, mientras que Brown (1993) y Brown et al. (2001) lo hacen a los 1500 m snm. Lógicamente, los límites altitudinales de las unidades de vegetación son aproximados y responden a la ubicación regional de las cadenas montañosas y a su altitud total.
Estos resultados sobreestiman la exclusividad de las especies en los distintos pisos altitudinales, ya que solo se muestreó una sola parcela (1 ha) por elevación. Sin embargo, reflejan adecuadamente que las especies tienen un óptimo ecológico a lo largo del gradiente altitudinal. Si se comparan los resultados de este estudio con los de otros estudios (Tabla 3) se observa que la composición de especies de árboles de sitios ubicados a la misma altitud pero en distintas latitudes se parecen más entre sí que las especies del mismo sector latitudinal, pero a otra altitud (Morales et al., 1995; Morales, 1996).
La similitud en un mismo piso altitudinal disminuyó con la distancia (Fig. 2), esto implica que el recambio de especies o diversidad Beta aumentó con la distancia. Este resultado sugiere que incluso pequeñas distancias (<500 m) pueden jugar un rol importante en estructurar la composición de especies a escala local. El recambio es un parámetro en el que inciden tanto la riqueza específica como la densidad relativa por especie. De este modo, al bajar el número de especies, baja la probabilidad de recambio (Malizia, 2004). En este estudio se encontró un menor recambio de especies en la parcela de Bosque Montano (donde se presenta la menor riqueza), mientras que el mayor recambio se dio en la parcela de Transición, la que presenta la mayor densidad de individuos (Fig. 2, Tabla 2).
Diversidad
En el Neotrópico, Gentry (2001) describe un patrón de riqueza y la diversidad de especies que disminuyen al aumentar la altitud. Sin embargo, estudios recientes (Rahbek, 2005) indican como patrón más frecuente una curva en "forma de joroba", con valores máximos a altitudes intermedios. Este estudio aporta evidencia para el primer patrón, mientras Malizia et al. (2012) en un estudio regional en Yungas observaron el segundo. Esto pone de manifiesto la importancia de estudios locales para reconocer estos patrones. Para mamíferos, Ojeda (1999) y Ojeda et al.(2008) encontraron una disminución de riqueza y diversidad de norte a sur. Esto también se da si se compara este estudio con otros en Yungas (Tabla 3), con excepción para un sitio en Toldos.
Estructura
La densidad de individuos no tuvo un patrón claro en relación a la altitud. Carranza (2005) encontró para el Parque Provincial Potrero de Yala una disminución en la densidad de árboles al aumentar la altitud en las laderas con exposición Sur y Este, pero no en las laderas con exposición Norte y Oeste. En cambio, Brown et al. (2001) no observan cambios importantes en los valores de densidad entre un bosque situado a 600 m snm y otro a 1600 m snm. Esto sugiere que la densidad se mantiene relativamente constante entre altitudes en Yungas. En general, el área basal tiende a aumentar con la altitud (Lieberman et al., 1996; Vázquez y Givnish, 1998). Esto no ocurrió en Zapla, probablemente debido a la alta mortalidad de árboles encontrada en la parcela a 2050 m snm (54 árboles muertos).
La abundancia de individuos por especie y la frecuencia de ocurrencia de las especies estuvieron positivamente correlacionadas, sugiriendo que las especies más abundantes tienden a ocupar más sitios, esto quiere decir que las especies no crecen agrupadas. Como excepción, están las especies que están ubicadas por debajo de la línea de correlación, que son las que si tienden a presentar una distribución agregada, como Prunus tucumanensis y Allophylus edulis, que presentó un pico de abundancia a los 1600 m snm (192 individuos), pero no fue censada a los 2050 m snm. Las especies ubicadas por arriba de la línea de correlación son las que están más desagregadas espacialmente. Un ejemplo de esto es Cedrela lilloi, que estuvo presente en todo el gradiente y además en varias sub-parcelas adentro de cada parcela. Otras especies con patrones similares fueron Sambucus nigra subsp. peruviana y Podocarpus parlatorei.
La dominancia (λ) aumentó con la altitud (Tabla 2), siendo máxima en el Bosque Montano, donde la riqueza fue menor pero la densidad de individuos fue intermedia. En general, esto es así en todos los bosques montanos, donde unas pocas especies involucran la mayoría de los individuos (Carranza, 2005). Este patrón de pocas especies abundantes y muchas raras se da en todos los pisos altitudinales: las tres especies más abundantes por piso altitudinal (que varían entre elevaciones) aportan 41% de los individuos a los 1150 m snm, 86% a los 1600 m snm y 76% a los 2050 m snm. Las especies raras (definidas por un solo individuo por parcela) representan 38% a los 1150 m snm, 35% a los 1600 m snm y 20% a los 2050 m snm.
La composición, diversidad y estructura de las comunidades de árboles de las Serranías de Zapla variaron en función de la elevación. Este trabajo comprueba que la altitud impone límites a los rangos de las especies probablemente a través de variables climáticas, ecologías y fisiológicas. Los distintos pisos altitudinales de Zapla se parecen más a sitios ubicados a elevaciones similares aunque a cientos de kilómetros, que entre sí (Tabla 3). A pesar de la marcada influencia de los factores ambientales ligados a la altitud (temperatura y precipitación), que determinan la ocurrencia de bosques similares a elevaciones similares, los factores locales (ej. incendios y extracción maderera) imponen condiciones particulares que a su vez condicionan parámetros del bosque como área basal y densidad de individuo, esto refuerza la importancia de estudios a escala local. El presente estudio es un trabajo de ecología general donde se comparan tres tipos de bosques del extremo norte de Yungas de Argentina, a través de diferentes índices de diversidad y similitud. Si bien se encuentran diferencias estadísticamente significativas entre ellas, es un trabajo acotado a una zona puntual de la estructura de Yungas.
Conclusiones
Este trabajo describe la composición, diversidad y estructura de un bosque en el extremo sur del Sector Norte de las Yungas argentinas. El recambio de especies fue importante a cortas distancias en un mismo piso altitudinal. A pesar de la influencia de los factores regionales, esto refuerza la importancia de estudios a escala local para dilucidar cómo responden las comunidades de árboles a las condiciones locales. Las especies tienden a ser exclusivas de un intervalo altitudinal determinado y luego desaparecen o son escasas en otras elevaciones. Esto refleja que las especies tienen un óptimo ecológico a lo largo del gradiente ambiental.
Los sitios Selva Montana a 1150 m snm y Bosque Montano a 2050 m snm se diferencian claramente entre sí como comunidades diferentes. La comunidad a 1600 m smn presenta una composición específica intermedia, aunque se parece más al bosque montano. Dentro de las parcelas, se encuentra un mayor recambio de especies a medida que aumenta la distancia entre los cuadrados comparados, lo que podría reflejar una limitación en la dispersión de los propágulos. El Bosque Montano estudiado es más homogéneo en términos de composición de especies que la Selva Montana. La riqueza específica y la diversidad de las comunidades arbóreas decrecen con incrementos en la altitud.
La densidad de individuos y el área basal del bosque presentan valores máximos a una altitud intermedia. Hay una dominancia más marcada a mayor altitud: se encuentran menos especies por hectárea, sin diferencias significativas entre los valores de área basal. La comunidad estudiada es una comunidad boscosa con pocas especies abundantes y muchas especies raras. De las 46 especies encontradas 17.4% (8) estuvo representada por un solo individuo y menos de 10% de las especies (5) cuenta con más de 100 individuos. Hay una correlación positiva entre la abundancia y la frecuencia de ocurrencia de las especies, sugiriendo que las especies más abundantes son aquellas que pueden ocupar una diversidad mayor de sitios. Este estudio indica la importancia de contar con gradientes altitudinales completos para la conservación de las selvas subtropicales de montaña del norte de la Argentina y que cada altitud tiene sus especies exclusivas.
Reconocimientos
Agradecemos a R. Guevara por la ayuda con el trabajo de campo y a la Fundación ProYungas por su apoyo logístico. La Fundación Nacional de Ciencias (EEUU) financió el establecimiento de las parcelas. Este trabajo se realizó en el marco del trabajo de tesis de licenciatura de la primera autora en la carrera de biología de la Facultad de Ciencias Agrarias de la Universidad Nacional de Jujuy.
Referencias
Analytical Software. 2003. Statistix for Windows. Analytical Software. [ Links ]
Anderson-Teixeira, K.J., A.D. Miller, J.E. Mohan, T.W. Hudiburg, B.D. Duval y E.H. DeLucia. 2013. Altered dynamics of forest recovery under a changing climate. Global Change Biology 19(7):2001-2021. [ Links ]
Ayup, M.M., L. Montti, R. Aragón y H.R. Grau. 2014. Invasion of Ligustrumlucidum (Oleaceae) in the southern Yungas: Changes in habitat properties and decline in bird diversity. ActaOecologica 54:72-81. [ Links ]
Bach, K., M. Schawe, S. Beck, G. Gerold, S.R. Gradstein y M. Moraes R. 2003. Vegetación, suelos y clima en los diferentes pisos altitudinales de un bosque montano de Yungas, Bolivia: Primeros resultados. Ecología en Bolivia 38(1):3-15. [ Links ]
Bianchi, A.R. 1981. Las precipitaciones del noroeste argentino. INTA. Salta. [ Links ]
Blundo, C., L.R. Malizia, J.G. Blake y A.D. Brown. 2012. Tree species distribution in Andean forests: influence of regional and local factors. Journal of Tropical Ecology 28(1):83-95. [ Links ]
Brown, A.D. 1993. Fitogeografía y conservación de las Selvas de Montaña del Noroeste de Argentina. In: S. P. Churchill, H. Balslev, E. Forero y J.L. Luteyn, eds. Biodiversity and conservation of neotropicalmontane forests.The New York Botanical Garden. Nueva York. p:663-672. [ Links ]
Brown, A. D. y E. Ramadori. 1988. Patrón de distribución, diversidad y características ecológicas de las especies arbóreas de las selvas y bosques montanos del N.O. de la Argentina. In: Actas VI Congreso Forestal Argentino. Santiago del Estero. p:177-181. [ Links ]
Brown, A. D., S. Chalukian y L. Malmierca. 1985. Estudio florístico estructural de un sector de la selva semidecidua del Noroeste argentino. I. Composición florística, densidad y diversidad. Darwiniana 26(1):27-41. [ Links ]
Brown, A. D., H. R. Grau, L. R. Malizia y A. Grau. 2001. Argentina. In: M. Kapelle y A.D. Brown, eds.Bosques nublados del Neotrópico. San José. INBio. p:623-659. [ Links ]
Cabrera, A. 1976. Regiones fitogeográficas de la República Argentina. Enciclopedia de Agricultura, Jardinería y Fruticultura. Vol. 2. Acme. Buenos Aires. [ Links ]
Carranza, A.V. 2005. Bosque y selva montanos en Yala, Jujuy -Argentina. Universidad Nacional de Jujuy, EdiUnju. Jujuy. [ Links ]
Colwell, R.K. 2000. EstimateS: statistical estimation of species richness and shared species from samples. ver 8.2.0. [ Links ]
Condit, R., N. Pitman, E.G. Leigh Jr, J. Chave, J. Terborgh, R.B. Foster, P. Núñez, S. Aguilar, R. Valencia, G. Villa, H.C. Muller-Landau, E. Losos y S.P. Hubbell. 2002. Beta-diversity in tropical forest trees. Science 295(1):666-669. [ Links ]
Digilio, A. P. y P. Legname. 1966. Los árboles indígenas de la provincia de Tucumán. Opera Lilloana 15:1-107. [ Links ]
Gentry, A. H. 2001. Patrones de diversidad y composición florística en los bosques de las montañas neotropicales. In: M. Kapelle y A.D. Brown, eds. Bosques nublados del Neotrópico. INBio. San José. p:85-123. [ Links ]
Girardin, C.A.J., Y. Malhi, L.E.O.C. Aragao, M. Mamani, W. Huaraca Huasco, L. Durand, K.J. Feeley, J. Rapp, J.E. Silva-Espejo, M. Silman, N. Salinas y R.J. Whittaker. 2010. Net primary productivity allocation and cycling of carbon along a tropical forest elevational transect in the Peruvian Andes. Global Change Biology 16(12):3176-3192. [ Links ]
Grau, H. R. y A.D. Brown. 1993. Patterns of tree species diversity along latitudinal and altitudinal gradients in the Argentinean subtropical montane forests. In: S.P. Churchill, H. Balslev, E. Forero y J.L. Luteyn, eds. Biodiversity and conservation of neotropicalmontane forests. The New York Botanical Garden. Nueva York. p:295-300. [ Links ]
Homeier, J., S.W. Breckle, S. Günter, R.T. Rollenbeck y C. Leuschner. 2010. Tree Diversity, Forest Structure and Productivity along Altitudinal and Topographical Gradients in a Species-Rich Ecuadorian Montane Rain Forest: Ecuadorian Montane Forest Diversity and Structure. Biotropica 42(2):140-148. [ Links ]
Legname, P. 1982. Los árboles indígenas del noroeste argentino. Opera Lilloana 34:1-226. [ Links ]
Lieberman, D., M. Lieberman, R. Peralta y G.S. Hartshorn. 1996. Tropical forest structure and composition on a large-scale altitudinal gradient in Costa Rica. Journal of Ecology 84(1):137-152. [ Links ]
Malizia, L.R. 2004. Diversity and distribution of tree species in subtropical Andean forests. St. Louis, Missouri: University of Missouri- St. Louis Department of Biology Program in Ecology, Evolution and Systematics. [ Links ]
Malizia, L.R., S. Pacheco, C. Blundo y A.D. Brown. 2012. Caracterización altitudinal, uso y conservación de las Yungas Subtropicales de Argentina. Ecosistemas 21(1-2):53-73. [ Links ]
Mármol, L.A. 1992. Uso y conservación de ecosistemas en montañas subtropicales del Noroeste argentino. Salta: Consejo de Investigación. Universidad Nacional de Salta. [ Links ]
Morales, J. M. 1996. Comunidades arbóreas no saturadas en las Yungas de Argentina. Ecología Austral 6:17-22. [ Links ]
Morales, J. M. y A.D. Brown. 1996. Bosques montanos con diferente intensidad de explotación. Bosques y Desarrollo 17:51-52. [ Links ]
Morales, J. M., M. Sirimbra y A.D. Brown. 1995. Riqueza de árboles en las Yungas argentinas. In: A.D. Brown y H.R. Grau, eds. Investigación, conservación y desarrollo en selvas subtropicales de montaña. Proyecto de Desarrollo Forestal - L.I.E.Y. Tucumán. p:163-174. [ Links ]
Moreno, C.E. 2001. Métodos para medir la biodiversidad. Vol. 1. M&T-Manuales y Tesis SEA. Zaragoza. [ Links ]
Ojeda, R.A. 1999. Biodiversidad y conservación de mamíferos de la interfase tropical-templada de Argentina. In: S.D. Matteucci, O.T. Solbrig, J. Morello y G. Halffter, eds.Biodiversidad y uso de la tierra: conceptos y ejemplos de Latinoamérica. EUDEBA-UNESCO Colección CEA. Buenos Aires. p:443-462. [ Links ]
Ojeda, R.A., R.M. Barquez, J. Stadler y R. Brandl. 2008. Decline of Mammal Species Diversity Along the Yungas Forest of Argentina. Biotropica 40(4): 515-521. [ Links ]
Rahbek, C. 2005. The role of spatial scale and the perception of large-scale species-richness patterns. Ecology Letters 8(2):224-239. [ Links ]
R Development Core Team. 2012. The R Project for Statistical Computing (version 2.15.2). Viena, Austria. [ Links ]
Ricklefs, R. E. 1998. Invitación a la ecología. La economía e la naturaleza. 4th ed. Buenos Aires, Argentina: Panamericanana. [ Links ]
Rosenberg, M. S. y C.D. Anderson. 2011. PASSaGE: pattern analysis, spatial statistics and geographic exegesis. Version 2.Methods in Ecology and Evolution 2(3):229-232. [ Links ]
Stephenson, N.L. y P.J. Mantgem. 2005. Forest turnover rates follow global and regional patterns of productivity: Patterns in forest turnover rates. Ecology Letters 8:(5)524-531. [ Links ]
The Plant List. 2013. version 1.1. [ Links ]
Vazquez, G. y T.J. Givnish. 1998. Altitudinal gradients in tropical forest composition, structure and diversity in the Sierrra de Manantlán. Journal of ecology 86(1):999-1020. [ Links ]
Zamora Nasca, L., L. Montti, R. Grau y L. Paolini. 2014. Efectos de la invasión del ligustro, Ligustrumlucidum, en la dinámica hídrica de las Yungas del noroeste Argentino. Bosque (Valdivia) 35(2):195-205. [ Links ]
Nota
Este documento se debe citar como: Cuyckens, G.A.E., L.R. Malizia y C. Blundo. 2015. Composición, diversidad y estructura de comunidades de árboles en un gradiente altitudinal de selvas subtropicales de montaña (Serranías de Zapla, Jujuy, Argentina). Madera y Bosques 21(3):137-148.

