










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkMadera y bosques
versión On-line ISSN 2448-7597versión impresa ISSN 1405-0471
Madera bosques vol.14 no.1 Xalapa mar. 2008
Artículo de investigación
Dinámica sucesional del encinar de la sierra de Álvarez, San Luis Potosí, México
Succession dynamics of the sierra de Alvarez oak forests in San Luis Potosí, México
Pedro Castillo Lara1, Jorge Alberto Flores Cano1, Juan Rogelio Aguirre Rivera2 y Richard I. Yeaton H.2
1 Programa Multidisciplinario de Posgrado en Ciencias Ambientales (PMPCA), Universidad Autónoma de San Luis Potosí, Área de Recursos Naturales. Altair No. 200. Fracc. Del Llano. C.P. 78377. San Luis Potosí, S.L.P. pcastillol57@yahoo.com.mx; florescano.bq@gmail.com.
2 Instituto de Investigación de Zonas Desérticas de la Universidad Autónoma de San Luis Potosí. Altair No. 200. Fracc. Del Llano. C.P. 78377.
Manuscrito recibido el 15 de diciembre de 2006.
Aceptado el 19 de junio de 2007
RESUMEN
El efecto de la competencia interespecífica sobre la dinámica de los bosques templados en México ha sido poco estudiado. En este trabajo, por medio de técnicas de inferencia fuerte, se documenta la dinámica de un bosque de encino de la Sierra Madre Oriental. En particular, se proponen métodos para determinar cuáles especies son importantes en la dinámica de una comunidad arbórea, la secuencia de establecimiento de esas especies, el remplazo competitivo de una especie por otra y, finalmente, cómo la biología de las especies afecta su estado sucesional. Los resultados presentan una secuencia de especies de encinos rojos (Quercus coccolobifolia, Q. crassifolia y Q. affinis) remplazándose sucesionalmente, y un encino blanco (Q. obtusata) que se presenta como subdominante en la etapa sucesional tardía dominada por Q. affinis.
Palabras clave: Bosques de encino, inferencia fuerte, remplazo competitivo, sucesión.
ABSTRACT
The effect of interspecific competition on the dynamics of temperate forests in Mexico has rarely been investigated. In this study, the dynamics of an oak forest in the Sierra Madre Oriental are described using techniques involving strong inference. In particular, methods are presented to determine which species are important in the tree community, the sequence of establishment of these species, the competitive replacement of one species by another and, finally, how each species' biology affects their succesional status. The results show a sequence of three species of red oaks (Quercuscoccolobifolia, Q. crassifolia and Q. affinis) replacing one another over time with a fourth species, a white oak Q. obtusata, functioning as a subdominant to Q. affinis in the late succesional stage of the forest.
Key words: Oak forests, strong inference, competitive replacement, succession.
INTRODUCCIÓN
Los tipos de vegetación en México han sido descritos en detalle (Flores et al., 1971; Rzedowski, 1978). Rzedowski (1978) compendió lo conocido sobre la ecología general de dichos tipos de vegetación y sugiere que el siguiente paso en el estudio de las comunidades vegetales sería lograr entender su dinámica, con énfasis en aquellas especies más importantes involucradas en dichos procesos. Una forma de abordar esta dinámica es a través del concepto del remplazo espacial sobre el tiempo (Watt, 1947). Al respecto, uno de los primeros estudios en México fue realizado por Yeaton y Romero-Manzanares (1986) en el matorral xerófilo del sur del desierto chihuahuense. Ellos estudiaron el remplazo, a través del tiempo, de Acacia shaffneri por Opuntia streptacantha, para lo cual utilizaron técnicas de inferencia fuerte, propuestas inicialmente por Platt (1964).
La dinámica de los bosques de dosel cerrado es difícil de apreciar, pues a simple vista dan la apariencia de ser estáticos (Shugart, 1984). Sin embargo, en su interior ocurren cambios continuos debidos a las variaciones en el ambiente físico y biótico. Estos cambios modifican su estructura y composición específica a través del tiempo. Whitmore (1978) señala que en la dinámica de los bosques templados pueden identificarse tres fases: a) la perturbación, b) la invasión secuenciada de varias especies, y c) la madurez, última etapa en la sucesión. En cada fase pueden identificarse especies características, cuyos atributos distintivos se refieren a sus formas de dispersión, colonización y crecimiento en condiciones de competencia por la luz y otros recursos (Whitmore, 1978; Martínez, 1985).
La fase de perturbación se manifiesta con la apertura de claros de diferente tamaño y forma, en dependencia del tipo, amplitud e intensidad del disturbio. Estos claros son colonizados primeramente por especies intolerantes a la sombra, tasas fotosintéticas y puntos de saturación elevados en presencia de luz y, como consecuencia, una superficie fotosintética mayor (Horn, 1974; Bidwell, 1974; Bazzaz, 1998). En la fase de invasión, las condiciones del sotobosque cambian de tal forma que resulta inadecuado para las colonizadoras, pero favorable para el arribo de otras especies mejor adaptadas a las nuevas condiciones (Connell y Slatyer, 1977). Las especies dominantes en esta etapa presentan como características principales mayor tolerancia a la sombra, alta capacidad para crecer y competir bajo condiciones de poca luz, mayor eficiencia fotosintética bajo la sombra, puntos de compensación bajos, menor gasto energético para la respiración y, como consecuencia, menor superficie foliar (Horn, 1974; Bidwell, 1974; Bazzaz, 1998).
Finalmente, durante la fase de madurez, las comunidades vegetales adquieren una estructura y composición complejas y relativamente estables (Perry, 1994). Sin embargo, aun en esta fase los cambios continúan, al modificarse la abundancia relativa de las especies (Forcier, 1975). Es en esta etapa cuando la competencia entre individuos promueve el proceso de remplazo. Aunque entre los ecólogos aún hay desacuerdo sobre la importancia de la competencia en la organización de las comunidades vegetales, al respecto existen evidencias sólidas sobre su relevancia (Callaway y Walter, 1997; Flores y Yeaton, 2000).
El papel de la competencia en la dinámica de los bosques de México es muy poco conocido. Los únicos estudios de sucesión se han realizado en bosques dominados por coníferas (Yeaton et al., 1987; González-Espinosa et al., 1991; Sánchez-Velásquez y García-Moya, 1993). Se carece de estudios de sucesión en bosques dominados por encinos en México, y estudios del remplazo espacial a través del tiempo entre especies de encino se desconocen en la literatura científica en nivel mundial.
OBJETIVOS
El objetivo principal de este estudio es documentar la dinámica de la sucesión tardía en bosques densos de encino, por medio de técnicas de inferencia fuerte (Platt, 1964), para responder a las siguientes preguntas: ¿Cómo reconocer cuáles especies son más importantes en la dinámica de una asociación arbórea? ¿Cuáles especies colonizan primero las áreas abiertas? ¿Existe algún patrón de remplazo entre las especies que componen los bosques de encino? ¿Cómo una especie desplaza a otra especie de la etapa previa? ¿Existe una especie que controla la fase final de la sucesión en los bosques de encino?
MÉTODOS
Área de estudio
Este estudio se realizó en la vecindad del paraje conocido como El Carrizal (21° 58' N, 100° 34' O) en la sierra de Álvarez, San Luis Potosí. Esta sierra forma parte de la Sierra Madre Oriental y, en su sección sur, está formada por montañas altas con laderas abruptas y convexas de origen volcánico (INEGI, 2002). Los suelos predominantes en la región son litosoles de textura media (Arriaga et al., 2000). El clima de la región está clasificado como seco templado, con una temperatura promedio anual de 16 °C (García, 1988). La amplitud diaria de temperatura del mes más frío (enero) oscila entre -3 y 18 °C y la media del mes más caliente (mayo) es superior a 18 °C. La precipitación promedio anual es de 566 mm, de la cual el 92% ocurre en los meses de mayo a octubre y el resto en los meses de noviembre a abril (García, 1988). Los promedios de temperatura y precipitación fueron calculados para un periodo de 18 años, no especificado por García (1988) y corresponden a la estación más cercana al área de estudio (La Salitrera, S.L.P.).
En la sierra de Álvarez existen cinco tipos de vegetación principales, de los cuales el bosque de encino es el más extenso (Rzedowski, 1961; Arriaga et al., 2000). El sitio del presente estudio es una ladera con exposición norte, donde las condiciones de humedad y temperatura han mantenido una comunidad de árboles con dosel cerrado, dominado por varias especies del género Quercus. Este sitio se encuentra a una altitud sobre el nivel medio del mar de 2 166 m y presenta pendientes de 20 hasta 35°. La ubicación y topografía del sitio han dificultado el aprovechamiento del arbolado, por lo cual la comunidad de encinos se mantiene prácticamente sin disturbio. Las áreas relativamente más planas, localizadas en la parte baja de la ladera de estudio, fueron desmontadas y utilizadas previamente en la producción de maíz, frijol y chícharo, y luego abandonadas hace alrededor de cincuenta años. Actualmente, estas áreas son mantenidas abiertas debido al pastoreo ligero de bovinos y equinos, pero están en el proceso de repoblación de especies leñosas, particularmente de especies de encinos, existentes en el bosque inmediato.
Trabajo de campo
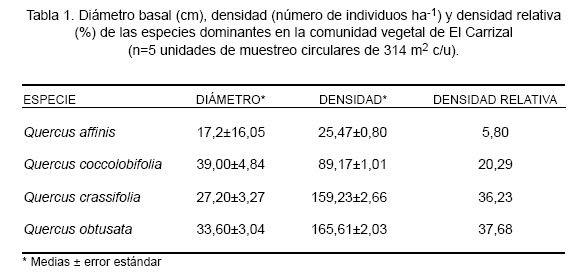
La composición de especies y la abundancia relativa del componente arbóreo, fueron estimadas en cinco parcelas circulares de 20 metros de diámetro (314 m2). Estas parcelas fueron espaciadas en los aproximadamente 400 metros de la ladera en el área de estudio a intervalos de 20 m, a partir de aproximadamente 100 m del campo abierto en la base de la ladera, para terminar aproximadamente a 100 m de la cúspide. Se Dinámica del encinar de la sierra de Álvarez, San Luis Potosí, México registró el número de individuos de cada especie arbórea con diámetro en la base del tronco mayor que 2,5 cm. Arbutus xalapensis, Buddleia cordata, Carya ovata, Crataegus parryana, Junlperus flaccida, Pinus arizonica y P. teocote, especies con menos que el 5% de densidad en el total de las muestras, se consideraron poco importantes en la dinámica de la comunidad y fueron excluidas del estudio. Las medias y errores estándar fueron calculados para las densidades (número de individuos por hectárea) y diámetros de los troncos, para las especies registradas. Aunado a ello, las densidades relativas fueron calculadas, al dividir el promedio de la densidad media de cada especie entre la densidad media total. Estos datos fueron usados para determinar las especies dominantes en el sitio de estudio.
La invasión relativa de las especies de encino fue estudiada en cinco parcelas alargadas (franjas) de 30 x 5 metros, dispuestas en secuencia continua y longitudinalmente paralelas al borde del bosque; la primera de ellas quedó bajo el dosel y las otras cuatro en el campo abierto adyacente. El número de individuos menores que un metro de altura, para cada especie de encino, fue registrado en cada franja. Este proceso fue repetido en cinco sitios diferentes, adyacentes al sitio de estudio. Estos datos proporcionaron información tanto de la repoblación natural en las áreas abiertas, como del patrón de remplazo a través de una secuencia sucesional. El número de individuos de cada especie de encino registrado en estas franjas, fue sumado y analizado estadísticamente con la prueba de X2 para k muestras independientes, donde los encabezados de las filas fueron las cuatro especies de encino y los encabezados de las columnas fueron las cinco franjas de 30 x 5 m, la de bajo el dosel y las cuatro ubicadas en los claros (Siegel y Castellan, 1988).
La técnica de simetría de doseles (Flores y Yeaton, 2000) se usó para comprobar la existencia de patrones de remplazo entre las especies de encino que componen el bosque. Esta técnica, hasta ahora utilizada para inferir el patrón de sucesión entre arbustos y árboles en los matorrales xerófilos del Altiplano Mexicano, mide la capacidad de una especie para invadir el espacio de otra especie. El efecto de la invasión del área del dosel causa un crecimiento asimétrico en la segunda especie y, eventualmente, su remplazo. Se buscaron al azar alrededor de 25 pares de árboles de las cuatro especies de Quercus, cuyos doseles estuvieran solapados. Algunas combinaciones de pares no ocurrieron; sólo 19 pares de Q. obtusata con Q. affinis y 17 pares de Quercus coccolobifolia con Q. crassifolia fueron localizados en el sitio de estudio. Los datos adicionales para complementar suficientes pares de las dos últimas especies, fueron tomados en un sitio relativamente similar, localizado a la misma elevación, a 1,5 km al este del sitio de estudio. Esta área se encuentra en la parte baja de la ladera y en una etapa sucesional temprana debido a la extracción selectiva de individuos adultos de Q. affinis y Q. crassifolia. En el plano formado por los troncos del par de encinos, se midió la distancia entre ambos individuos, el diámetro máximo del dosel de cada individuo, y el radio máximo de sus doseles sin solapo. Para cada individuo, el diámetro máximo del dosel se dividió entre el radio del dosel sin solapo con la otra especie. De acuerdo con Flores y Yeaton (2000), para una especie los valores medios de esta razón estadísticamente menores que 2,0 indican su asimetría por inhibición y eventual remplazo por su competidora, la cual para ello deberá ser estadísticamente simétrica (2,0) o asimétrica inhibidora (>2,0). Los valores de simetría para los pares de especies fueron contrastados estadísticamente a través de la prueba de U de Wilcoxon-Mann-Whitney (Siegel y Castellan, 1988).
Durante el registro de datos para el estudio de la simetría de doseles, se observó que en dos de las combinaciones de pares estudiados, Quercus crassifolia-Q. coccolobifolia y Q. affinis-Q. crassifolia, muchos de los individuos con crecimiento asimétrico presentaban su ápice principal roto. Para evaluar la importancia de esta observación, se cuantificó la condición del tallo principal de individuos de Q. coccolobifolia y Q. crassifolia, creciendo bajo el dosel de Q. affinis, la especie de encino aparentemente dominante. Así, se registró si sus troncos principales estaban rotos o intactos. Estos datos fueron contrastados estadísticamente con una prueba de bondad de ajuste de x2 para cada especie (Siegel y Castellan, 1988).
Con un taladro de Pressler se tomaron muestras del crecimiento radial del tronco de diez individuos de Quercus crassifolia apareados con individuos de Q. affinis y con doseles asimétricos. Esto se realizó con el fin de explorar si el remplazo se asocia con alteraciones en el tronco de la especie suprimida. Antes de su muestreo, se registró el diámetro de cada individuo de Q. crassifolia. Posteriormente, con el taladro se tomó una muestra de su madera, atravesando completamente el tronco del árbol, a una altura del suelo aproximada de 1 m y en el plano definido por los troncos de la pareja. A continuación, las longitudes de las dos secciones radiales de estas muestras fueron medidas a partir del punto de crecimiento inicial del árbol, tanto en la dirección del solapo de doseles como hacia el extremo opuesto. Las diferencias entre las dos secciones radiales, fueron calculadas para cada individuo mediante sustracción del radio dirigido hacia el individuo apareado de Q. affinis, al radio en la dirección opuesta. Estas diferencias en el crecimiento radial fueron calculadas como porcentaje del diámetro medio del tronco. Los valores de las diferencias (positivos o negativos) fueron contrastados estadísticamente a través de una prueba binomial para una muestra (Siegel y Castellan, 1988).
RESULTADOS
El componente arbóreo de la comunidad vegetal de El Carrizal está compuesto por nueve especies arbóreas, de las cuales Quercus crassifolia, Q. coccolobifolia, Q. obtusata y Q. affinis presentaron el mayor porcentaje de ocurrencia (Tabla 1). De los 177 individuos registrados en las cinco unidades de muestreo, dichas especies representaron el 89%, lo cual indica que son las más importantes en la dinámica vegetal de esta comunidad.
La colonización de las áreas abiertas y abandonadas, adyacentes al borde del bosque, por las cuatro especies destacadas de encino, muestra tendencias decrecientes a partir del dosel del bosque (Tabla 2). La mayor abundancia de plántulas y juveniles de Quercus affinis se registró en forma altamente significativa bajo el dosel del bosque y en el borde del mismo, en comparación con los valores esperados (%2 = 36,7, p < 0,001), y fue significativamente menor con respecto a los valores esperados en las áreas abiertas (X2 = 49,1, p < 0,001 en el borde del dosel del bosque; y X2 = 31,2, p < 0,01 en las franjas localizadas entre 5 y 10 m del borde del bosque).

En contraste, las plántulas de Q. coccolobifolia fueron significativamente menos abundantes bajo el dosel del bosque, con respecto al valor esperado (X2 = 92,3, p < 0,001), y significativamente más abundantes que el valor esperado en todas las áreas abiertas [prueba de X2 en un rango desde 165,3 (p < 0,001) en la franja de 5 a 10 m,hasta 28,7 (p < 0,05) en la franja localizada de 15 a 20 m de distancia del borde del bosque). La densidad de plántulas de Quercus obtusata declinó de la franja bajo el dosel del bosque hacia las franjas de las áreas abiertas, pero sin que estas diferencias fueran estadísticamente significativas en relación con el valor esperado y de acuerdo con el estadístico utilizado. El número de individuos registrado de Q. crassifolia fue muy pequeño como para derivar alguna conclusión probable acerca de su recolonización en las áreas abiertas próximas al borde del bosque.
De los seis pares posibles de las cuatro especies de encino para el análisis de simetría de doseles, sólo tres de ellas ocurrieron en el sitio de estudio (Tabla 3). Faltaron los pares posibles de Quercus crassifolia con Q. obtusata y de Q. coccolobifolia con Q. affinis. Así, Quercus affinis se localizó apareado con Q. crassifolia y Q. obtusata, y ambas especies mostraron asimetría significativa de su dosel (Tabla 3). Al mismo tiempo, los doseles de Q. coccolobifolia se registraron significativamente distorsionados al estar emparejados con los de Q. crassifolia (Tabla 3).

En relación con la integridad del tallo de los individuos de las especies apareadas con Q. affinis, los 17 individuos de Q. coccolobifolia localizados en el sitio de estudio, presentaron su tronco principal roto (Tabla 4, x2 = 9,7, p < 0,01). De modo similar, el 72% de los individuos de Q. crassifolia habían sufrido fractura de su tallo principal (Tabla 4, x2 = 5,7, P < 0,02).

La diferencia media relativa entre las dos secciones radiales de los diez troncos evaluados muestra que el crecimiento radial del tallo de Quercus crassifolia, fue estadísticamente mayor en la dirección opuesta al individuo apareado de Q. affinis (X= 10,0 ± 2,1%; p< 0,01).
DISCUSIÓN
Los resultados del estudio de densidad absoluta y relativa indican que la comunidad vegetal de El Carrizal está compuesta por nueve especies arbóreas, pero sólo cuatro de ellas son importantes en la dinámica de este sistema. Rzedowski (1961) menciona que dos de las especies estudiadas (Q. crassifolia y Q. coccolobifolia) son las dominantes fisonómicas en los bosques de encino-pino de la sierra de Álvarez. En bosques templados maduros, sin disturbio aparente, la composición arbórea es mínima, pues llega a ser monoespecífica (García-Arévalo et al., 2004), pero otras veces presenta de cuatro hasta nueve especies (Quintana-Ascencio y González-Espinosa, 1993). De acuerdo con la densidad de las especies dominantes, pareciera que la comunidad estudiada se encuentra en una fase sucesional de intermedia a tardía, debido al predominio de individuos de especies que pueden ser consideradas de la fase intermedia (Q. crassifolia y, posiblemente, Q. obtusata), menos individuos de la especie reconocida como colonizadora (Q. coccolobifolia), y muy pocos individuos de Q. affinis, especie tardía en este gradiente, como se discute enseguida.
De las tres secciones del género Quercus, en México sólo dos son abundantes en especies, Quercus o encinos blancos (81 especies) y Lobatae o encinos rojos (76 especies) (Valencia, 2004). De las especies dominantes en el área de estudio, Q. coccolobifolia, Q. crassifolia y Q. affinis pertenecen a la sección Lobatae, y Q. obtusata a la sección Quercus. Estas dos secciones son evolutivamente diferentes. Así, con base en datos ecológicos y de distribución, se supone que los encinos rojos son menos tolerantes a condiciones xéricas (Nixon, 1984). Debido a lo anterior, en México y Estados Unidos de Norteamérica hay una mayor presencia de encinos blancos en los sistemas montañosos que limitan con los desiertos (Nixon, 1984, 1993). Los encinos blancos producen bellotas más grandes que los rojos, lo cual se relaciona con el contenido de humedad y por lo tanto de reservas para germinar en condiciones de relativa sequedad (Zavala, 2004). Los encinos rojos por su parte, en general crecen donde las condiciones de humedad son mejores. Algunas excepciones son Q . affinis y Q. eduardii del bosque de Pinus cembroides en Nuevo León (Valdez y Ag u i I ar, 1983), Q. coahuilensis en el matorral submontano, y Q. coccolobifolia asociado a terrenos pedregosos con exposición sur en Aguascalientes (De la Cerda, 1989). En el área de estudio, Q. affinis alcanza alturas de hasta 25 m y sólo se encuentra en cañadas y laderas expuestas al norte (García et al., 1999). La escasa presencia de Q. obtusata (encino blanco) en el área de estudio (2 166 m), parece deberse a que, aunque su distribución altitudinal es de 680 a 2 800 m (Valencia, 2004), su mayor abundancia se registra en altitudes intermedias.
El tamaño de la bellota está correlacionado con la facilidad de la planta para establecerse y crecer bajo el dosel del bosque (Beon y Bartsch, 2003). Buckley et al. (1998) mencionan que entre las características de las especies pioneras está el porte bajo y la semilla pequeña, con mayor facilidad de dispersión; mientras que las especies consideradas como dominantes en un bosque maduro, o tardías en la secuencia sucesional, producen frutos con dispersión limitada, pero con energía suficiente para sustentar el crecimiento de las plántulas bajo un dosel establecido de antemano ( M i nckl er, 1980; Jackson, 1984). En efecto, de las cuatro especies estudiadas, Q. coccolobifolia es de porte bajo (6 a 15 m), produce la bellota más pequeña (de 0,8 a 1,0 cm de largo) (García y Aguirre, en preparación), lo cual le facilita colonizar las áreas abiertas. En cambio, Q. affinis posee una bellota más grande, cuyo tamaño fluctúa entre 0,9 y 1,3 cm de largo, lo cual dificulta su transporte a mayores distancias por los depredadores.
El tamaño de la bellota y su relación con los depredadores es también importante en la dispersión y recolonización de las áreas abiertas. La dispersión de frutos por aves y pequeños mamíferos es de gran importancia para el éxito de la repoblación de los encinos de bosques maduros, debido principalmente al almacenamiento eficiente de bellotas (Mellanby, 1968). Los vertebrados (aves y mamíferos) se consideran los dispersores primarios de bellotas (Barnett, 1977; Boucher, 1981; Darley-Hill y Jhonson, 1981). Los roedores pequeños (Peromyscus spp. y Sigmodon spp.) transportan las bellotas entre 10 y 30 m del centro de provisión (Sork, 1984; Bonfil y Soberón, 1999); las ardillas (Sciurus spp. y Sper -mophillus spp.) las mueven hasta unos cientos de metros, y algunas aves (Aphelocoma spp., Corvus sp., Cyanocitta sp., Picoides spp., Melanerpes spp.) las desplazan hasta 4 o 5 km (Darley-Hill y Jhonson, 1981; Zavala, 1995; Zavala y García, 1996). Así, la bellota más pequeña de Quercus coccolobifolia representa menor problema para ser transportada hacia las áreas abiertas, a distancias relativamente grandes de la fuente.
El tamaño de la hoja y su relación con los procesos de la fotosíntesis y respiración, es otro atributo que tiene que ver con la adaptabilidad a diferentes niveles de luz o sombra. Así, es probable que Quercus coccolobifolia requiera de tasas fotosintéticas elevadas desde las primeras etapas de su desarrollo, debido a que su bellota más pequeña contiene menos reservas, lo cual se compensaría con una mayor superficie foliar. Las hojas de Q. coccolobifolia son de 10 a 19 cm de longitud y de 7 a 16 cm de anchura (de la Cerda, 1989; Bello y Labat, 1987; Zavala, 2003; García y Aguirre, en preparación). Por el contrario, Q. affinis posee hojas de 4 a 9 cm de longitud y de 2 a 3 cm de anchura (García y Aguirre, en preparación). Asimismo, el mayor contenido de reservas en las bellotas y el menor gasto energético para la respiración bajo la sombra, podrían conferirle a Q. affinis una mayor eficiencia fotosintética con menor superficie foliar.
Las características biológicas de las especies estudiadas y los resultados de este trabajo concuerdan con lo señalado por Horn (1974), Bidwell (1974) y Bazzaz (1998), en el sentido de que las especies colonizadoras, además de ser intolerantes a la sombra, poseen hojas más grandes. Mientras que las propias de los estados avanzados en la sucesión, son más tolerantes a la sombra y presentan menor superficie foliar. Sin embargo, Buckley et al. (1998) afirman que en general las especies propias de lugares abiertos y con altas tasas de luminosidad, poseen hojas pequeñas y gruesas, mientras que las especies propias de lugares sombreados y de mayor humedad, presentan hojas grandes y delgadas, posiblemente porque se están refiriendo a las especies correspondientes a las comunidades maduras de esos dos ambientes. Rzedowski (1978) menciona que si bien en general los encinares con especies de hoja pequeña se distribuyen en áreas de mayor aridez, y en las áreas de mayor humedad se encuentran los encinares con especies de hoja grande, existen algunas excepciones, como sucede con la distribución de Q. resinosa y Q. magnoliifolia. Al respecto, también debe considerarse el hábito foliar (caducifolio o perennifolio); así, la especie tardía de este estudio (Q. affinis) es perennifolia, mientras que Q. coccolobifolia es caducifolia. La pérdida total de las hojas parece ser una estrategia evolutiva de Q.coccolobifolia y otras colonizadoras para resistir la temporada seca del año (Reich et al., 1992). En efecto, las especies caducifolias suelen presentar altas tasas fotosintéticas y área foliar grande (Reich et al., 1992). En consecuencia, suelen crecer más rápido que las perennifolias (Antúnez et al., 2001), de ahí que éstas se distribuyan por lo general en áreas donde las condiciones ambientales les permiten la fotosíntesis durante todo el año (Chabot y Dicks, 1982). En ciertas regiones de México, el encinar perennifolio tiende a presentarse en lugares con climas templados y húmedos y suelos profundos, mientras que el encinar caducifolio suele encontrarse en áreas con clima cálido o templado y suelos someros (Guzmán, 1985).
El estudio de la repoblación de los claros adyacentes al bosque, indica que efectivamente Quercus coccolobifolia es la especie colonizadora en la comunidad estudiada, debido a que sus plántulas mostraron mayor tolerancia a luz directa, al establecerse principalmente en las áreas abiertas. Y, en el otro extremo de la secuencia sucesional, Q. affinis es la especie tardía, tolerante a sombra, debido a que sólo se estableció bajo el dosel del bosque. Q. crassifolia y Q. obtusata se ubican entre los extremos de este gradiente formado por Q. coccolobifolia y Q. affinis, ya que el número de sus plántulas y juveniles declinó gradualmente desde bajo el dosel y en el borde del mismo, hasta las áreas abiertas, por lo cual se pueden considerar como especies intermedias en esta secuencia. Esta tendencia de colonización preferentemente bajo el dosel del bosque se ha registrado en México también para Q. rugosa (Bonfil y Soberón, 1999), y para Q. crisipipilis y Q. laurina ( Quintana-Ascencio, 1989).
Otro aspecto que corrobora la secuencia sucesional postulada en este trabajo es la estimación de las edades de las especies que forman esta comunidad vegetal. Los individuos con mayor diámetro de Quercus coccolobifolia tienen una edad que varía de 76 a 347 años, mientras que en Q. crassifolia es de 142 a 222 años. La edad de los individuos de Q. affinis no se estimó con precisión, pero las características del ambiente donde crece y sus propios atributos, principalmente el hecho de ser perennifolio, hacen pensar que en general sea menor que la de las especies anteriores. Al respecto, (Löf, 2000) menciona que cuando las condiciones ambientales son favorables, los encinos presentan hasta tres crecimientos al año. Si se cumple este hecho para Q. affinis, se puede pensar que sus individuos alcanzan mayor tamaño a una menor edad.
Se debe considerar también la importancia de las perturbaciones, las cuales crean claros de diferente tamaño; en estos claros las condiciones de mayor luminosidad y temperatura, así como los menores niveles de humedad, permiten el establecimiento de especies de porte bajo, intolerantes a la sombra, bellota pequeña, capacidad de rebrote y gran superficie foliar, características que posee Q. coccolobifolia (Valdez y Aguilar, 1983; De la Cerda, 1989; Zavala, 1995).
Las teorías contemporáneas confieren a la competencia un papel central en la sucesión ecológica (Tilman, 1986; Huston y Smith, 1987; Cook, 1996), y se le ha reconocido como un proceso biótico importante en la estructuración de las comunidades vegetales (Connell, 1983; Shoener, 1983; Flores y Yeaton, 2000). El crecimiento asimétrico del dosel y del tronco, y el daño en las porciones apicales de las especies competitivamente inferiores, son efectos probables de la superioridad competitiva de una especie sobre otra. La ruptura de los ápices como resultado del crecimiento asimétrico del dosel y del tronco, está relacionada con las características físicas de la madera. Si bien, en relación con la densidad de la madera, Quercus affinis y Q. crassifolia son similares, en elasticidad son diferentes (Sotomayor, 2005). Por su parte, Q. coccolobifolia es la especie que presenta menor densidad y menor elasticidad del conjunto (Sotomayor, 2005). Aunque estas características varían entre regiones (De la Paz-Pérez, 2000), indudablemente son indicadores de la menor resistencia de la madera de Q. crassifolia y Q. coccolobifolia, en comparación con la de Q. affinis, lo cual también contribuye a la superioridad competitiva de esta especie.
En general, cualquier perturbación en un ecosistema trae como consecuencia inmediata la reducción de la dominancia de ciertas especies ya establecidas y la creación de nichos disponibles para la colonización y el crecimiento de nuevas especies, con lo cual se propicia un patrón en el remplazo de especies. La composición específica puede cambiar de manera continua como reacción a las variaciones más o menos aleatorias en el ambiente (Flores y Yeaton, 2000). Estos cambios se presentan por el efecto combinado de colonización y facilitación, seguidos de una presión competitiva que da lugar a un patrón oscilatorio en la dinámica de las poblaciones y que a su vez puede conducir a esquemas de sucesión cíclica (Horn, 1976; Yeaton, 1978; Vandermeer, 1980; Cody, 1993; Flores y Yeaton, 2000).
Aunque los factores físicos del medio podrían explicar la composición y estructura de estas comunidades vegetales (Noy Meir, 1979; Flores y Yeaton, 2000), en condiciones naturales, el remplazo por competencia de una especie por otra puede llegar a determinar la distribución de las especies en elespacio y en el tiempo. Así, de acuerdo con Callaway y Walker (1997), las comunidades vegetales deben en parte su organización a las relaciones bióticas entre las especies dominantes.
CONCLUSIONES
Los estudios realizados en el presente trabajo son experimentos naturales que permiten entender la dinámica de los bosques de encino, desde las especies que dominan los procesos en las diferentes fases de la sucesión, hasta la forma como las especies competitivamente superiores desplazan a otras.
Los resultados corroboran que la competencia interespecífica es importante en la estructuración de los encinares de la sierra de Álvarez. Las especies que dominan cada etapa de la dinámica de esta comunidad, forman una jerarquía competitiva donde Quercus coccoiobifoiia coloniza los claros del bosque. Esta especie cumple la función de nodriza de individuos jóvenes de Q. crassifolia y Q. obtusata, las cuales con el tiempo la remplazan. Éstas, a su vez, son remplazadas por Q. affinis, especie competitivamente superior o dominante con la madurez de la comunidad estudiada.
El estudio de la dinámica de los bosques de encino es complejo. Sin embargo, con las técnicas propuestas en este trabajo, es posible hacerlo debido a la coexistencia de los tres tipos de especies características de las fases de la dinámica de los bosques templados (apertura de claros-colonización, construcción y madurez). Además, con ellas es posible inferir el remplazo en el tiempo y evaluar la importancia de la facilitación y de la competencia en la formación de cada una de estas tres fases.
RECONOCIMIENTOS
Se agradece el apoyo del Fondo de Apoyo a la Investigación (FAI) de la UASLP (convenio C04-FAI-10-37.80) y al CO POCYT (convenio FMSLP-2002-5599) para financiar parte de esta investigación. El CONACYT otorgó sendas becas a Pedro Castillo Lara y Jorge Alberto Flores Cano para realizar sus estudios de doctorado en el PMPCA de la UASLP, de las cuales este trabajo constituye una parte.
REFERENCIAS
Antúnez, I., E.C. Retamosa y R. Villar. 2001. Relative growth rate in phylo-genetically related deciduous and evergreen woody species. Oecologia 128:172-180. [ Links ]
Arriaga, L., J.M. Espinoza, C. Aguilar, E. Martínez, L. Gómez y E. Loa (Coords.). 2000. Regiones terrestres prioritarias de México. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO). México, D.F. 465 p. [ Links ]
Barnett, R.J. 1977. The effect of burial by squirrels on germination and survival of oak and hickory nuts. American Midland Naturalist 98:319,329. [ Links ]
Bazzaz, F.A. 1998. Plants in changing environments: Linking physiological, population and community ecology. Cambridge University Press, Nueva York, EUA. 320 p. [ Links ]
Bello G., M.A. y J.N. Labat. 1987. Los encinos (Quercus) del estado de Michoacán, México. CEMCA-SARH (INIFAP). México. 97 p. [ Links ]
Beon, M.S. y N. Bartsch. 2003. Early seedling growth of pine (Pinus densi -flora) and oaks (Quercus serrata, Q.mongolica, Q. variabilis) in response to light intensity and soil moisture. Plant Ecology 167:97-105. [ Links ]
Bidwell, G.S. 1974. Plant physiology. Mac Millan Nueva York, EUA. 643 p. [ Links ]
Bonfil, C. y J. Soberón. 1999. Quercus rugosa seedling dynamics in relation to its re-introduction in a disturbed Mexican landscape. Applied Vegetation Science 2(2):189-200. [ Links ]
Boucher, D.H. 1981. Seed predation by mammals and forest dominance by Quercus oleoides, a tropical lowland oak. Oecologia 49:409-414. [ Links ]
Buckley, D.S., T.L. Sharik y J.G. Isebrands. 1998. Regeneration of northern red oak: positive and negative effects of competitors removal. Ecology 79(1):65-78. [ Links ]
Callaway, R.M.y L.R. Walker. 1997. Competition and facilitation: A synthetic approach to interactions in plant communities. Ecology 78:1958-1965. [ Links ]
Chabot, D.F. y D.J. Dicks. 1982. The ecology of leaf life spans. Ann. Rev. Ecol. Sys. 13:229-259. [ Links ]
Cody, M.L. 1993. Do cholla cacti (Opuntia spp., subgenus Cylindropuntia) use or need nurse plants in the Mojave Desert. J. Arid Envir. 24:139-154. [ Links ]
Connell, J.H. y R.O. Slatyer. 1977. Mechanisms of succession in natural communities and their role in community stability and organization. American Naturalist 111:1119-1144. [ Links ]
Connell, J.H. 1983. On the prevalence and relative importance of interspecific competition: evidence from field experiments. American Naturalist 122: 661-696. [ Links ]
Cook, J.E. 1996. Implications of modern successional theory for habitat typing: a review. Forest Science 42(1):62-75. [ Links ]
Darley-Hill, S. y W.C. Jhonson. 1981. Acorn dispersal by the blue jay (Cyanocitta cristata). Oecologia 50:231-232. [ Links ]
De la Cerda L., M. 1989. Encinos de Aguascalientes. Universidad Autónoma de Aguascalientes, Aguascalientes, México. 184 p. [ Links ]
De la Paz-Pérez O., C. 2000. Relación estructura propiedades físico-mecánicas de la madera de algunas especies de encinos (Quercus) mexicanas. Tesis Doctoral. UNAM. México, D. F. 266 p. [ Links ]
Flores M., G., J. Jiménez L., X. Madrigal S., F. Moncayo R. y F. Takaki T. 1971. Memoria del mapa de vegetación de la República Mexicana. Secretaría de Recursos Hidráulicos, México, D. F. 59 p. [ Links ]
Flores F., J.L. y R.I. Yeaton H. 2000. La importancia de la competencia en la organización de las comunidades vegetales en el Altiplano Mexicano. Interciencia 25(8):365-371. [ Links ]
Forcier, L.K. 1975. Reproductive strategies and the co-occurrence of climax tree species. Science 189:808-810. [ Links ]
García-Arévalo, A., J.J. Mendoza y J. Nocedal. 2004. Asociaciones vegetales de los bosques del municipio de Guanaceví, Durango. Madera y Bosques 10(1):21-34. [ Links ]
García S., F., J.R. Aguirre R., J. Villa-nueva D. y J. García P. 1999. Contribución al conocimiento florístico de la sierra de Álvarez, San Luis Potosí, México. Polibotánica 10: 73-103. [ Links ]
García S., F. y J.R. Aguirre R. Guía de campo para la identificación de las especies arbóreas de la sierra de Álvarez, S.L.P., México. (En preparación). [ Links ]
García, E. 1988. Modificaciones al sistema de clasificación climática de Köppen. Enriqueta García de Miranda (Ed.). México D. F. 217 p. [ Links ]
Guzmán M., R. 1985. Reserva de la Biosfera de la Sierra de Manantlán, Jalisco. Estudio descriptivo. Tiempos de Ciencia 1:10-26. [ Links ]
González-Espinosa, M., P. F. Quintana-Ascencio, N. Ramírez-Marcial y P. Gaytán. 1991. Secondary succession in disturbed Pinus-Quercus forests in the highlands of Chiapas, Mexico. Journal of Vegetation Science 2:351-360. [ Links ]
Huston, M.y T. Smith. 1987. Plant succession: life history and competition. American Naturalist 130:168-198. [ Links ]
Horn, H.S. 1974. The ecology of secondary succession. Ann. Rev. Ecol. Sys. 5:25-37. [ Links ]
Horn, H. S. 1976. Succession. In: R. M. May (Ed.). Theoretical ecology: principles and applications. Blackwell, Oxford, Reino Unido. pp. 187-204. [ Links ]
INEGI. 2002. Síntesis Geográfica del estado de San Luis Potosí. Aguascalientes, México. 200 p. [ Links ]
Jackson, J. 1984. Ecology of Missouri forests. Jefferson City: Missouri Department of Conservation, Jefferson City, EUA. 51 p. [ Links ]
Löf, M. 2000. Establishment and growth in seedlings of Fagus syivatica and Quercus robur: influence of interference from herbaceous vegetation. Canadian Journal Forestry Res. 30: 855-864. [ Links ]
Martínez R., M. 1985. Claros, ciclos vitales de los árboles tropicales y regeneración natural de las selvas altas perennifolias. In: A. Gómez-Pompa y S. del Amo (Eds.). Investigaciones sobre la regeneración de selvas altas en Veracruz, México. CECSA, México D. F. pp. 191-240. [ Links ]
Mellanby, K. 1968. The effects of some mammals and birds on regeneration of oak. The Journal of Applied Ecology 5(2):359-366. [ Links ]
Minckler, L.S. 1980. Woodland ecology: Environmental forestry for the small l and owner. Syracuse University Press, Syracuse, Nueva York, EUA. 160 p. [ Links ]
Nixon, K.C. 1984. A biosystematic study of Quercus series Virentes (the live oaks) with phylogenetic analices of Fagales, Fagaceae and Quercus. PhD. Dissertation. The University of Texas at Austin, Austin, Texas, EUA. 169 p. [ Links ]
Nixon, K.C. 1993. The genus Quercus in Mexico. In: T.P. Rammammorthy, R. Bye, A. Lot and J. Fa (Eds). Biological diversity of Mexico: Origins and distribution. Oxford University Press, Nueva York, EUA. pp 447-458. [ Links ]
Noy-Meir, I. 1979. Structure and function of desert ecosystem. Isr. J. Bot. 28: 1-19. [ Links ]
Perry, D.A. 1994. Forest ecosystems. The Johns Hopkins University Press, Baltimore, Maryland, EUA. 649 p. [ Links ]
Platt, J.R. 1964. Strong inference. Science 146:347-353. [ Links ]
Quintana-Ascencio, P.F. 1989. La condición sucesional de dos encinos dominantes (Quercus laurina Humb. & Bonpl. y Q. crispipilis Trel.) de los bosques de Los Altos de Chiapas. Tesis de Maestría en Ciencias. Colegio de Posgraduados, Chapingo, Edo. de México, México. 142 p. [ Links ]
Quintana-Ascencio, P.F. y M. González-Espinosa. 1993. Afinidad fitogeográfica y papel sucesional de la flora leñosa de los bosques de pino-encino de Los Altos de Chiapas, México. Acta Botánica Mexicana 21:43-57. [ Links ]
Reich, P.B., M.B. Walters y D.S. Ellsworth. 1992. Leaf life-span in relation to leaf plant, and stand characteristics among diverse ecosystem. Ecological Monographs 62:365-392. [ Links ]
Rzedowski, J. 1961. Vegetación del estado de San Luis Potosí. Tesis de Doctorado. Facultad de Ciencias, UNAM, México, D.F. 228 p. [ Links ]
Rzedowski, J. 1978. Vegetación de México. Limusa, México. 432 p. [ Links ]
Sánchez-Velázquez, L.R. y E. García-Moya. 1993. Sucesión forestal en los bosques mesófilo de montaña y de Pinus de la sierra de Manantlán, Jalisco, México. Agrociencia 3(1):7-26. [ Links ]
Shoener, T.W. 1983. Field experiments on interspecific competition. American Naturalist 122:240-285. [ Links ]
Shugart, H.H. 1984. The theory of forest dynamics: The ecological implications of forest succession models. Springer-Verlag, Nueva York, EUA. 278 p. [ Links ]
Siegel, S. y N.J. Castellan. 1988. Nonparametric statistics for behavioral sciences. 2nd ed. McGraw-Hill, Nueva York, EUA. 399 p. [ Links ]
Sork, V.L. 1984. Examination of seed dispersal and survival in red oak, Quercus rubra (Fagaceae), using metal-tagged acorns. Ecology 65:1020-1022. [ Links ]
Sotomayor C., J.R. 2005. Características mecánicas y clasificación de la madera de 150 especies mexicanas. Investigación e Ingeniería de la Madera 1(1):3-20. [ Links ]
Tilman, D. 1986. Evolution and differentiation in terrestrial plant communities: the importance of the soil resource: light gradient. In: J. Diamond and T. J. Case (Eds.). Community ecology. Harper & Row, Nueva York, N.J. EUA. pp. 359-380. [ Links ]
Valencia A., S. 2004. Diversidad del género Quercus (Fagaceae) en México. Bol. Soc. Bot. Méx. 75:33-53. [ Links ]
Valdez T., V. y M.L. Aguilar E. 1983. El género Quercus en las unidades fisonómico-florísticas del municipio de Santiago, N. L., México. Bol. Téc. Inst. Nac. Inv. For. México 98:275-288. [ Links ]
Vandermeer, J. 1980. Saguaros and nurse trees: a new hypothesis to account for population fluctuations. Southwestern Naturalist 25:357-360. [ Links ]
Watt, A.S. 1947. Pattern and process in the plant community. Journal of Ecology 35:1-22. [ Links ]
Whitmore, T.C. 1978. Gaps in the forest canopy. In: P.B. Tomlinson and M.H. Zimmermann (Editors). Tropical trees as living systems. Cambridge University Press, Londres, Reino Unido. pp. 639-655. [ Links ]
Yeaton, R.I. 1978. A cyclical relationship between Larrea tridentata and Opuntia leptocaulis in the northern Chihuahuan Desert. Journal of Ecology 66:651-656. [ Links ]
Yeaton, R.I. y A. Romero-Manzanares. 1986. Organization of vegetation mosaics in the Acacia schaffneri-Opuntia streptacantha association, southern Chihuahuan Desert, Mexico. Journal of Ecology 74: 211-217. [ Links ]
Yeaton, R.I., A. Romero-Manzanares, C. G. Vera y S. Vielegas. 1987. Tree succession in the subalpine forest of the neo-volcanic range, south-central Mexico. Southwestern Naturalist 32(3): 335-345. [ Links ]
Zavala Ch., F. 1995. Encinos y robles, notas fitogeográficas. Universidad Autónoma Chapingo, Chapingo, Texcoco, Edo. de México, México. 44 p. [ Links ]
Zavala Ch., F. y E. García Moya. 1996. Frutos y semillas de encino. Universidad Autónoma Chapingo, Chapingo, Texcoco, Edo. de México, México. 47 p. [ Links ]
Zavala Ch., F. 2003. Identificación de encinos de México. Universidad Autónoma Chapingo, Chapingo, Texcoco, Edo. de México, México. 188 p. [ Links ]
Zavala Ch., F. 2004. Desecación de bellotas y su relación con la viabilidad en nueve especies de encinos mexicanos. Ciencia Ergo Sum 11(2): 177-185. [ Links ]
Nota
Este documento debe citarse como: Castillo Lara, P., J.A. Flores Cano, J.R. Aguirre Rivera y R.I. Yeaton H. 2008. Dinámica sucesional del encinar de la sierra de Álvarez, San Luis Potosí, México. Madera y Bosques 14(1):21-35.

