











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
Introduction
In 2019, tomatillo was grown in 30 states of the Mexican Republic (Sistema de Información Agroalimentaria y Pesquera [SIAP], 2021), where it is cultivated with everything from the traditional rainfed system with direct sowing to fertigation with mulching and trellising. In the spring-summer and autumn-winter cycles, it is grown in both irrigated and rainfed systems, so the product is available in the market all year round (Peña-Lomelí, Santiaguillo-Hernández, & Magaña-Lira, 2007).
Most farmers use seed that they get from their crops. In some parts of the country, farmers use commercial varieties obtained from the multiplication of outstanding landrace materials, but which have not undergone a robust breeding program (Magaña-Lira, Santiaguillo-Hernández, & Grimaldo-Juárez, 2011), and aspects such as identity, genetic purity, and quality (physical, physiological, sanitary and genetic), among other agronomic attributes, cannot be guaranteed. Therefore, it is important to produce certified seed of improved and registered varieties in order to give certainty to the producer of the quality of the seed and improve crop productivity (Santiaguillo-Hernández et al., 2012).
Tomatillo has gametophytic self-incompatibility determined by two independent genes with multiple alleles (Mulato-Brito, Peña-Lomelí, Sahagún-Castellanos, Villanueva-Verduzco, & López-Reynoso, 2007; Pandey, 1957), which is why it is not possible to form highly inbred lines or hybrids with classical techniques. Consequently, its genetic improvement is based on selection (Peña-Lomelí & Márquez-Sánchez, 1990).
Inbreeding is the mating of individuals that are related to each other by ancestry. The essential consequence of two individuals having a common ancestor is that both of them may carry replicates of one of the genes present in the ancestor and, if they mate, they can inherit it. The most notable effect of inbreeding is loss of vigor or inbreeding depression, characterized by a reduction in the phenotypic value shown by traits related to reproductive ability or physiological efficiency (Falconer & Mackay, 1996).
Heterosis occurs in crosses of inbred lines and in intervarietal crosses, as in the case of tomatillo (Sahagún-Castellanos, Gómez-Ruiz, & Peña-Lomelí, 1999). Peña-Lomelí et al. (1998) worked with eight varieties analyzed in a dialelic and found that the mean heterosis was significant for total yield (17.19 %) and number of fruits per plant (10.42 %), so it would be possible to obtain hybrids with better traits than the varieties. Santiaguillo-Hernández, Cervantes-Santana, and Peña-Lomelí (2004) found significant positive average heterosis for total yield per plant (21.4 %) in plant-to-plant crosses between Verde Puebla and CHF1 Chapingo varieties, although in fruit quality traits the average heterosis was negative.
Peña-Lomelí et al. (1999) point out that it is possible to obtain hybrids from crosses between maternal half-sib families (MHSF), and Santiaguillo-Hernández et al. (2004) note that they can also be obtained by plant-to-plant crosses. However, maintaining the stability of families through seed-increasing cycles can be a problem due to the small number of individuals, which can generate inbreeding depression. One way to conserve the genotype of families or progenitor plants of hybrids is in vitro multiplication (Andrade-Rodríguez, López-Peralta, González-Hernández, García-Velázquez, & Peña-Lomelí, 2005), although the complete protocol for mass propagation is not yet available.
Therefore, the objective of this work was to study the effect of inbreeding in families of four varieties of tomatillo obtained through three generations of mating by fraternal and plant-to-plant crosses.
Materials and methods
The tomatillo varieties evaluated were Tecozautla, Diamante, Manzano and Morado San Miguel (Servicio Nacional de Inspección y Certificación de Semillas [SNICS], 2021). In each one, three MHSFs obtained from six seed-increasing cycles by fraternal crosses (Generation 1) were studied.
In the autumn-winter 2018-2019 cycle, 114 plants of each family (12 in total) were established in a hydroponic greenhouse system. The five best plants of each family were selected using the criteria of height, vigor and number of flowers at 15 days after transplantation (dat). Plant-to-plant and fraternal crosses were made in the selected plants using the controlled pollination technique described by Peña-Lomelí, Magaña-Lira, Gámez-Torres, Mendoza-Celino, and Pérez-Grajales (2018).
The plant-to-plant crosses were made in a chain. The first plant was crossed as a male with the second, this with the third and so on, and the fifth plant was crossed as a male with the first. For the fraternal crosses, pollen from the five plants was placed in a Petri dish and mixed, and then each plant was pollinated. In plant-to-plant and fraternal crosses, each selected plant gave rise to a full-sib family (FSF) and a MHSF, respectively (Generation 2).
In the fall-winter 2019-2020 cycle, families resulting from Generation 2 were established in a hydroponic greenhouse system. As some crosses did not produce enough seed, plants from families having both FSFs and MHSFs were chosen. Therefore, five plants within each family were selected. In the previous cycle, both types of crosses were made for each plant, and in the new selection cycle only one type of cross was made per plant, which depended on its origin. If the plant came from a FSF, plant-to-plant crosses were made; if it came from a MHSF, fraternal crosses were made. This gave rise to new FSFs and MHSFs (Generation 3). For field evaluation, all the families obtained were used.
In the spring-summer cycle of 2020, the field evaluation was made of those families obtained in both selection-cross cycles in the greenhouse and that had enough seed, plus the original families (that came from the six seed-increasing cycles by fraternal crosses); this gave rise to 82 families (Generations 1, 2 and 3). Sowing was done on February 29, and 200-cavity polystyrene trays with peat moss as substrate were used. One to two seeds were placed per cavity depending on their availability. Seedlings were kept under greenhouse conditions and watered every other day with 50 % Steiner’s nutrient solution for three weeks; thereafter, they were irrigated daily with 100 % Steiner's nutrient solution (Steiner, 1984). Fifteen days after sowing, one plant per cavity was thinned.
The evaluation was established in lot 10-C (19° 27’ 60’’ N and 98° 54’ 19’’ W) of the experimental field of the Colegio de Postgraduados, Montecillo campus, State of Mexico. The land was prepared with fallowing and harrowing; the furrowing was done at 1 m apart. Transplanting was carried out on April 16 with 30 cm spacing between plants. A randomized complete block design with four replications was established. Each experimental unit consisted of a 4.2 m furrow (15 plants). Irrigation was carried out weekly using siphons and fertilization was done with 200 N - 100 P2O5 - 50 K2O. The first fertilization was made one week after transplantation: half the urea and all the diammonium phosphate and potassium chloride were applied. The second fertilization was performed 28 dat and the rest of the urea was applied. In addition to weekly applications of foliar fertilizer (Bayfolan Forte® + surfactant), preventive sprayings for pests and diseases (Confidor® [Imidacloprid] and Lannate® [Methomyl]) were carried out. Weeds were controlled with three hoeing and two weeding operations, and 112.5 ga.i.·ha-1 of Provence 75 WG® (Isoxaflutole) were applied after earthing up.
At 40 dat, data were taken on plant height and number of bags formed (set fruits). Height (PH, cm) was taken from ground level to the tip of the highest branch. For the number of bags (NB), calyces with fruit development were counted. For both variables, the mean per experimental unit was obtained.
Harvesting was carried out in two harvests: June 30 and July 10. In both, yield per plant (YP1 and YP2, kg) and total yield per plant (TYP, kg) were evaluated; in addition, 10 fruits were weighed at random (W10F1 and W10F2, g) and only in the first harvest was the volume (V1, mL) measured. From the weight and volume of 10 fruits, the bulk density of fruits in the first harvest (D1, g·mL-1) was estimated, which in turn was used to estimate the volume in the second harvest (V2, mL). The average weight and volume of the 10 fruits (AW10F and AV, respectively) were also obtained. The weight of 10 fruits was considered as an estimator of their size (very large, large and medium fruit) (Peña-Lomelí, Ponce-Valerio, Sánchez-del Castillo, & Magaña-Lira, 2014).
The analysis of variance was performed according to the following model: Y ijkl = μ + β l + V i + G (i)j + C (i)k + GC (i)jk + ε ijkl , where Y ijkl is the observed value of the variable Y, µ is the overall mean, β l is the effect of block l, V i is the effect of variety i, G (i)j is the effect of generation j nested in variety i, C (i)k is the effect of the type of cross k nested in variety i, GC (i)jk is the generation x cross interaction nested in variety i and ε ijkl is the random error. Mean comparison tests (Tukey, P ≤ 0.05) were performed between varieties, between generations for each variety and between crosses for each variety.
Results and discussion
The analysis of variance (Table 1) shows that Variety had a highly significant effect (P ≤ 0.01) on all variables. This indicates different behaviors among varieties, which is related to the distinctive traits of each one of them, such as fruit size and yield per plant. The effect of the Generation nested in Variety factor was not significant in YP2 and D1; in PH and V2 the effect was significant (P ≤ 0.05), and in the rest of the variables it was highly significant (P ≤ 0.01). This may indicate inbreeding depression across generations in at least one of the varieties studied. The Cross nested in Variety factor had no significant effect on YP2 and D1, but it was significant (P ≤ 0.05) for PH, W10F2, V1 and V2; the effects were highly significant (P ≤ 0.01) for the rest of the variables. This implies that, at least for one variety, the type of cross produces a different effect. The Generation x Cross interaction was not significant.
Table 1 Mean squares of the analysis of variance for twelve variables evaluated in four varieties of tomatillo (Physalis ixocarpa Brot. ex Horm.).
| Source | DF | PH | NB | YP1 | YP2 | TYP | D1 |
|---|---|---|---|---|---|---|---|
| Block | 3 | 258.50** | 151.76** | 0.433** | 0.060* | 0.496** | 0.0015 |
| V | 3 | 501.37** | 326.12** | 0.903** | 0.078** | 1.165** | 0.1182** |
| G(V) | 7 | 55.93* | 21.51** | 0.589** | 0.024 | 0.748** | 0.0177 |
| C(V) | 4 | 74.40* | 27.72** | 0.317** | 0.021 | 0.390** | 0.0032 |
| GxC(V) | 2 | 69.15 | 2.11 | 0.047 | 0.020 | 0.066 | 0.0029 |
| Error | 308 | 26.40 | 7.07 | 0.074 | 0.017 | 0.094 | 0.0102 |
| Total | 327 | ||||||
| CV | 19.68 | 35.20 | 40.68 | 76.75 | 36.45 | 11.03 | |
| W10F1 | W10F2 | AW10F | V1 | V2 | AV | ||
| Block | 3 | 11,705.0 | 834.6 | 2,888.8 | 19,074.9 | 1,361.5 | 5,558.5 |
| V | 3 | 604,945.7** | 110,752.1** | 306,452.1** | 974,867.1** | 189,781.4** | 503,826.6** |
| G(V) | 7 | 102,098.8** | 19,256.3** | 51,244.2** | 95,553.7** | 17,199.2* | 46,768.8** |
| C(V) | 4 | 39,958.3** | 13,733.2* | 21,726.6** | 50,067.6* | 18,928.5* | 28,955.8** |
| GxC(V) | 2 | 7,895.7 | 1,795.1 | 2,008.0 | 10,884.1 | 2,902.3 | 3,433.9 |
| Error | 308 | 11,453.5 | 5,103.1 | 5,929.9 | 15,044.8 | 6,807.4 | 7,876.9 |
| Total | 327 | ||||||
| CV | 26.41 | 35.90 | 25.49 | 27.31 | 37.34 | 26.49 |
Source = source of variation; V = variety; G(V) = generation nested in variety; C(V) = type of cross nested in variety; GxC(V) = generation x type of cross interaction, nested in variety; CV = coefficient of variation; DF = degrees of freedom; PH = plant height (cm); NB = number of bags; YP1 and YP2 = yield per plant in harvest 1 and 2, respectively (kg); TYP = total yield per plant (kg); D1 = density of 10 fruits in harvest 1 (g·mL-1); W10F1 and W10F2 = weight of 10 fruits in harvest 1 and 2, respectively (g); AW10F = average weight of 10 fruits (g); V1 and V2 = volume of 10 fruits in harvest 1 and 2, respectively (mL); AV = average volume of 10 fruits (mL); ** y * = significant with P ≤ 0.01 and P ≤ 0.05, respectively.
The Manzano variety had the greatest PH, while the others did not differ statistically from each other (P ≥ 0.05). Diamante had the highest NB, followed by Morado, Tecozautla and Manzano. In YP1 and TYP, Diamante was superior, although in the second harvest it was not different from Morado. D1 was no different in Diamante, Manzano and Morado, but all three were superior to Tecozautla. In W10F1 and AW10F. Tecozautla was superior, but in W10F2 it was not different from Diamante. Regarding the volume variables, Tecozautla was superior in all three (Table 2).
Table 2 Comparison of means among four varieties of tomatillo (Physalis ixocarpa Brot. ex Horm.).
| Variety | PH | NB | YP1 | YP2 | TYP | D1 |
|---|---|---|---|---|---|---|
| Tecozautla | 25.63 bz | 6.65 c | 0.610 b | 0.137 b | 0.747 bc | 0.832 b |
| Diamante | 24.38 b | 11.11 a | 0.841 a | 0.227 a | 1.068 a | 0.964 a |
| Manzano | 30.46 a | 5.08 d | 0.598 b | 0.125 b | 0.723 c | 0.935 a |
| Morado | 23.77 b | 7.92 b | 0.658 b | 0.210 a | 0.868 b | 0.949 a |
| HSD | 2.08 | 1.08 | 0.110 | 0.054 | 0.124 | 0.041 |
| W10F1 | W10F2 | AW10F | V1 | V2 | AV | |
| Tecozautla | 490.0 a | 238.2 a | 364.1 a | 590.6 a | 287.9 a | 439.3 a |
| Diamante | 402.1 b | 210.2 ab | 306.1 b | 420.6 b | 221.0 b | 320.7 b |
| Manzano | 430.3 b | 193.2 b | 311.7 b | 462.7 b | 208.1 b | 335.5 b |
| Morado | 294.0 c | 154.2 c | 224.1 c | 311.6 c | 163.6 c | 237.6 c |
| HSD | 43.3 | 28.9 | 31.2 | 49.6 | 33.4 | 35.9 |
PH = plant height (cm); NB: number of bags; YP1 and YP2 = yield per plant in harvest 1 and 2, respectively (kg); TYP = total yield per plant (kg); D1 = density of 10 fruits in harvest 1 (g·mL-1); W10F1 and W10F2 = weight of 10 fruits in harvest 1 and 2, respectively (g); AW10F = average weight of 10 fruits (g); V1 and V2 = volume of 10 fruits in harvest 1 and 2, respectively (mL); AV = average volume of 10 fruits (mL); HSD = honestly significant difference. zMeans with the same letter within each column do not differ statistically (Tukey, P ≤ 0.05).
Tecozautla was the variety with the largest fruit, but with the lowest density. Diamante had the highest earliness and yield. The growth habit in Manzano was more erect than in the others. Morado was the variety with the smallest fruits. The foregoing coincides with what was reported by Sánchez-Martínez and Peña-Lomelí (2015) regarding the distinguishing traits of these varieties.
PH was only different among generations for the Manzano variety, where Generation 2 had the highest value (Figure 1a). Generation 1 of the Diamante variety had fewer bags than Generations 2 and 3, which were not different from each other. In the rest of the varieties, no differences in NB were found (Figure 1b). YPC1 and TYP were not different between Generations 1 and 2 in all varieties, but in Tecozautla and Diamante a significantly lower yield was obtained in Generation 3 (Figures 1c and 1e). According to Márquez-Sánchez (2007), decreased yield is a measurable indicator of inbreeding depression. YP2 and D1 were not different among generations for any variety (Figures 1d and 1f).
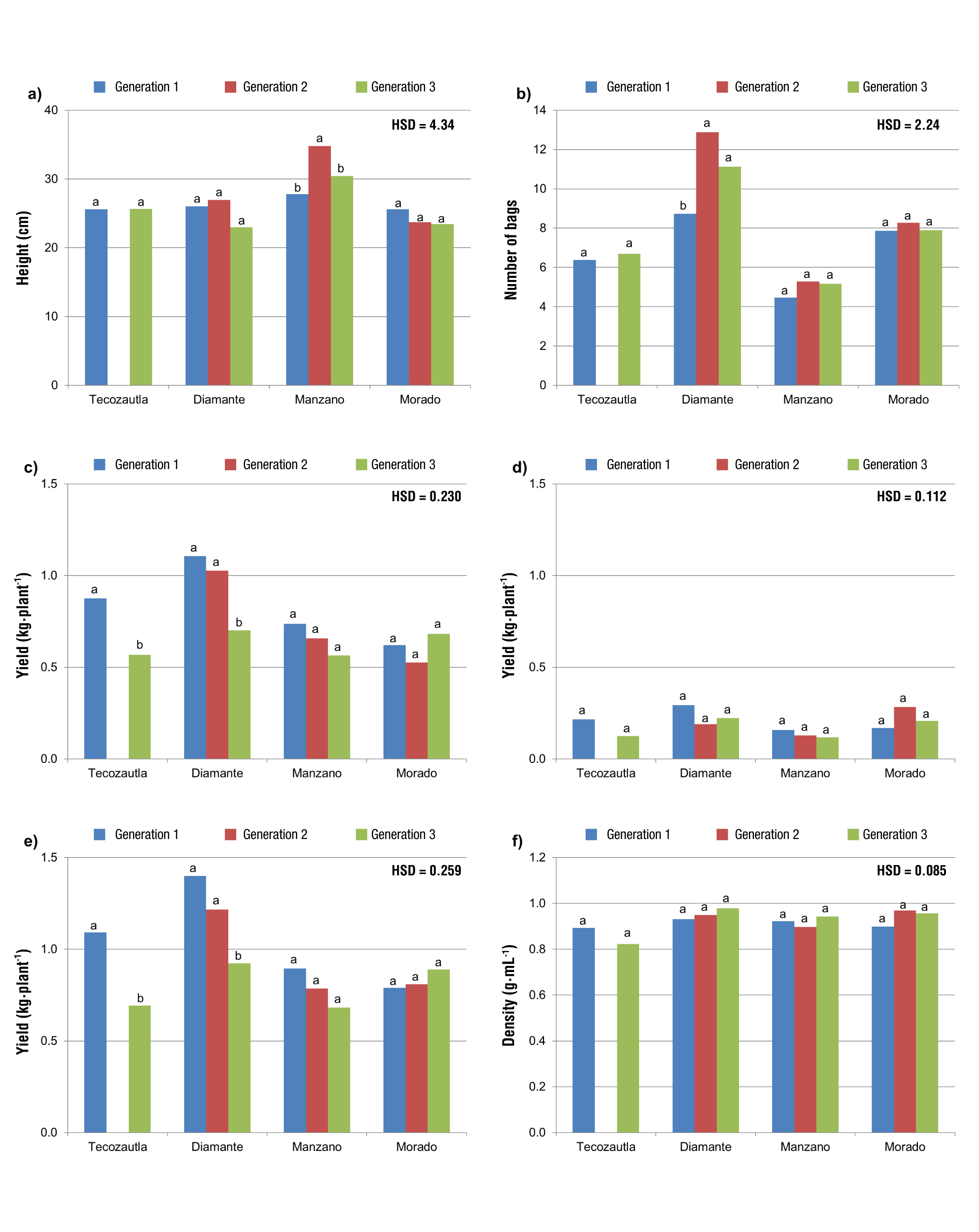
Figure 1 Comparison of means in three generations of crosses for four varieties of tomatillo (Physalis ixocarpa Brot. ex Horm.): a) plant height (PH, cm), b) number of bags (NB), c) yield per plant in harvest 1 (YP1, kg), d) yield per plant in harvest 2 (YP2, kg), e) total yield per plant (TYP, kg) and f) density of 10 fruits in harvest 1 (D1, g·mL-1). HSD = honestly significant difference. Means with the same letter in each variety do not differ statistically (Tukey, P ≤ 0.05).
In Tecozautla and Diamante, Generation 1 was superior in W10F1, W10F2, AW10F, V1, V2 and AV. Similarly, Generations 2 and 3 were not different from each other in Diamante (Figure 2). It should be remembered that there was not enough seed from Tecozautla’s Generation 2 for field evaluation. In Manzano, a decrease was only observed in W10F1 and V1 (Figures 2a and 2d), where Generation 1 was significantly superior to Generations 2 and 3, between which there were no significant differences. For the rest of the variables, no differences were found among generations. The Morado variety did not show significant differences among generations for any variable (Figures 1 and 2).

Figure 2 Comparison of means in three generations of crosses for four varieties of tomatillo (Physalis ixocarpa Brot. ex Horm.): a) weight of 10 fruits in harvest 1 (W10F1, g), b) weight of 10 fruits in harvest 2 (W10F2, g), c) average weight of 10 fruits (AW10F, g), d) volume of 10 fruits in harvest 1 (V1, mL), e) volume of 10 fruits in harvest 2 (V2, mL) and f) average volume of 10 fruits (AV, mL). HSD = honestly significant difference. Means with the same letter in each variety do not differ statistically (Tukey, P ≤ 0.05).
In summary, the comparison of means indicates that fruit yield, size and volume decreased in Tecozautla and Diamante across generations. Yield decreased after the second generation, while size and volume did so after the first generation. In Manzano only fruit size and volume in the first harvest were affected, in both cases after Generation 1. This is an indicator of inbreeding depression, as pointed out by Márquez-Sánchez (2007). Based on the above, it can be said that inbreeding depression had a greater effect on the Tecozautla and Diamante varieties, which may be due to their initial level of inbreeding. Both have more selection cycles than Manzano and Morado (Peña-Lomelí et al., 2018; Sánchez-Martínez & Peña-Lomelí, 2015), so it is inferred that there is more inbreeding in Tecozautla and Diamante.
PH was only different between cross types in Manzano, where families obtained by plant-to-plant crosses were superior (Figure 3a). NB and YPC1 were different in Morado, with families obtained from fraternal crosses being superior (Figures 3b and 3c). The type of cross had an effect on yield variables in Diamante, since, in both harvests and total yield, families from plant-to-plant crosses had lower yields (Figures 3c, 3d and 3e). In the Tecozautla variety, no effect of the type of cross was found on PH, NB or yield variables (Figure 3). Density was not affected by the type of cross.

Figure 3 Comparison of means in two types of crosses for four varieties of tomatillo (Physalis ixocarpa Brot. ex Horm.): a) plant height (PH, cm); b) number of bags (NB), c) yield per plant in harvest 1 (YP1, kg), d) yield per plant in harvest 2 (YP2, kg), e) total yield per plant (TYP, kg) and f) density of 10 fruits in harvest 1 (D1, g·mL-1). HSD = honestly significant difference. Means with the same letter in each variety do not differ statistically (Tukey, P ≤ 0.05).
Due to the type of family generated by each cross, it was expected that the variables evaluated would have lower expression in the families derived from plant-to-plant crosses, since mating between full siblings generates greater inbreeding than between half-siblings (Falconer & Mackay, 1996). This may be because the crosses were made between selected plants in both cases and possibly still have high heterozygosity. Hallauer, Carena, and Miranda (2010) note that the change in vigor is directly proportional to the change in heterozygosity in the population, i.e., the higher the heterozygosity, the higher the yield.
Fruit size (weight and volume) in the first harvest was different between the type of crosses in Tecozautla and Morado. In both cases, a significant decrease was observed in plant-to-plant crosses with respect to fraternal crosses (Figures 4a and 4d). In Tecozautla and Diamante, differences were found between the type of crosses for W10F2 and V2 (Figures 4b and 4e). The families obtained by plant-to-plant crosses had lower values than those of the fraternal crosses. In the average fruit size of Tecozautla, Diamante and Morado, it was also found that the families of plant-to-plant crosses were inferior to those of fraternal crosses (Figures 4c and 4f). In general, no differences were found between the type of crosses for Manzano (Figures 3 and 4).
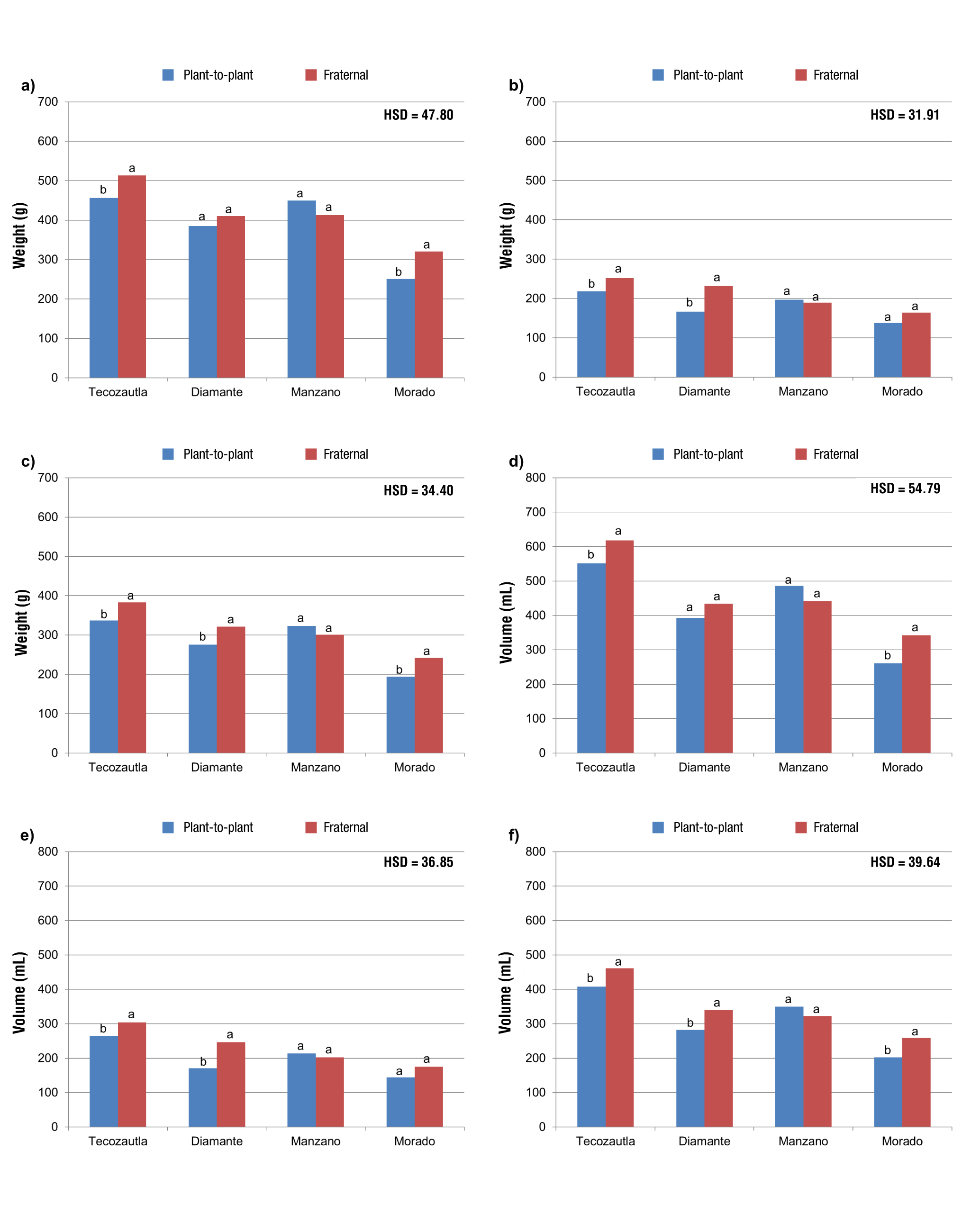
Figure 4 Comparison of means of two types of crosses for four varieties of tomatillo (Physalis ixocarpa Brot. ex Horm.): a) weight of 10 fruits in harvest 1 (W10F1, g), b) weight of 10 fruits in harvest 2 (W10F2, g), c) average weight of 10 fruits (AW10F, g), d) volume of 10 fruits in harvest 1 (V1, mL), e) volume of 10 fruits in harvest 2 (V2, mL) and f) average volume of 10 fruits (AV, mL). HSD = honestly significant difference. Means with the same letter in each variety do not differ statistically (Tukey, P ≤ 0.05).
Tecozautla performed differently in the variables related to fruit size and volume. Diamante was different in TYP, AW10F and AV. Manzano was different only in PH. Morado had different behavior in YP1, W10F1, V1 and AV. In Tecozautla, Diamante and Morado, the fraternal cross was superior, whereas in Manzano the plant-to-plant cross was better. In general, this indicates that plant-to-plant crosses generated a greater inbreeding depression than fraternal crosses, except in Manzano.
The greater inbreeding depression in the plant-to-plant cross is due to the fact that only two individuals come into play during pollination, while five are involved in the fraternal cross. With fewer individuals, the probability of close relatives mating increases and thus homozygosity as well, so the frequency of dominant phenotypes decreases, and the deleterious effects of recessive alleles increase. Inbreeding is greater in FSFs than in MHSFs (Falconer & Mackay, 1996; Hallauer et al., 2010).
Results show that it is possible to develop a genetic improvement program by hybridization in tomatillo, despite the self-incompatibility, since it has been demonstrated that plant-to-plant crosses do indeed generate inbreeding. However, it is necessary to consider that to generate a level of inbreeding similar to that of a selfing cycle, several cycles of plant-to-plant crosses are required (Falconer & Mackay, 1996); therefore, the time needed to obtain highly inbred FSFs would be much longer.
Conclusions
Inbreeding depression occurred between generations in fruit yield, size and volume, which decreased across generations.
The Tecozautla and Diamante varieties showed a greater inbreeding depression than Manzano and Morado, as they had a decreasing trend in the variables evaluated across generations.
The type of cross had a different effect depending on the variety. In the variables related to fruit size, the plants originated by plant-to-plant crosses had lower values in Tecozautla, Diamante and Morado, which indicates that a greater inbreeding depression is generated with this type of cross.
It is possible to obtain highly inbred full-sib families for a tomatillo hybridization breeding program.

