










Serviços Personalizados
Journal
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista Chapingo. Serie horticultura
versão On-line ISSN 2007-4034versão impressa ISSN 1027-152X
Rev. Chapingo Ser.Hortic vol.24 no.1 Chapingo Jan./Abr. 2018
https://doi.org/10.5154/r.rchsh.2016.11.029
Artículo científico
Crecimiento y contenido de azúcares de tubérculo de papa en cuatro estados de madurez en condiciones de invernadero
1Benemérita Universidad Autónoma de Puebla, Facultad de Ingeniería Agrohidráulica. Av. Universidad s/n, San Juan Acateno, Teziutlán, Puebla, C. P. 73695, MÉXICO.
2Universidad Autónoma Chapingo, Departamento de Fitotecnia. Carretera México-Texcoco km 38.5, Chapingo, Estado de México, C. P. 56230, MÉXICO.
3Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Centro - Altos de Jalisco. Carretera Tepatitlán-Lagos de Moreno km 8, colonia Rancho las Cruces, Tepatitlán de Morelos, Jalisco, C. P. 47600, MÉXICO.
El contenido de azúcares en el tubérculo de papa determina el momento óptimo de cosecha, el tipo de procesamiento y su manejo durante el almacenamiento. La presente investigación tuvo como objetivo evaluar el crecimiento y rendimiento de cuatro variedades de papa en condiciones de invernadero y determinar el contenido de azúcares solubles de este tubérculo en diferente estado de madurez. Para ello, se muestrearon papas (5 ± 1 g) de cuatro variedades en cuatro estados de madurez, desde el inicio de la tuberización hasta la madurez fisiológica, en los que se determinó el contenido de azúcares solubles por cromatografía líquida de alta resolución (HPLC). La variedad Alpha tuvo el mayor ciclo biológico (1,230 grados día, P ≤ 0.05); en tanto que Mondial presentó el máximo rendimiento (380 g, P ≤ 0.05), 75, 31 y 31 % más que Vivaldi, Alpha y Atlantic, respectivamente, atribuido al mayor peso promedio de tubérculo (61 g, P ≤ 0.05). El contenido de sacarosa, glucosa y fructosa más alto (7.65, 2.22 y 0.37 mg·g-1, respectivamente) se presentó 21 días después del inicio de tuberización; cantidades que disminuyeron (P ≤ 0.05) durante el desarrollo de tubérculos. Atlantic destaca con los niveles más bajos (P ≤ 0.05) de sacarosa, glucosa, fructosa y azúcares totales en los diferentes estados de madurez del tubérculo. El peso promedio del tubérculo se correlacionó en forma negativa (P ≤ 0.05) con los contenidos de sacarosa, azúcares reductores (glucosa y fructosa) y totales.
Palabras clave: Solanum tuberosum; fenología; peso de tubérculos; sacarosa; azúcares reductores
The sugar content in the potato tuber determines the optimum harvest time, the type of processing and its handling during storage. The aim of this study was to evaluate the growth and yield of four potato varieties under greenhouse conditions and to determine the soluble sugar content of this tuber at different maturity stages. For this, potatoes (5 ± 1 g) of four varieties were sampled at four maturity stages, from the beginning of tuberization to physiological maturity, in which the soluble sugar content was determined by high-performance liquid chromatography (HPLC). The Alpha variety had the longest growth cycle (1,230 degree days, P ≤ 0.05), while Mondial had the highest yield (380 g, P ≤ 0.05), 75, 31 and 31 % more than Vivaldi, Alpha and Atlantic, respectively, attributed to the highest average tuber weight (61 g, P ≤ 0.05). The highest content of sucrose, glucose and fructose (7.65, 2.22 and 0.37 mg·g-1, respectively) was recorded 21 days after the start of tuberization, quantities that decreased (P ≤ 0.05) during tuber development. Atlantic stands out with the lowest levels (P ≤ 0.05) of sucrose, glucose, fructose and total sugars in the different stages of tuber maturity. Average tuber weight was negatively correlated (P ≤ 0.05) with the contents of sucrose, reducing sugars (glucose and fructose) and total sugars.
Keywords: Solanum tuberosum; phenology; tuber weight; sucrose; reducing sugars
Introducción
La papa (Solanum tuberosum L.) es el cuarto cultivo alimenticio de mayor importancia a nivel mundial, superado por el arroz (Oryza sativa L.), trigo (Triticum aestivum L.) y maíz (Zea mays L.), y juega un papel crucial en la seguridad alimentaria y el desarrollo económico de la humanidad (Zhang, Hou, Liu, Xie, & Song, 2014). Los tubérculos son ricos en carbohidratos, proteínas, minerales y vitaminas, además de poseer ciertas propiedades antioxidantes (Suttle, 2008).
La producción de materia seca en el cultivo de papa es determinada por factores de diferente índole; uno de los que más influye es la duración del ciclo biológico, expresado a través de un conjunto de fases y etapas fenológicas, y que a su vez dependen del ambiente y del cultivar (Kooman, Fahem, Tegera, & Haverkort, 1996). En condiciones no limitantes, la temperatura puede afectar en forma drástica la producción de biomasa, especialmente cuando supera los 20 °C durante la etapa de llenado del tubérculo (Rykaczewska, 2015); por lo que la regulación de este factor es preponderante, especialmente en la agricultura protegida.
El rendimiento, definido como el producto del número de tubérculos por planta y su peso (Lynch, Kozub, & Kawchuk, 2001), está determinado por el genotipo, el ambiente y su interacción (Milton & Allen, 1995). Así, la formación de tubérculos en la planta de papa es un proceso complejo que involucra diferentes sistemas biológicos inducidos mediante señales ambientales, como el fotoperíodo corto, temperaturas bajas y contenido alto de humedad en el suelo (Cenzano, Abdala, & Hause, 2007).
La inducción de la tuberización se produce en las hojas a través del fitocromo. Inicialmente, el ácido giberélico activa una señal transmisible a la región subapical del estolón; esto causa que la división celular genere, en lugar de crecimiento longitudinal, una expansión radial (Cenzano et al., 2007; Hannapel, Chen, Rosin, Banerjee, & Davies, 2004; Viola et al., 2007) mediante el transporte apoplástico hacia el meristemo apical y la región subapical (Hancock, Roberts, & Viola, 2008). De esta manera, el desarrollo del tubérculo ocurre gracias a la descarga de fotoasimilados en células del parénquima lateral que produce ensanchamiento de la región subapical por la acumulación de proteínas y carbohidratos, que más tarde son transformados en almidones (Cenzano et al., 2007; Hancock et al., 2008).
Durante la etapa de tuberización ocurre la acumulación masiva de almidón en el tubérculo, lo que representa una gran demanda de carbohidratos (Sabba et al., 2007); esto provoca que los niveles más altos de sacarosa, glucosa y fructosa se registran en tubérculos jóvenes o inmaduros (Stark & Love, 2003), y a medida que alcanzan la madurez fisiológica dichas concentraciones tienden a disminuir (Knowles, Driskill, & Knowles, 2009). En algunas investigaciones se reportan valores de 0.2 a 1.5 % de sacarosa y de 0.01 a 0.7 % de azúcares reductores en tubérculos inmaduros (Kolbe & Stephan-Beckmann, 1997; Kumar & Ezekiel, 2006); mientras que durante la madurez fisiológica, se han encontrado de 0.1 a 0.6 % de sacarosa y de 0.04 a 0.4 % de azúcares reductores (Knowles et al., 2009; Kumar & Ezekiel, 2006; Sabba et al., 2007).
La cuantificación exacta de azúcares en papa es importante para propósitos científicos y comerciales. Los principales azúcares presentes en este tubérculo son la sacarosa, glucosa, fructosa (Morales-Fernández et al., 2015) y pequeñas cantidades de maltosa durante la brotación (Ilin, Durovka, & Markovic, 1997), cuyos contenidos son afectados por el genotipo, el estado fenológico, el manejo del cultivo, factores ambientales y condiciones de almacenamiento (Thompson, Love, Sowokinos, Thornton, & Shock, 2008).
Con base en lo anterior, la presente investigación tuvo como propósito evaluar el crecimiento y rendimiento de cuatro variedades de papa en condiciones de invernadero y determinar el contenido de azúcares solubles de este tubérculo en diferente estado de madurez.
Materiales y métodos
La investigación se realizó en el Instituto de Horticultura de la Universidad Autónoma Chapingo (19° 29’ latitud norte y 98° 53’ longitud oeste, a 2,250 msnm), entre los meses de octubre (2008) y febrero (2009), bajo condiciones de invernadero.
Las variedades comerciales de papa estudiadas fueron Alpha, Atlantic, Mondial y Vivaldi. En la siembra se usaron minitubérculos de 23 a 32 mm de diámetro en estado fisiológico de dominancia apical. Se establecieron dos minitubérculos de cada variedad en bolsas de polietileno color negro con capacidad de 6 L, con sustrato compuesto por la mezcla de turba - perlita (1:1 v/v), a 10 cm de profundidad. Las bolsas se colocaron con separación de 30 y 10 cm entre hileras y filas, respectivamente.
En la siembra, al sustrato de cada bolsa se le aplicó 1.2 g de la mezcla de N, P, K, Ca y Mg, con 34, 31, 27, 4 y 4 %, respectivamente. Durante el ciclo de cultivo, el riego se aplicó de forma manual a capacidad de campo, de acuerdo con las necesidades hídricas del cultivo. Desde la emergencia y hasta la madurez fisiológica del tubérculo, se registró diariamente la temperatura máxima y mínima del aire (Figura 1) con termómetro de columna de mercurio marca Taylor® modelo 5458; con estos datos se obtuvo la temperatura media. La temperatura máxima fluctúo entre 27 y 45 °C, y la mínima entre 1 y 11 °C (Figura 1). Se observaron temperaturas superiores a 30 °C e inferiores a 10 °C en la estación de crecimiento; las cuales tuvieron duración corta a través de un día.
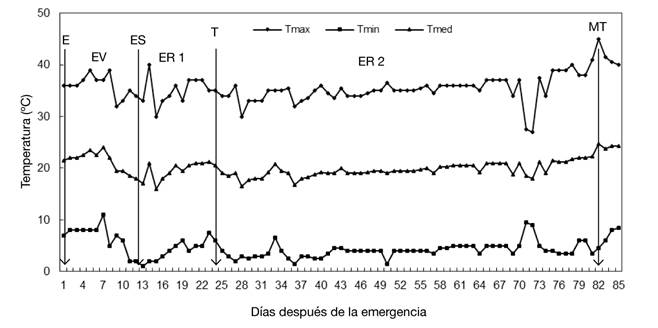
Figura 1 Temperatura del aire registrada durante la estación de crecimiento en condiciones de invernadero. Chapingo, México. Ciclo otoño-invierno, 2008-2009. Las flechas indican las fases de emergencia E, inicio de estoloneo (ES), tuberización (T) y madurez fisiológica del tubérculo (MT), y los intervalos entre éstas a las etapas vegetativa (EV), reproductiva inicial (ER 1) y reproductiva final (ER 2). Valores promedio de cuatro variedades.
Con el método residual clásico, y al considerar una temperatura base de 6 °C (Cao & Tibbits, 1995), se determinó el número de grados día acumulados (GD) desde la emergencia hasta el inicio de cada fase fenológica. A partir de esta información se determinó la duración de la etapa fenológica en cada genotipo estudiado.
La fase de emergencia (E) se determinó cuando 50 % de las plantas de la unidad experimental habían emergido; el inicio de estoloneo (ES), cuando se observó el primer estolón en el tallo principal. El inicio de tuberización o de crecimiento del tubérculo (T) se consideró cuando engrosó el ápice del estolón. La madurez fisiológica del tubérculo (MT), se estableció visualmente con base en la pérdida del color verde del follaje o inicio de la senescencia de éste.
La etapa vegetativa (EV) fue el periodo transcurrido desde la emergencia hasta el inicio del estoloneo, durante el cual ocurre el crecimiento de la parte aérea y establecimiento del sistema radical en la parte subterránea. La etapa reproductiva inicial (ER 1), caracterizada por la generación y crecimiento de estolones, ocurrió entre el inicio de estoloneo y el de tuberización. La etapa reproductiva final (ER 2) sucedió desde el inicio de la tuberización hasta la madurez fisiológica del tubérculo, y en ella ocurrió el crecimiento de éste.
En la cosecha se cuantificó el número de tubérculos por planta (NTP) con diámetro mayor a 5 mm y el peso de tubérculos por planta (PTP, en g). El peso promedio de tubérculo por planta (PPTP, en g) se obtuvo a partir del cociente PTP/NTP.
La materia seca de la parte aérea (MSA, en g), que incluyó hojas y tallos, así como la materia seca de la parte subterránea (MSR, en g), que comprendió raíz y parte del estolón, se determinaron en la cosecha a partir de dos plantas de la unidad experimental. Las estructuras se secaron durante 72 h a 75 °C en estufa marca Blue M® modelo POM-326F.
Cuantificación de carbohidratos solubles por cromatografía líquida de alta resolución (HPLC)
Obtención de muestras
Durante el desarrollo del tubérculo se realizaron cuatro muestreos; el primero 21 días después del inicio de la tuberización y los otros tres en intervalos de 14 ± 2 días, donde el último coincidió con la madurez fisiológica. Las muestras se realizaron a partir de dos plantas establecidas en una unidad experimental. Para las determinaciones del contenido de azúcares se seleccionó el tubérculo de mayor tamaño, del cual se consideraron dos duplicados (Cuadro 1). Cada muestra consistió de 5 ± 1 g de la parte central del tubérculo, misma que se colocó en nitrógeno líquido (-196 °C) hasta su análisis.
Cuadro 1 Peso de tubérculos de cuatro variedades de papa con diferente estado de madurez. Chapingo, México.
| Variedad | Estado de madurez del tubérculo | |||||||
|---|---|---|---|---|---|---|---|---|
| Uno | Dos | Tres | Cuatro | |||||
| DDT z | Peso y (g) | DDT | Peso (g) | DDT | Peso (g) | DDT | Peso (g) | |
| Alpha | 25 | 14 | 40 | 32 | 54 | 45 | 67 | 69 |
| Atlantic | 9 | 14 | 23 | 29 | 37 | 39 | 50 | 66 |
| Mondial | 26 | 21 | 40 | 47 | 54 | 67 | 67 | 84 |
| Vivaldi | 24 | 8 | 38 | 9 | 52 | 26 | 64 | 41 |
| Media | 21 | 14 | 35 | 29 | 49 | 44 | 62 | 65 |
zDDT: días después del inicio de la tuberización; y promedio de 2 tubérculos.
Extracción de carbohidratos solubles
Las muestras congeladas de tubérculo se molieron individualmente en una licuadora, con 20 mL de etanol grado HPLC a 70 %, y llevadas a temperatura de ebullición durante 5 min. El extracto se decantó y al residuo se le realizó una segunda extracción con 10 mL de etanol. Al final, los extractos se mezclaron. Posteriormente, la mezcla se centrifugó por 15 min a 4,000 x g en una centrífuga HERLE modelo Z230A (Labnet International, Inc. Edison, NJ. U.S.A), se filtró con papel Whatman número 4 y se le agregó manitol en concentración de 4 mg·mL-1 como estándar interno. Finalmente, el extracto se aforó a 25 mL con el mismo solvente empleado para la extracción.
Del extracto aforado se tomaron 10 mL para concentrar a sequedad con presión reducida a 40 °C, en un rotoevaporador BUCHI R-215 (Switzerland). El residuo se resuspendió en 2 mL de agua grado HPLC para pasarlo a través de una columna que contenía resinas intercambiadoras de iones, una básica (0.5 mL de Dowex - 1 x 8 - Fluka) y la otra ácida (0.5 mL de Dowex - 50W X8 - Fluka), en proporción 1:1 (p/p). La muestra purificada se pasó por un acrodisco de nylon de 0.45 μm con el fin de colectar la muestra en un vial de 1.5 mL.
Análisis de azúcares en HPLC
Se utilizó un equipo de cromatografía líquida de alta resolución, serie 200 Perkin Elmer (Boston Ma. U.S.A), compuesto por un auto muestreador, bomba cuaternaria, degasificador, detector de índice de refracción y horno para columnas. El sistema se operó con el programa computacional TotalChrom versión 6.2.1. Se usó una columna Rezex, RCM-monosaccharide Ca+2 (8 %) de 30 cm de longitud y 7.8 mm de diámetro. Como fase móvil se empleó agua grado HPLC. La velocidad de flujo fue de 0.6 mL·min-1, el volumen de muestra inyectado de 20 μL y el tiempo de corrida de 25 min. La temperatura de la columna se mantuvo a 85 °C. Se elaboraron curvas patrón con estándares comerciales de sacarosa, glucosa y fructosa (Sigma, MN), en concentraciones de 0.5, 1.0, 2.5 y 5 mg·mL-1 (Morales-Fernández et al., 2015; Rodríguez-Saona & Wrolstad, 1997). Cada punto de las curvas se inyectó al menos tres veces para obtener los datos del área asociada a cada concentración; con esta información se obtuvieron las ecuaciones de regresión para cada azúcar.
El experimento se estableció bajo el diseño de bloques completos al azar con tres repeticiones, y la unidad experimental consistió de cuatro contenedores (bolsas) llenas con sustrato. Con los datos obtenidos se realizó un análisis de varianza y de regresión. Además, se aplicaron pruebas de comparación de medias de Tukey (P ≤ 0.05); para ello se empleó el paquete Statistical Analysis System (SAS, 2004).
Resultados y discusión
Fenología del cultivo
El desarrollo fenológico del cultivo se adecuó en condiciones de invernadero. La temperatura media observada de 20 °C coincidió con la óptima para el cultivo de papa (Rykaczewska, 2015); lo que permitió el desarrollo y acumulación de materia seca de tubérculos adecuados, sin observarse tuberización secundaria y defectos fisiológicos en el tubérculo, característico de cultivos desarrollados en altas temperaturas (Rykaczewska, 2015).
Las variedades de papa tuvieron comportamiento diferencial (P ≤ 0.05) con respecto al ciclo biológico. Alpha y Vivaldi fueron contrastantes, ya que la última requirió 11 % menos GD a la madurez fisiológica del tubérculo; mientras que Atlantic y Mondial tuvieron duración intermedia (1194 GD). Excepto en Atlantic, las variedades iniciaron el estoloneo (200 GD) y tuberización (309 GD) en el mismo momento, y tuvieron comportamiento estadísticamente similar en las etapas vegetativas (200 GD) y reproductiva inicial (109 GD). Sin embargo, en la fase reproductiva final fue en la que ocurrieron mayores variaciones (P ≤ 0.05) que repercutieron en la disminución del ciclo biológico de las variedades (Cuadro 2), situación similar a la reportada por Morales-Fernández et al. (2011), debido a las diferencias genotípicas y ambientales en el desarrollo del cultivo (Rykaczewska, 2015).
Cuadro 2 Requerimiento de grados día para alcanzar las principales fases y etapas fenológicas de cuatro variedades de papa cultivadas en condiciones de invernadero. Chapingo, México.
| Variedad | Etapas (GD) | |||
|---|---|---|---|---|
| ES | EV | ER 1 | ER 2 | |
| Alpha | 198 az | 198 a | 109 b | 923 a |
| Atlantic | 122 b | 122 b | 307 a | 760 c |
| Mondial | 203 a | 203 a | 96 b | 900 b |
| Vivaldi | 199 a | 199 a | 123 b | 771 c |
| DMSH | 23 | 23 | 40 | 17 |
1GD: grados día; ES: inicio de estoloneo; T: inicio de tuberización; MT: madurez fisiológica del tubérculo; EV: etapa vegetativa; ER 1: etapa reproductiva inicial; ER 2: etapa reproductiva final; DMSH: diferencia mínima significativa honesta.
zMedias con la misma letra dentro de cada columna no difieren estadísticamente (Tukey, P ≤ 0.05).
Rendimiento y sus componentes, y contenido de materia seca
Se presentaron temperaturas altas (Figura 1) en la etapa reproductiva inicial (ER 1); lo que, de acuerdo con Struik, Haverkort, Vreugdengil, Bus, y Dankert (1990), pudo ser benéfico para promover la ramificación e incremento del número de estolones, condición que incrementa directamente el número de tubérculos y estos, a su vez, repercuten en el rendimiento (Haverkort, van de Waart, & Bodlaender, 1990).
La variedad Mondial tuvo mayor rendimiento por planta y superó (P ≤ 0.05) a Vivaldi, Alpha y Atlantic en 75, 31 y 31 %, respectivamente (Cuadro 3). Este parámetro está asociado al mayor peso promedio del tubérculo, ya que, junto con el número de tubérculos por planta, son los principales componentes que definen el rendimiento de papa (Zvomuya & Rosen, 2002).
Las variedades Alpha y Atlantic produjeron el mayor (P ≤ 0.05) número de tubérculos por planta, 36 % más que el resto de las variedades; aunque su rendimiento fue menor (P ≤ 0.05) que Mondial, debió al menor peso promedio de tubérculos por planta. Estos resultados concuerdan con los de Walworth y Carling (2002), quienes detectaron que el incremento del número de tubérculos por planta disminuye su tamaño final.
Cuadro 3 Rendimiento y sus componentes, y acumulación de materia seca en cuatro variedades de papa cultivadas en condiciones de invernadero. Chapingo, México.
| Variedad | NTP 1 | PTP (g) | PPTP (g) | MSA (g) | MSR (g) |
|---|---|---|---|---|---|
| Alpha | 10 az | 247 b | 24 b | 22 a | 3 b |
| Atlantic | 12 a | 277 b | 22 b | 28 a | 9 a |
| Mondial | 7 b | 380 a | 61 a | 29 a | 7 a |
| Vivaldi | 7 b | 95 c | 11 c | 7 b | 1 b |
| DMSH | 2 | 44 | 7 | 8 | 3 |
1NTP: número de tubérculos por planta; PTP: peso fresco de tubérculos por planta; PPTP: peso promedio de tubérculo por planta; MSA: materia seca de la parte aérea; MSR: materia seca de la parte subterránea; DMSH: diferencia mínima significativa honesta.
zMedias con la misma letra dentro de cada columna no difieren estadísticamente (Tukey, P ≤ 0.05).
El contenido de biomasa de la parte aérea fue similar (P ≤ 0.05) en las variedades Alpha, Atlantic y Mondial, las cuales superaron 73 % a Vivaldi; mientras que la biomasa de la parte subterránea fue 75 % mayor en Atlantic y Mondial con respecto de Alpha y Vivaldi (Cuadro 3). El contenido alto de biomasa aérea y subterránea de Mondial se asoció a un mayor peso de tubérculos por planta. Resultados similares son los de Morales-Fernández et al. (2011), quienes indican la importancia de ambos caracteres sobre el rendimiento de papa.
A pesar de que Alpha y Atlantic tuvieron contenido alto de materia seca aérea, el comportamiento de su rendimiento fue diferente al de Mondial, lo que pudo ser debido a un desbalance en la competencia entre el crecimiento de ambas partes y por el mayor número de tubérculos por planta, lo que provocó menor desarrollo y tamaño de tubérculos (Walworth & Carling, 2002) y, por lo tanto, generó menor rendimiento.
Contenido de azúcares en tubérculos de papa
Durante su crecimiento y desarrollo, el tubérculo es el principal sitio de demanda de fotoasimilados, los cuales son traslocados desde los sitios de síntesis (hojas), principalmente en forma de sacarosa (Fernie, Willmitzer, & Trethewey, 2002). En la presente investigación, el contenido promedio de sacarosa, glucosa, fructosa y azúcares totales fue, respectivamente, 63, 65, 38 y 63 % mayor en 21 que en 62 días después del inicio de tuberización y, en general, la cantidad de azúcares solubles disminuyó conforme transcurrió el crecimiento de los tubérculos (Cuadro 4).
Cuadro 4 Contenido de azúcares en tubérculos de papa en diferente estado de madurez. Chapingo, México.
| Estado de madurez | DDT 1 | Concentración de azúcares (mg·g -1 PF) | |||
|---|---|---|---|---|---|
| Sacarosa | Glucosa | Fructosa | Totales | ||
| EM 1 | 21 | 7.65 az | 2.22 a | 0.37 a | 10.25 a |
| EM 2 | 35 | 5.44 b | 1.19 b | 0.25 b | 6.89 b |
| EM 3 | 49 | 3.29 c | 0.47 c | 0.20 b | 3.96 c |
| EM 4 | 62 | 2.81 c | 0.77 c | 0.23 b | 3.82 c |
| DMSH | 1.01 | 0.31 | 0.10 | 1.26 | |
1DTT: días después del inicio de tuberización; PF: peso en fresco; DMSH: diferencia mínima significativa honesta.
zMedias con la misma letra dentro de cada columna no difieren estadísticamente (Tukey, P ≤ 0.05).
Otras investigaciones coinciden con la presente al detectar que las concentraciones mayores de sacarosa, glucosa y fructosa se presentan en tubérculos inmaduros (Knowles, Pavek, Knowles, & Holden, 2008; Kumar & Ezekiel, 2006), y que la concentración más baja de sacarosa ocurre durante la madurez (Knowles et al., 2009; Sabba et al., 2007), como ocurrió en el presente estudio en EM4 debido a la reducción del suministro de fotosintatos a consecuencia de la senescencia del follaje de las plantas, como lo sugiere Kumar y Ezekiel (2006). Así, la concentración baja de este azúcar puede ser utilizado como un indicador de cosecha (Chen, Zhang, Miao, & Asakura, 2010); aunque debido a lo complicado del registro y costo, es difícil considerar su uso en sistemas de producción convencionales, ante la posibilidad de detectar este estado mediante la senescencia del follaje.
Al analizar el comportamiento de los azúcares solubles de acuerdo con el estado de madurez del tubérculo (Figura 2), se observó concentración mayor (P ≤ 0.05) en el inicio de crecimiento en comparación con la madurez fisiológica, tanto en sacarosa (10.44 a 1.81 mg·g-1) como en azúcares totales (13.40 a 2.16 mg·g-1). En este periodo, las concentraciones de glucosa y fructosa fueron menores de 3.30 y 0.60 mg·g-1, respectivamente. Los tubérculos en estado de madurez uno (EM 1) tuvieron el contenido mayor de azúcares solubles que aquellos cosechados en madurez fisiológica (EM 4).
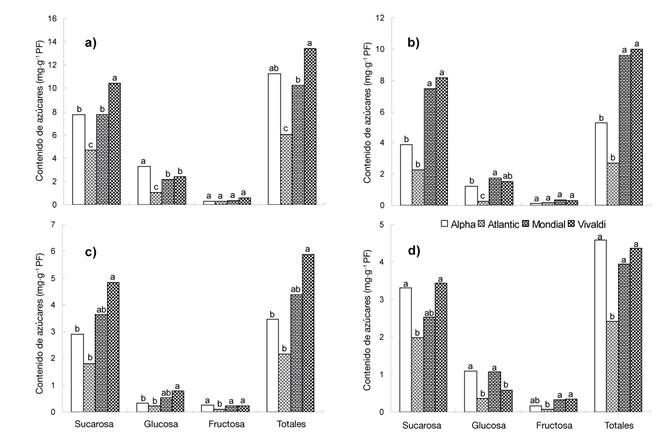
Figura 2 Contenido de azúcares solubles en tubérculos de papa con distintos estado de madurez: a) uno (21 DDT), b) dos (35 DDT), c) tres (49 DDT) y d) cuatro (62 DDT). Chapingo, México. Ciclo otoño-invierno, 2008-2009. Medias con la misma letra dentro de cada tipo de azúcar no difieren estadísticamente (Tukey, P ≤ 0.05).
Los tubérculos de la variedad Vivaldi acumularon más sacarosa (10.44 mg·g-1) y azúcares totales (13.40 mg·g-1) que el resto de los genotipos estudiados (Figura 2a). A pesar de estas diferencias, se apreció que, a medida que se alcanzó la madurez, las cantidades de sacarosa, glucosa, fructosa y azúcares totales en el tubérculo fueron inferiores a 5 mg·g-1 (Figura 2d). Dichos valores se encuentran dentro del intervalo reportado por Knowles et al. (2009) para el procesamiento y condiciones de manejo durante el almacenamiento.
La interacción entre variedades y estados de madurez del tubérculo indicó que Vivaldi exhibió mayor variación (P ≤ 0.05) en la cantidad de azúcares al cambiar de un estado de madurez a otro (Cuadro 5); asimismo, en EM 1 y EM 2 tuvo 80 y 81 % mayor contenido de sacarosa y azúcares totales que Atlantic. En el caso de Alpha, en EM 1 tuvo mayor cantidad de glucosa (92 %) que Atlantic en EM 2 y EM 3. La variedad Vivaldi, en EM 1, tuvo mayor contenido de fructosa (88 %) que Atlantic en EM 4, lo cual se explica por el desarrollo específico de los genotipos.
Cuadro 5 Comparación de medias de la interacción variedad por estado de madurez del tubérculo de papa, en condiciones de invernadero, sobre el contenido de azúcares. Chapingo, México.
| Variedad | EM 1 | Concentración de azúcares (mg·g -1 PF) | ||||
|---|---|---|---|---|---|---|
| DDT | Sacarosa | Glucosa | Fructosa | Totales | ||
| Alpha | 1 | 25 | 7.71 abz | 3.26 a | 0.30 bcd | 11.27 ab |
| Alpha | 2 | 40 | 3.90 de | 1.22 def | 0.15 def | 5.27 de |
| Alpha | 3 | 54 | 2.89 de | 0.32 hi | 0.25 bcde | 3.47 de |
| Alpha | 4 | 67 | 3.31 de | 1.09 defg | 0.17 cdef | 4.58 de |
| Atlantic | 1 | 9 | 4.71 cde | 1.05 defgh | 0.29 bcd | 6.06 cd |
| Atlantic | 2 | 23 | 2.26 de | 0.26 i | 0.19 bcdef | 2.72 de |
| Atlantic | 3 | 37 | 1.81 e | 0.24 i | 0.10 ef | 2.16 e |
| Atlantic | 4 | 50 | 1.98 de | 0.36 ghi | 0.07 f | 2.41 de |
| Mondial | 1 | 26 | 7.73 ab | 2.17 bc | 0.35 bc | 10.26 ab |
| Mondial | 2 | 40 | 7.48 abc | 1.75 bcd | 0.36 b | 9.60 bc |
| Mondial | 3 | 54 | 3.63 de | 0.53 fghi | 0.22 bcdef | 4.37 de |
| Mondial | 4 | 67 | 2.53 de | 1.07 defgh | 0.33 bcd | 3.94 de |
| Vivaldi | 1 | 24 | 10.44 a | 2.39 b | 0.57 a | 13.40 a |
| Vivaldi | 2 | 38 | 8.15 a | 1.53 cde | 0.31 bcd | 9.99 ab |
| Vivaldi | 3 | 52 | 4.84 bcd | 0.80 efghi | 0.23 bcdef | 5.87 cde |
| Vivaldi | 4 | 64 | 3.44 de | 0.58 fghi | 0.35 bc | 4.37 de |
| DMSH | 2.97 | 0.75 | 0.18 | 3.76 | ||
1EM: estado de madurez; DDT: días después de tuberización; DMSH: diferencia mínima significativa honesta.
zMedias con la misma letra dentro de cada columna no difieren estadísticamente (Tukey, P ≤ 0.05).
La variación mostrada en el contenido de azúcares entre variedades, aún en el mismo estado de madurez, refleja la habilidad que tienen algunos genotipos para acumular carbohidratos durante el desarrollo del cultivo; aunque este comportamiento depende del genotipo (Park et al., 2009), estrés hídrico (Bethke, Sabba, & Bussan, 2009) y nutrimental (Ilin et al., 1997), y de la temperatura (Rykaczewska, 2015) durante el crecimiento.
En el presente estudio se observaron temperaturas extremas puntuales con duración corta durante el día; las cuales no lograron generar un estrés generalizado que limitase el desarrollo de los cultivares, a excepción de la variedad Atlantic que produjo bajas concentraciones de azúcares solubles en los diferentes estados de madurez, aun cuando su rendimiento fue menor (P ≤ 0.05) que el de la variedad más sobresaliente. Estos resultados son similares a los reportados por Morales-Fernández et al. (2015) durante la brotación del tubérculo.
La presencia de la variación genotípica, demostrada especialmente a través del comportamiento de Atlantic, sugiere la posibilidad de realizar selección de variedades de acuerdo con las condiciones de producción. Asimismo, al considerar el origen de las variedades empleadas en el presente estudio, las cuales no fueron desarrolladas para su establecimiento en invernadero, se observa que el factor ambiental influyó en su desarrollo y, por lo tanto, en su rendimiento (Rykaczewska, 2015).
Los niveles bajos de azúcares totales y sobre todo de sacarosa, mostrados en algunas variedades con EM 3, sugieren que pudieron ser cosechadas en dicha condición. Según Knowles et al. (2009) y Sabba et al. (2007), los valores menores de sacarosa se encuentran durante la madurez fisiológica del tubérculo, criterio que podría ser utilizado como un indicador de cosecha, independientemente de las características fenotípicas del follaje de las plantas.
El contenido de azúcares solubles presentó asociación significativa alta con el peso del tubérculo (Figura 3). En promedio de variedades, la sacarosa (Figura 3a), azúcares reductores (glucosa y fructosa; Figura 3b) y totales (Figura 3c) disminuyeron a medida que el tubérculo aumentó de peso; mientras que de manera individual, las variedades estudiadas mostraron la misma tendencia al asociar los tres azúcares con el crecimiento del tubérculo. Estos resultados concuerdan con los de Kumar y Ezekiel (2006) al haber detectado una asociación negativa entre el contenido de azúcares solubles con el peso del tubérculo.
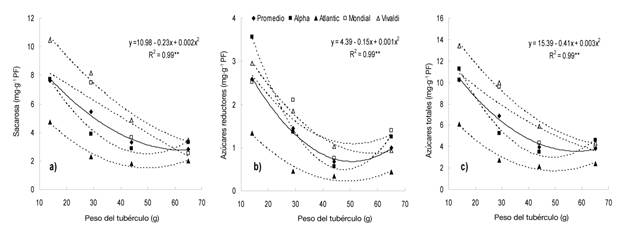
Figura 3 Relación entre el contenido de sacarosa (a), azúcares reductores (b) y totales (c) con el tamaño del tubérculo en cuatro variedades de papa bajo condiciones de invernadero. Chapingo, México. Ciclo otoño-invierno, 2008-2009. El modelo de regresión representa al promedio de variedades.
Stark y Love (2003) indican que el contenido mayor de azúcares solubles en los tubérculos se presenta cuando estos son de peso bajo o tienen grado de inmadurez alto, como sucedió con los de EM 1 en este trabajo, debido a que la tasa de transporte de azúcares desde las hojas al tubérculo supera a la de conversión a almidón. Dicha condición puede cambiar con el crecimiento del tubérculo o cuando éste alcanza la madurez fisiológica y el ambiente no es un factor condicionante (Knowles et al., 2009).
Las concentraciones de azúcares (sacarosa, glucosa, fructosa y totales) registradas en la madurez fisiológica de los tubérculos producidos en condiciones de invernadero fueron inferiores a 5 mg·g-1 (Cuadro 5), valores que se encuentran dentro del intervalo reportado por Knowles et al. (2009) para el procesamiento y condiciones de manejo durante el almacenamiento. Para azúcares reductores en la madurez, los valores obtenidos se encontraron en el rango reportado por Kumar y Ezekiel (2006) y Sabba et al. (2007), que fue de 0.04 a 0.40 %. Sin embargo, de acuerdo con Vázquez-Carrillo, Santiago-Ramos, Ybarra-Moncada, Rubio-Covarrubias, y Cadena-Hinojosa (2013), el consumo de estos tubérculos debería ser en fresco o para cocción. Lo anterior debido a que durante el procesamiento de frituras la reacción temperatura-aminoácidos-azúcares reductores de la papa produce el oscurecimiento.
Conclusiones
La duración más larga del ciclo biológico de las variedades no mostró asociación con un rendimiento mayor de tubérculos en términos de peso. El componente del rendimiento que más contribuyó en la producción fue el peso promedio de tubérculo por planta, expresado con mayor magnitud en una variedad de ciclo intermedio.
El contenido de azúcares solubles fue modificado por el estado de madurez y el peso del tubérculo. La concentración mayor se presentó a los 21 días después del inicio de tuberización (EM 1). Esta variación también fue afectada por el componente genético, ya que hubo diferenciación en las concentraciones de sacarosa, glucosa, fructosa y azúcares totales, lo cual fue independiente del estado de madurez del tubérculo.
Se detectó una relación inversa entre el peso de tubérculo y los contenidos de sacarosa, azúcares reductores (glucosa y fructosa) y totales.
Las bajas concentraciones de azúcares (sacarosa, glucosa y fructuosa) en el tubérculo puede considerarse como un indicador de la madurez, aunque los valores de éstas no pueden ser generalizados. Sin embargo, la dificultad y costo de dichas determinaciones hacen impráctico este procedimiento.
References
Bethke, P. C., Sabba, R., & Bussan, A. J. (2009). Tuber water and pressure potentials decrease and sucrose contents increase in response to moderate drought and heat stress. American Journal of Potato Research, 86(6), 519-532. doi: 10.1007/s12230-009-9109-8 [ Links ]
Cao, W., & Tibbitts, T. W. (1995). Leaf emergence on potato stems in relation to thermal time. Agronomy Journal, 87(3), 474-477. doi: 10.2134/agronj1995.00021962008700030013x [ Links ]
Cenzano, A., Abdala, G., & Hause, B. (2007). Cytochemical immuno-localization of allene oxide cyclase, a jasmonic acid biosynthetic enzyme, in developing potato stolons. Journal of plant physiology, 164(11), 1449-1456. doi: 10.1016/j.jplph.2006.10.007 [ Links ]
Chen, J. Y., Zhang, H., Miao, Y., & Asakura, M. (2010). Nondestructive determination of sugar content in potato tubers using visible and near infrared spectroscopy. Japan Journal of Food Engineering, 11(1), 59-64. Retrieved from http://www.jsfe.jp/journal/PDF/vol11_1_7.pdf [ Links ]
Fernie, A. R., Willmitzer, L., & Trethewey, R. N. (2002). Sucrose to starch: a transition in molecular plant physiology. Trends in Plant Science, 7(1), 35-41. doi: 10.1016/S1360-1385(01)02183-5 [ Links ]
Hancock, R. D., Roberts, A. G., & Viola, R. (2008). A role for symplastic gating in the control of the potato tuber life cycle. Plant signaling & behavior, 3(1), 27-29. doi: 10.4161/psb.3.1.4813 [ Links ]
Hannapel, D. J., Chen, H., Rosin, F. M., Banerjee, A. K., & Davies, P. J. (2004). Molecular controls of tuberization. American journal of potato research, 81(4), 263-274. doi: 10.1007/BF02871768 [ Links ]
Haverkort, A. J., van de Waart, M., & Bodlaender, K. B. A. (1990). The effect of early drought stress on numbers of tubers and stolons of potato in controlled and field studies. Potato Research 33(1), 89-96. doi: 10.1007/BF02358133 [ Links ]
Ilin, Z., Durovka, M., & Markovic, V. (1997). Effect of fertility and irrigation on sugar content in potato tubers. Acta Horticultorae, 462, 303-309. Retrieved from http://www.actahort.org/books/462/462_44.htm [ Links ]
Knowles, N. R., Driskill, E. P., & Knowles, L. O. (2009). Sweetening responses of potato tubers of different maturity to conventional and non-conventional storage temperature regimes. Postharvest Biology and Technology, 52(1), 49-61. doi: 10.1016/j.postharvbio.2008.08.015 [ Links ]
Knowles, N. R., Pavek, M. J., Knowles, L. O., & Holden, Z. (2008). Developmental profiles and postharvest behavior of long-season processing cultivars. In: Proceedings of the 47th annual Washington State potato conference (pp. 5-7). [ Links ]
Kolbe, H., & Stephan-Beckmann, S. (1997). Development, growth and chemical composition of the potato crop (Solanum tuberosum L.). II. Tuber and whole plant. Potato Research, 40(2), 135-153. doi: 10.1007/BF02358240 [ Links ]
Kooman, P. L., Fahem, M., Tegera, P., & Haverkort, A. J. (1996). Effects of climate on different potato genotypes 2. Dry matter allocation and duration of the growth cycle. European Journal of Agronomy, 5(3), 207-217. doi: 10.1016/S1161-0301(96)02032-1 [ Links ]
Kumar, D., & Ezekiel, R. (2006). Developmental changes in sugars and dry matter content of potato tuber under sub-tropical climates. Scientia horticulturae, 110(2), 129-134. doi: 10.1016/j.scienta.2006.06.022 [ Links ]
Lynch, D. R., Kozub, G. C., & Kawchuk, L. M. (2001). The relationship between yield, mainstem number, and tuber number in five maincrop and two early-maturing cultivars. American journal of potato research, 78(2), 83-90. doi: 10.1007/BF02874763 [ Links ]
Milton, P. J., & Allen, S. D. (1995). Breeding Field Crops. Iowa, USA: Iowa State University Press, Ames. [ Links ]
Morales-Fernández, S. D., Mora-Aguilar, R., Salinas-Moreno, Y., Rodríguez-Pérez, J. E., Colinas-León, M. T., & Lozoya-Saldaña, M. T. (2015). Growth, yield and sugar content of potato tubers at different physiological ages. Revista Chapingo Serie Horticultura, 21(2), 129-146. doi: 10.5154/r.rchsh.2014.06.031 [ Links ]
Morales-Fernández, S. D., Mora-Aguilar, R., Rodríguez-Pérez, J. E., Salinas-Moreno, Y., Colinas-León, M., & Lozoya-Saldaña, H. (2011). Desarrollo y rendimiento de papa en respuesta a la siembra de semilla-tubérculo inmadura. Revista Chapingo. Serie horticultura, 17(1), 67-75. doi: 10.5154/r.rchsh.2011.17.010 [ Links ]
Park, S. W., Jeon, J. H., Kim, H. S., Hong, S. J., Aswath, C., & Joung, H. (2009). The effect of size and quality of potato microtubers on quality of seed potatoes in the cultivar ‘Superior’. Scientia Horticulturae, 120(1), 127-129. doi: 10.1016/j.scienta.2008.09.004 [ Links ]
Rodríguez-Saona, L. E., & Wrolstad, R. E. (1997). Influence of potato composition on chip color quality. American Potato Journal, 74(2), 87-106. doi: 10.1007/BF02851555 [ Links ]
Rykaczewska, K. (2015). The Effect of High Temperature Occurring in Subsequent Stages of Plant Development on Potato Yield and Tuber Physiological Defects. American Journal of Potato Research, 92(3), 1-11. doi: 10.1007/s12230-015-9436-x [ Links ]
Sabba, R. P., Bussan, A. J., Michaelis, B. A., Hughes, R., Drilias, M. J., & Glynn, M. T. (2007). Effect of planting and vine-kill timing on sugars, specific gravity and skin set in processing potato cultivars. American Journal of Potato Research, 84(3), 205-215. doi: 10.1007/BF02986270 [ Links ]
Stark, J. C., & Love, S. l. (2003). Tuber quality. In: Stark, J. C., & Love, S. L. (Eds.), Potato production systems (pp. 329-343). Idaho: Center of Potato Research and Education. [ Links ]
Statistical Analysis System (SAS Institute). (2004). SAS/STAT 9.1 user’s guide. Cary, NC, USA: Author. [ Links ]
Struik, P. C., Haverkort, A. J., Vreugdengil, D., Bus, C. B., & Dankert, R. (1990). Manipulation of tuber-size distribution of a potato crop. Potato Research, 33(4), 417-432. doi: 10.1007/BF02358019 [ Links ]
Suttle, J. (2008). Symposium introduction: enhancing the nutritional value of potato tubers. American Journal of Potato Research, 85(4), 266-266. doi: 10.1007/s12230-008-9033-3 [ Links ]
Thompson, A. L., Love, S. L., Sowokinos, J. R., Thornton, M. K., & Shock, C. C. (2008). Review of the sugar end disorder in potato (Solanum tuberosum L.). American Journal of Potato Research, 85(5), 375-386. doi: 10.1007/s12230-008-9034-2 [ Links ]
Vázquez-Carrillo, M. G., Santiago-Ramos, D., Ybarra-Moncada, M. C., Rubio-Covarrubias, Á.O., & Cadena-Hinojosa, M. A.(2013). Variables fisicoquímicas y calidad de fritura de clones de papa desarrollados para los Valles Altos de México. Agrociencia, 47(1), 1-12. Retrieved from http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S1405-31952013000100005 [ Links ]
Viola, R., Pelloux, J., van der Ploeg, A., Gillespie, T., Marquis, N., Roberts, A. G., & Hancock, R. D. (2007). Symplastic connection is required for bud outgrowth following dormancy in potato (Solanum tuberosum L.) tubers. Plant Cell Environment, 30(8), 973-983. doi: 10.1111/j.1365- 3040.2007.01692.x [ Links ]
Walworth, J. L., & Carling, D. E. (2002). Tuber initiation and development in irrigated and non-irrigated potatoes. American Journal of Potato Research, 79(6), 387-395. doi: 10.1007/BF02871683 [ Links ]
Zhang, H., Hou, J., Liu, J., Xie, C., & Song, B. (2014). Amylase analysis in potato starch degradation during cold storage and sprouting. Potato research, 57(1), 47-58. doi: 10.1007/s11540-014-9252-6 [ Links ]
Zvomuya, F., & Rosen, C. J. (2002). Biomass partitioning and nitrogen use efficiency of ‘Superior’ potato following genetic transformation for resistance to Colorado potato beetle. Journal of the American Society for Horticultural Science, 127(4), 703-709. Retrieved from http://journal.ashspublications.org/content/127/4/703.full.pdf+html?sid=69371c99-6a40-4875-bc0e-91659a33821b [ Links ]
Recibido: 23 de Noviembre de 2016; Aprobado: 14 de Octubre de 2017
 This is an open-access article distributed under the terms of the Creative Commons Attribution License
This is an open-access article distributed under the terms of the Creative Commons Attribution License
