











 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroduction
The genus Physalis has great cultural, economic and biological importance in Mexico. It has been present in the Mexican diet since pre-Columbian times and its use continues to this day. It is mainly consumed fresh, although it is also industrialized, both for the domestic market and for export. Some species of this genus have medicinal and ornamental uses, which are little known and exploited (Santiaguillo-Hernández & Blas-Yáñez, 2009). In particular, the tomatillo grown in Mexico is Physalis ixocarpa Brot. ex Horm. (Santiaguillo-Hernández, Cedillo-Portugal, & Cuevas-Sánchez, 2010). It can be cultivated in both irrigated and rain-fed systems, and practically all year round (spring-summer cycle and autumn-winter cycle). Also, the tomatillo is grown in diverse climatic conditions, which suggests great adaptability on the part of the species (Peña-Lomelí & Sanatiaguillo-Hernández, 1999; Santiaguillo-Hernández et al., 2010).
Among vegetables, the tomatillo ranks sixth in terms of area cultivated in Mexico. From 2006 to 2015, an average of 49,177 ha of planted area per year was maintained (Sistema de Información Agrícola y Pesquera - Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación [SIAP-SAGARPA], 2015). The importance acquired by this crop is due to the increase in per capita consumption (4.5 kg) at the national level, as well as an increase in its exports to the United States and Canada, the main market for green tomatoes, although there are also limited purchases by The Netherlands and the United Kingdom (Peña-Lomelí et al., 2002). The national average yield (16.10 t∙ha-1) is considered low in relation to the productive potential of the crop estimated at 40 t∙ha-1. The causes of the low yield may be the use of varieties with low productive potential, inefficient production techniques, use of low quality seed and inefficient pest and disease control (Peña-Lomelí, Ponce-Valerio, Sánchez-del Castillo, & Magaña-Lira, 2014; Peña-Lomelí & Sanatiaguillo-Hernández, 1999; SIAP-SAGARPA, 2015).
In tomatillo there is self-incompatibility, which is given by two independent loci with multiple alleles. Pollen is incompatible when one or more alleles are present in both the pollen grain and the style. In the process between pollination and fertilization, pollen usually does not germinate, but if it does, the pollen tube cannot penetrate through the stigma (Pandey, 1957). As a consequence, its breeding by hybridization based on inbred lines is complicated, especially if adequate pollination techniques are not available (Peña-Lomelí & Márquez-Sánchez, 1990). However, several studies indicate that breeding by interpopulation hybridization is promising for the tomatillo, since high intra and inter varietal heterosis values have been obtained (Peña-Lomelí, Molina-Galán, Márquez-Sánchez, & Sahagún-Castellanos, 1999; Peña-Lomelí et al., 1998; Sahagún-Castellanos, Gómez-Ruiz, & Peña-Lomelí, 1999; Santiaguillo-Hernández, Cervantes-Santana, & Peña-Lomelí, 2004).
Taking advantage of the heterosis in tomatillo implies, among other things, having an efficient technique for the realization of manual crosses. In this sense, due to self-incompatibility, cross-pollination (either through sibling crosses or from plant to plant) is facilitated, since in general it is not necessary to emasculate the flowers crossed (Peña-Lomelí & Márquez-Sánchez, 1990). In this regard, Santiaguillo-Hernández, Cervantes-Santana, Peña-Lomelí, Molina-Galán, and Sahagún-Castellanos (2005) mention that self-incompatibility in tomatillo is not complete and possibly lower in some varieties, while Mulato-Brito, Peña-Lomelí, Sahagún-Castellanos, Villanueva-Verduzco, and López-Reynoso (2007) report the existence of self-compatible mutants, which would imply the need to emasculate populations.
Tomatillo flowers are bisexual (perfect or hermaphroditic), solitary and emerge from the dichotomy of the branches. When the plant blossoms, it requires 30 to 32 °C. Temperatures above these values, during blossoming, can cause dehydration of the pollen tube, incomplete pollination and malformed fruits (Saray-Meza & Loya-Ramírez, 1977). In general, the flowers open before the dehiscence of the anthers occurs, between 8:00 and 12:00 in the day. Natural pollination in tomatillo is mostly performed by insects, mainly bees, although it can also be by the wind. Once the flower has been pollinated it closes and does not reopen, then it begins to wither and finally fall (Pérez-Grajales, Márquez-Sánchez, & Peña-Lomelí, 1998). At 55 days after pollination, tomatillo seed can be considered physiologically mature (Pérez-Camacho et al., 2012).
Since the tomatillo requires cross-pollination to form fruits and inside a greenhouse the wind circulation is minimal and insufficient to move the pollen, natural pollination is deficient (Santiaguillo-Hernández, Vargas-Ponce, Grimaldo-Juárez, Sánchez-Martínez, & Magaña-Lira, 2009). This implies that commercial greenhouse tomatillo production has not been efficient because, among other factors, pollination is difficult and consequently reduces yield (Peña-Lomelí et al., 2014; Ponce-Valerio et al., 2012). However, when manual crosses are performed for breeding, the greenhouse can be used, since low natural fertilization allows most of the fruits under development to be the product of manual pollination.
Therefore, when it comes to breeding, it is necessary to determine the appropriate time to perform manual pollination in controlled environments in order to achieve greater fruit and seed production. Some studies indicate that the manual fertilization of tomatillo in the greenhouse should be done when the flower buds are closed, since it has been found that the stigma is more receptive at this stage. By contrast, the anthers release pollen after the flower has opened. This technique is used in breeding programs, especially because it ensures the purity of the crosses that are made. However, precise knowledge is required to identify the ideal state of floral buds, without which its efficiency in pollinated flowers and seed produced is low (Peña-Lomelí et al., 1998; Santiaguillo-Hernández et al., 2005).
Based on the above, the aim of this research was to evaluate floral bud pollination times of two tomatillo varieties, in order to determine the optimal time to perform manual pollination efficiently in each of them to achieve the greatest fruit set and seed production, under the hypothesis that the best time will be after the floral bud is covered, and that there will be no difference between the varieties.
Materials and methods
The research work was carried out from October to December 2015 in the experimental field greenhouses of the Plant Science Department at the Universidad Autónoma Chapingo (UACh), Mexico (19° 29' 31.0" North latitude and 98° 52' 20.4" West longitude, at 2,250 masl). According to the Köppen classification modified by García (1988), the area has a Cb(Wo)(W)b(i´) climate, defined as temperate subhumid with summer rains, winter precipitation of less than 5 %, little thermal oscillation, cool summers and average annual precipitation of 636 mm. The mean annual temperature is between 12 and 18 °C, with less than 5 °C variation.
The varieties used in this study were Diamante and Manzano Tepetlixpa, both registered in the name of the Universidad Autónoma Chapingo (Servicio Nacional de Inspección y Certificación de Semillas [SNICS], 2016) with breeder's title 1583 and 1584, respectively. For each one, two high-yield maternal half-sib families (MHSF) were selected, with the purpose of generating inbreeding through sibling crosses and heterosis by interfamilial hybridization. The four families obtained were those studied in the present study.
Diamante is the result of selective breeding and arises from the hybridization of the CHF1 Chapingo and Puebla SM3 varieties. Its name derives from the brilliance of the color of its fruit and it has a short shelf life. It is a precocious plant of semi-erect bearing with elliptical, green leaves and average-sized flowers. Its fruits are large with three loci; the fruits are green with medium firmness, a completely closed calyx cover and yellowish-brown seeds. It is adapted to the climatic conditions of the Valles Altos region (Mexico, Tlaxcala, Puebla and Hidalgo) in the spring-summer cycle, and has a yield of 30 t∙ha-1 or more under irrigation conditions with mulch (Sánchez-Martínez & Peña-Lomelí, 2015).
On the other hand, Manzano Tepetlixpa is a variety obtained by selective breeding and its name derives from the town where the original material was collected (Tepetlixpa, municipality of the State of Mexico). It is a precocious plant, of erect bearing, with elliptical, green leaves and average-sized flowers. Its fruits have three loci; the fruits are large, sweet and yellow with medium firmness and a completely closed, ribbed calyx cover. It has medium-sized, yellow brown seeds. It is distributed mainly in Mexico and Morelos (Cuautla) and has a yield of 30 t∙ha-1 or more in irrigation and trellis conditions (Sánchez-Martínez & Peña-Lomelí, 2015).
Three factors were studied. The first was time (in days), with five levels, which were the five days in which pollination was performed daily. The second was variety, corresponding to two tomatillo populations (Diamante and Manzano Tepetlixpa). The third was family, which consisted of two levels within each variety (F1 and F2). This generated a full 5 x 2 x 2 factorial treatment design. Additionally, a control was included, which consisted of covered flowers without pollination, in order to verify that there were no self-compatible mutants within the populations studied (Mulato-Brito et al., 2007).
The experiment was established in the 2015 autumn-winter cycle, under a randomized complete design with four replicates. The experimental unit consisted of ten floral buds per treatment. The crop was grown under greenhouse conditions with hydroponics; as a substrate, fine-grained red tezontle was used and the nutrient solution used was that of Steiner (Steiner, 1984).
For the pollination process, ten floral buds of each experimental unit were covered with flat glassine bags measuring 4 cm wide by 7 cm long, which were closed by a fold with the aid of a number three clip. The criterion for choosing the flowers was that they were completely closed, but that the petals already had a yellow tone. According to the treatment assigned to each experimental unit, manual pollination was carried out on days one (corresponding to the time when the buds were covered), two, three, four and five. In the control, the flowers were left covered all the time.
To pollinate, pollen was collected (in a Petri dish 6 cm in diameter) from the open flowers of all the plants of the replicate for each family. Subsequently, the flowers corresponding to that day were identified and pollinated one by one. To do this, the bag was removed, then, with the aid of a number 0 camel hair brush, pollen was placed on the stigma and the flower was again covered with the bag. Once fertilization occurred, the corolla dried up and the development of the calyx began (ten days after pollination). At that time, the bag was removed to prevent the fruit from forming. The fruits in development were identified with number two hang tags.
The harvest was carried out 75 days after pollination. The variables evaluated are described below. Number of fruits (NF): fruit set per treatment was quantified. Fruit weight (FW, g): fruits obtained from each experimental unit were weighed on an OHAUS® 5 K digital balance. Total seed weight (TSW, mg) and weight of 100 seeds (W100S, mg) of each experimental unit. Both evaluations were made in the ecology laboratory of UACh’s Plant Science Department with a GRAM SVI® precision laboratory balance. Finally, the total number of seeds (TNS) was determined indirectly using the following equation:

In the case of the experimental units in which a small number of seeds was obtained, a direct count was made.
With the data obtained, an analysis of variance was carried out on the basis of the randomized complete block experimental design, for which Statistical Analysis System software (SAS, 2002) was used. Subsequently, Tukey's range test (α ≤ 0.05) was performed for the factors whose effect was significant. In those variables that showed significant interactions, tests were carried out to compare the means of the levels of one factor with each of the levels of the other factor.
Results and discussion
Analysis of variance
The analysis of variance (Table 1) indicates that for the variety (VAR) factor there were significant effects (α ≤ 0.01), which implies that the five variables evaluated behave differently for each variety. This same effect was observed in the time (TIM) factor on NF, FW, TSW and TNS, which indicates that the pollination time affects the number of fruits and seeds harvested, but not their size. As regards the family nested in variety [FAM(VAR)], significant effects (α ≤ 0.01) were found in TNS and W100S, which means that there is a difference between families of the same variety, which is reflected in a different amount and size of seed.
Table 1 Mean squares of the analysis of variance of five variables evaluated in two families of the Diamante and Manzano Tepetlixpa tomatillo varieties (2015 autumn-winter cycle).
| SV 1 | DF | NF | FW (g) | TSW (mg) | TNS | DF | W100S (mg) |
|---|---|---|---|---|---|---|---|
| BLO | 3 | 4.98 | 40752.9 | 1241434.6 | 319342.4 | 3 | 235.2 |
| VAR | 1 | 17.11** | 109076.5* | 8070851.3** | 6276480.8** | 1 | 13852.3** |
| FAM(VAR) | 2 | 0.16 | 35805.1 | 1487656.3 | 1095696.1* | 2 | 2802.4** |
| TIM | 4 | 13.96** | 326066.5** | 13490055.0** | 4996175.5** | 4 | 787.7 |
| VAR*TIM | 4 | 1.27 | 75718.7** | 1123876.3 | 528129.6* | 4 | 357.6 |
| Error | 65 | 1.83 | 20408.5 | 859192.3 | 246529.9 | 64 | 345.2 |
| Total | 79 | 78 | |||||
| CV | 17.11 | 31.77 | 37.67 | 34.99 | 10.44 |
1SV: source of variation; FAM(VAR): family nested in variety; VAR*TIM: variety x time interaction; CV: coefficient of variation; DF: degrees of freedom; NF: number of fruits; FW: fruit weight; TSW: total seed weight; TNS: total number of seeds; W100S: weight of 100 seeds.
*, **: significant with α ≤ 0.05 and α ≤ 0.01, respectively.
The variety x time interaction was significant (α ≤ 0.01) for the FW and TNS variables, which implies that there is a different optimal pollination time for each variety. Therefore, it is necessary to study each level of the time factor within each variety to identify the optimal pollination time that maximizes the amount of seeds harvested.
Comparison of means of the varieties
The results of the comparison of means between varieties (Table 2) show that Diamante had a higher NF, TSW and TNS, but lower FW and W100S, which indicates that this variety had fruits of less weight with more smaller seeds. For Manzano Tepetlixpa, the highest values (α ≤ 0.01) were obtained in FW and W100S, which shows that this variety has larger fruits than Diamond, but with fewer and larger seeds. In this regard, Peña-Lomelí et al. (2014) mention that the Diamante variety has large fruits (41 g∙fruit-1) and a yield of 787 g∙plant-1, while Manzano Tepetlixpa has a yield of 972 g∙plant-1 with medium-sized fruits. The difference in fruit size may be due to the fact that in this work selected Manzano Tepetlixpa families were evaluated.
Table 2 Comparison of means of the variables evaluated in two tomatillo varieties (2015 autumn-winter cycle).
| Variety | NF 1 | FW (g) | TSW (mg) | TNS | W100S (mg) |
|---|---|---|---|---|---|
| Diamante | 8.38 az | 412.8 b | 2778.0 a | 1699.0 a | 164.9 b |
| Manzano Tepetlixpa | 7.45 b | 486.6 a | 2142.8 b | 1138.8 b | 191.5 a |
| HSD | 0.60 | 63.8 | 414.0 | 221.7 | 8.4 |
1NF: number of fruits; FW: fruit weight; TSW: total seed weight; TNS: total number of seeds; W100S: weight of 100 seeds; HSD: honest significant difference.
zMeans with the same letter within a column do not differ statistically (Tukey, α ≤ 0.05).
Comparison of means of pollination time (days)
Pollination on the second day produced the highest (α ≤ 0.01) FW and TSW, while that on day three was higher (α ≤ 0.01) in NF and TNS (Table 3). However, the results obtained for these variables on both days do not differ statistically. This implies that manually pollinating on the second or third day, after the floral bud has been covered, yields the best results and can be used as a practice in breeding programs.
Table 3 Comparison of means of the variables evaluated on five different days in which manual pollination was carried out (2015 autumn-winter cycle).
| Time (day) | NF 1 | FW (g) | TSW (mg) | TNS | W100S (mg) |
|---|---|---|---|---|---|
| 1 | 8.19 az | 410.8 b | 1,740.6 bc | 946.5 c | 188.4 a |
| 2 | 8.50 a | 608.2 a | 3,345.0 a | 1,916.8 ab | 176.3 ab |
| 3 | 8.63 a | 548.9 ab | 3,311.3 a | 2,000.3 a | 169.4 b |
| 4 | 7.94 a | 443.1 b | 2,597.5 ab | 1,473.1 b | 180.6 ab |
| 5 | 6.31 b | 237. 6 c | 1,307.5 c | 757.8 c | 175.3 ab |
| HSD | 1.34 | 141.7 | 919.5 | 492.6 | 18.6 |
1NF: number of fruits; FW: fruit weight; TSW: total seed weight; TNS: total number of seeds; W100S: weight of 100 seeds; HSD: honest significant difference.
zMeans with the same letter within each column do not differ statistically (Tukey, α ≤ 0.05).
In the pollinations performed from the fourth day, the variables FW, TSW and TNS showed a significant decrease compared to the second and third day. The same happened on the first day. The above indicates that pollinating closed floral buds, as recommended by Pérez-Grajales et al. (1998), is not the best technique to make crosses in tomatillo, as these should be covered one day and pollinated the next.
In the control (non-pollinated covered buds), only parthenocarpic fruits developed, so it was possible to quantify the NF and FW (data not shown), but no data was obtained for TSW, TNS and W100S. This confirms that in the populations used there were no self-compatible mutants that alter the results, such as those reported by Mulato-Brito et al. (2007).
Analysis of families in each variety
This factor had significant effects for the TNS and W100S variables. In Figure 1 it can be seen that family one of the Diamante variety obtained significantly higher TNS than family two, whereas the Manzano Tepetlixpa families did not exhibit a significant difference in this same variable.
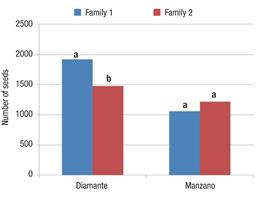
Figure 1 Total number of seeds (TNS) produced by fruits of the two families of the Diamante and Manzano Tepetlixpa varieties manually pollinated on five different days. Within each variety, bars with the same letter do not differ statistically (Tukey, α ≤ 0.05).
The W100S obtained from family one was significantly higher than that of family two, both of the Diamante variety, a difference that was not observed in Manzano Tepetlixpa (Figure 2).

Variety x time interaction
This interaction was found in FW and TNS and indicates that the behavior of the cultivars varies with the day on which the manual pollination was carried out.
When pollinating the third day, Diamante managed to reach a higher FW (Figure 3), but it did not statistically differ from days one, two and four, which indicates that with pollination during the first four days fruits of similar weight are obtained in the Diamante variety. On the other hand, Manzano Tepetlixpa reached its highest FW on the second day, although it did not statistically exceed the third day.
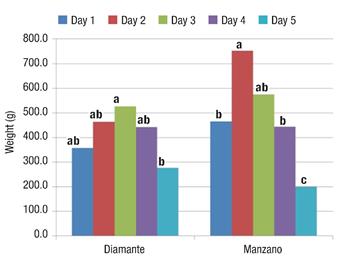
Figure 3 Fruit weight (FW) of the Diamante and Manzano Tepetlixpa tomatillo varieties manually pollinated on five different days. Within each variety, bars with the same letter do not differ statistically (Tukey, α ≤ 0.05)
In Diamante the highest TNS was obtained by pollinating the flower on the third day, although this value did not differ statistically from those obtained on day two and four. For the Manzano Tepetlixpa variety, on the second day of pollination, a higher TNS was obtained, although it was similar to that obtained on day three (Figure 4).
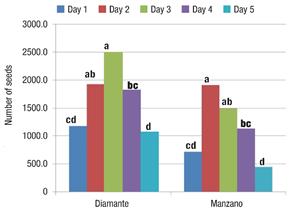
Figure 4 Total number of seeds (TNS) of the Diamante and Manzano Tepetlixpa tomatillo varieties manually pollinated on five different days. Within each variety, bars with the same letter do not differ statistically (Tukey, α ≤ 0.05).
When analyzing the variables FW and TNS together, it can be seen that both varieties obtained better results when pollinating on days two and three, since it is when better fruit development and a greater amount of seed is achieved. This indicates that the greatest amount of seed is obtained by pollinating on day three for Diamante and on day two for Manzano Tepetlixpa. The above differs from the findings reported by Pérez-Grajales et al. (1998), who point out that pollination should be done in closed flower buds, possibly because they used different varieties.
In summary, the findings of this research have helped to clarify the appropriate time interval to carry out manual pollination of the tomatillo. When pollinating the flowers two and three days after having covered the buds, fruit set was not significantly increased with respect to day one recommended by Pérez-Grajales et al. (1998), but seed production is doubled. This represents a substantive improvement in the efficiency of manual crosses, either within a breeding program or in the regeneration of accessions safeguarded in germplasm banks.
Conclusions
The Diamante variety produced a high number of seeds per fruit, while Manzano Tepetlixpa produced fruits with few seeds but with greater weight.
The suggested period for manual pollination is between two and three days after having covered the floral buds. For Manzano Tepetlixpa, better results were obtained on day two, and for Diamante, day three.
In Diamante differences were found between families for number of seeds and weight of 100 seeds; this implies that there is diversity among the families of this variety, unlike those of Manzano Tepetlixpa that did not differ in relation to the same variables.

