











 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroduction
Phytophthora infestans (Mont.) de Bary is the causal agent of late blight in potato (Solanum tuberosum L.), tomato (Solanum lycopersicum L.) and other solanaceae. The oomycete infects the entire plant and disperses its sporangia rapidly by the wind if the conditions are right. If the crop is not protected with fungicides with different modes of action, resistance to some products could occur and the potato crops are destroyed in a matter of weeks (Abad & Ochoa, 1995; Díaz-de la Cruz, Lozoya-Saldaña, Sahagun-Castellanos, & Peña-Lomelí, 2014). Nevertheless, the best management measure has still been the use of fungicides, but the populations of P. infestans have developed resistance to these products, favored by the continuous application of a single fungicide (Damicone, 2004).
Considering the above, metalaxyl is such an important product that it is still in use. It acts on specific sites of the pathogen, preventing the biosynthesis of proteins through interference in the synthesis of ribosomal RNA, which can be overcome by naturally resistant variants of the pathogen. This fact was reported in 1981 in Europe, and in 1993 in the U.S., Canada, Mexico and Ecuador (Davidse, Looijen, Turkesnteen, & van der Wal,1981; Drenth, Goodwin, Fry, & Davidse, 1993; Forbes et al., 1997; Matuszak, Fernández, Gu, Villarreal, & Fry, 1994; Marquinez, 1995; Power, Hamlen, & Morehart, 1995), to the point that this resistance is now considered an additional variable for characterizing P. infestans (Doster, Milgroom, & Fry, 1990; Dowley & O’Sullivan, 1985; Kato, Mizubuti, Goodwin, & Fry, 1997; Matuszak et al., 1994; Runno & Koppel, 2006).
Recently, other systemic substances have appeared on the market for the control of late blight, such as cyazofamid, which interferes with the mitochondrial respiration chain, and mandipropamid, which inhibits the synthesis of cellulose of the cell wall of the oomycete, acts on the protein PiCesA3 cellulose synthase, and is involved in the mycelium and haustoria formation during the initial incubation phase of the disease (Blum et al., 2010; Cohen et al., 2007). In addition, mandipropamid modifies the biosynthesis of the phospholipids and causes alterations in the F-actins (Griffiths, Dancer, O’ Neill, & Harwood, 2003).
Another product, fosetyl-Al, acts on the plant by stimulating its defense mechanisms, increasing its levels of peroxidases, phytoalexins, polysaccharides and the activity of the phenylalanine ammonia lyase (Brent & Hollomon, 2007; Gisi & Cohen, 1996; Robledo-Esqueda, Lozoya-Saldaña, & Colinas-León, 2012; Serrano-Cervantes, Lozoya-Saldaña, Colinas y León, & Leyva-Mir, 2016). The mode of action of this product is based on its influence on enzymatic phosphorylation and on the metabolism of the phosphorus, affecting the synthesis of different compounds that contain this element, essential for the growth of Phytophthora, such as nicotinamide adenine dinucleotide (NAD), adenosine triphosphate (ATP) and nucleotides (Gómez & Reis, 2011).
With respect to the pathogen, the heterothallic character of P. infestans confers evolutionary advantages, given that it increases the genetic variability and the exchange of virulence factors, thus on the population scale these recombination events generate more aggressive genotypes, adapting to various ecological niches and to the selection pressure by the fungicides (Adler et al., 2004; Erwin & Ribeiro, 1996; Goodwin, Sujkowski, & Fry, 1995; Knapova, Tenzer, Gessler, & Gisi, 2001). In Mexico, pathogen variability and resistance to metalaxyl have been reported in different agricultural regions (Garay-Serrano et al., 2007; Grünwald et al., 2006; Matuszak et al., 1994), particularly in Toluca Valley, state of Mexico, the center of origin of P. infestans (Grünwald et al., 2006; Grünwald & Flier, 2005).
With respect to the susceptibility of the fungicides, a new database was generated for a sexual population of the pathogen, with a wide range of sensitivity to metalaxyl, while for azoxistrobin, a chemical compound of the estrobilurine family, resistance was present (Evenhuis, Schepers, Bus, & Stegeman, 1996; Grünwald et al., 2001, 2006). These studies do not contemplate the area of Chapingo, Mexico, which is the second center of genetic diversification of the oomycete after Toluca (Goodwin, 1996), and it has the ideal climate for the development of the disease (Díaz de la Cruz et al., 2014), with wild populations of the oomycete (Alarcón-Rodríguez, Lozoya-Saldaña, Valadez-Moctezuma, García-Mateos, & Colinas-León, 2013).
Therefore, the objectives of the present study were to quantify the presence of late blight in the field in two growing cycles, detect the presence of variants of the oomycete resistant to agrochemicals, and compare the different levels of resistance in isolates from one cycle to another (in vivo first year vs in vivo second year) and in vivo with respect to in vitro in Chapingo, Mexico.
Materials and methods
Collection of isolates of P. infestans and infection readings
In the summer of 2011 and 2012, plots of potato cv. Fianna, susceptible to late blight, were established at the Autonomous University of Chapingo Experiment Station, located 25 km east of the Mexico City International Airport, at 2,250 masl with a C(Wo)(W)b(i)g climate, an average temperature of 15 °C and a reliable rainy season in the summer (645 mm). The experiment was conducting using a randomized block design with four rows of 4 m in length, with four replicates, under rainfed conditions and exposed to natural infection by the oomycete.
The treatments consisted of five weekly foliar sprayings of half dosages, according to the label, of metaxyl, fosetyl-Al, mandipropamid, propamocarb, cyazofamid, or water (absolute control) from 45 days after planting on. In order to get the disease progress curve, during the same period foliar infection readings were taken from ten individuals in the central rows per treatment, according to the Henfling scale (1987). The numbers of readings of the infection had a week of posterior offset with respect to the application of the products. Thus, the first reading corresponded to the second week of spraying, and the fifth reading was made a week after the fifth (and final) application of the products, that is, six weeks after the first spraying.
At the end of each cycle the comparison of means was made of the final reading, where the central rows were the useful plot. The area under the disease progress curve (AUDPC) was calculated according to Campbell and Madden (1990) and Shaner and Finney (1977), using the formula:
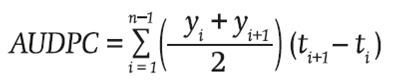
where the final value of AUDPC is the sum of the areas by lapses, which result from the multiplication of the average reading of two consecutive dates (y value) by the lapse (days, t value) between readings.
During the crop cycles samples of leaves and stems with simple lesions of blight were collected from each one of the treatments. They were subjected to moist chambers at room temperature (22-25 ºC) with white light for 24 h to induce the formation of sporangia, and disinfested potato slices were added over the lesion. After 2 d, mycelial growth which permeated the slices was observed. Mycelial tips were removed using a dissection needle and then sown in rye agar medium (Alarcón-Rodríguez et al., 2013) with oxytetracycline, and incubated at a constant temperature (17 °C).
Identification of mating types
To determine mating type (A1, A2), “unknown” isolates were plated at one end of a Petri dish with rye agar medium. At the other end of the dish a reference isolate of a known type was sown (A1 or A2, kindly provided by Dr. Sylvia P. Fernández Pavía, Universidad Michoacana de San Nicolás de Hidalgo) and maintained at a temperature of 17 ºC in darkness. In one week the colonial growths were assessed for developing sexual reproduction structures (oogonia, antheridia and oospores). The presence of oospores was interpreted as if the unknown isolate were from an opposite compatibility group to that of the confrontation. The presence of oospores when confronting the isolate with both types of known compatibility, or without confrontation, was interpreted as homothallic (H).
Resistance to fungicides in vitro
The evaluation of the resistance/susceptibility in vitro of the isolates to the fungicides applied in the field was carried out according to the methodology of Shattock (1988) and Deahl, DeMuth, Sinden, and Rivera-Peña (1995b). Mycelial fragments of the oomycete, isolated from the two crop cycles, were plated in selective rye-agar media with the fungicides added (0.1 and 10 mg∙L-1). The growth in vitro of each isolate was measured after an incubation period of 10 days at 17 ºC, and the result was quantified with the equation:

where PC = percentage of growth; DMCM = mean colony diameter (mm), 5 = diameter of the cylinder with mycelia originally sown at the center of the Petri dish (DM, diameter; C, cylinder; M, mycelia); and DMCA = diameter of the growth of the control without fungicides (DM, diameter; C, cylinder of growth, A, agar fungicide-free). For the classification, the criteria employed by Shattock (1988) and Deahl et al. (1995b) were adapted. They qualify the isolates with percentage of growth equal to or higher than 60 % as resistant to metalaxyl, intermediate from 10 % to 60 % growth, and as susceptible those with a percentage of radial growth less than 10 %. The comparison by categories was made in a completely randomized design (Statistical Analysis System [SAS Institute], 2004).
Results and discussion
Isolates and progress of the disease curve
P. infestans was highly sensitive to the fungicides during the first cycle, and all the isolates obtained were of the A1 mating type. In the second year all of the isolates were homothallic (Table 1). The A1 mating type is the most abundant in nature, and is the type that was dispersed throughout the world in the mid XIX century, but it is genetically limited, so it tends to be clonally conserved without much variation (Judelson, 1996).
Table 1 Number of isolates and mating type per year.
| Active ingredient | No. and MT z (2011) | No. and MT (2012) |
|---|---|---|
| Metalaxil | 3 A1 | 13 H |
| Fosetyl-Al | 3 A1 | 12 H |
| Mandipropamid | 5 A1 | 4 H |
| Ciazofamid | 7 A1 | 0 |
| Control | 5 A1 | 28 H |
| Total | 23 A1 | 57 H |
zMT: mating type (A1 and H [homothallic]).
Considering the absence of selection pressure in the zone prior to the assays due to the absence of commercial potato crops, it was expected that the population of P. infestans would be wild, predominating the above mentioned mating type and little infection in the treatments with the agrochemicals; thus, few were obtained in 2011. For the following year, all were homothallic, including the control, as evidence of the presence of these types that would bloom as a consequence of the situation of the previous year, given that both assays were established in the same plots in the two consecutive years.
Given that the oomycete population in the Chapingo area is quite wild because of the absence of a selection pressure (fungicides) in its natural environment, in the first year most of the A1 mating type (highly sensitive to fungicides) was eliminated, with no detection of homothallic populations that far. As a consequence, in the second year all of the pathogens present in the area were homothallic. This character had already been reported in Chapingo (Alarcón-Rodríguez et al., 2013) and in the Toluca Valley (López-Orona et al., 2013). The clonal nature of the isolates from Chapingo (all A1 the first year and all homothallic the second year) was later confirmed by microsatellite genotyping (Shakya, Larsen, Cuenca-Condoy, Lozoya-Saldaña, & Grünwald, 2016).
Percentage of infection
At the end of the two growing cycles, 100 % infection was detected in the unprotected controls. In 2011, the treatments that showed the highest protection were mandipropamid and metalaxyl, with 22 and 39 % infection, respectively, which was directly reflected in the value of the AUDPC (44 and 99, respectively, Figure 1, Table 2). In 2012 the treatment with the highest protection was again mandipropamid, with nearly 21 % final infection (AUDPC of 24), whereas metalaxyl permitted 87 % final foliar infection and 176 of AUDCP, and with the exception of mandipropamid, the rest of the fungicides allowed higher infection the second year with respect to the first (Figure 1, Table 2). This could be due to the selection pressure of the products on the pathogen during the previous cycle. Furthermore, in this second year the entire population of P. infestans was homothallic, which suggests that the pathogen could show higher genetic variability in 2012 than the previous year.

Figure 1 Progress of foliar infection:  metalaxil,
metalaxil,  fosetyl-Al,
fosetyl-Al,  mandipropamid,
mandipropamid,  ciazofamid,
ciazofamid,  control. zLines with the same letter do not differ statistically (Tuckey, P ≤ 0.05).
control. zLines with the same letter do not differ statistically (Tuckey, P ≤ 0.05).
Table 2 Final reading of foliar infection and area under the disease progress curve (AUDPC).
| Treatment | Last reading 2011 | AUDPC 2011 | Last reading 2012 | AUDPC 2012 |
|---|---|---|---|---|
| Metalaxil | 39.0 cz | 99 | 87.0 bc | 176 |
| Fosetyl-Al | 83.2 b | 185 | 98.2 ab | 226 |
| Mandipropamid | 22.0 d | 44 | 20.9 d | 24 |
| Ciazofamid | 99.5 a | 231 | 84.2 c | 219 |
| Control | 100.0 a | 265 | 100.0 a | 296 |
zMeans with the same letter within each column do not differ statistically (Tukey, P ≤ 0.05).
In the in vitro evaluations, the isolates grew over rye agar with 0.1 and 10 mg∙L-1 of the active ingredients. These doses were defined as a function of reports by Deahl, DeMuth, and Fry (1995a), Locher and Lorenz (1991), Marquinez (1995), Rekanović et al. (2011), Shattock (1988) and Sozzi, Schwinn, and Gisi (1992) who reported colonies with 2 to 100 % growth with respect to the controls.
Regarding the isolates exposed to cyazofamid in the field and in vitro, all of the isolates were sensitive in both crop cycles (Figure 2). The mode of action of this fungicide is fundamental for not developing resistance in the pathogen, given that it is related to the inhibition of the mitochondrial respiratory chain, in the reduction of ubiquinone (Qi) and the oxidation of ubiquinol (Qo) in cytochrome b (Temperli, Roos, & Itohl, 1990), with reduction of ATP. Cyazofamid limits the mobility of the zoospores, as well as the germination of sporangia and the formation of oospores (Ziogas, Markoglow, Theodosiou, Anagnostou, & Boutopoulou, 2006).

General discussion
The geographical area of Chapingo has ideal climatic conditions to grow potatoes, as well as for the development of the late blight disease in the summer. Nevertheless, potatoes are not extensively grown in the area. Thus, we would expect to deal with wild P. infestans populations in our assays, highly sensitive to agrochemicals, for there is no selection pressure of the pathogen by fungicides. However, the results came out otherwise. The best treatment in the field was mandipropamid, with consistency in control of the pathogen in both years, but the studies in vitro did not show the same results, given that although all of the populations were sensitive to the product in vitro the first year, populations bloomed with intermediate and high resistance to this product in the laboratory the second year.
The rest of the products showed inconsistencies in the advance curves of the disease in the field from one year to the next. Metalaxyl, for example, permitted an infection of 39 % in one year and 87 % in another. Furthermore, there was almost total infection in the treatment with cyazofamid the first year and in that of fosetyl-Al the second. This was also reflected in the in vitro assays, where the populations exposed to cyazofamid were consistently the most susceptible, without showing resistance, which would be just the opposite from what would be expected when in the field this product did not control the disease.
On the other hand, fosetyl-Al failed to control the blight in the field in both years, but there were intermediate and resistant populations of the oomycete when exposed to the product in vitro. The inconsistencies of the oomycete regarding fungicide resistance/sensitivity under in vivo/in vitro conditions, as well as from one year to another in the field, do not support behavior predictability. These results expose the great genetic plasticity of the oomycete in relation to its sensitivity to fungicides, and questions the in vitro assays of the products as potential fungicides along the development process, because in vitro pathogen control does not guarantee the same results in vivo.
Conclusions
There was high severity by Phytophthora infestans in both years in the field (in vivo), where three of the treatments presented curves and values of the disease that were different from one year to another. The half doses of the agrochemicals partially protected foliage, delaying the epidemic but allowing the infection and blooming of strains of P. infestans that were resistant to the products. The in vitro resistance of the oomycete to the fungicides was partial, depending on the doses to which it was exposed, without coincidence of resistance/susceptibility when compared with its behavior in vivo (field). The inconsistency of the behavior of P. infestans to these two conditions (in vivo, in vitro) does not allow the anticipation of responses of the oomycete under diverse situations of selection pressure from agrochemicals, which demonstrates its high genetic plasticity.

