










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista Chapingo. Serie horticultura
versão On-line ISSN 2007-4034versão impressa ISSN 1027-152X
Rev. Chapingo Ser.Hortic vol.18 no.3 Chapingo Set./Dez. 2012
https://doi.org/10.5154/r.rchsh.2012.06.31
¿Variedades sintéticas derivadas de cruzas simples o de cruzas dobles?
Synthetic varieties derived from single or double crosses?
Jaime Sahagún–Castellanos* y Clemente Villanueva–Verduzco
Departamento de Fitotecnia, Universidad Autónoma Chapingo. km 38.5 Carretera México–Texcoco, C. P. 56230, Chapingo Estado de México (*Autor para correspondencia) Correo–e: jsahagunc@yahoo.com.mx.
Recibido: 15 de junio, 2012.
Aceptado: 13 de julio, 2012.
Resumen
Las variedades sintéticas (VSs) formadas con cruzas simples (SinCS) o dobles (SinCD) permiten evitar el alto costo de semilla de variedades híbridas como las de maíz (Zea mays L.) y cebolla (Allium cepa L.). Además, con estas VSs se podrían predecir más económicamente las medias genotípicas (MGs) de las VSs que se generarían con las líneas progenitoras de tales híbridos (SinL). Sin embargo, se sabe que el SinL, SinCS y el SinCD generados con las mismas líneas pueden tener coeficientes de endogamia (CEs) diferentes, pero se desconoce por qué. Este desconocimiento limita al fitomejorador para hacer que el SinCS y SinCD representen mejor al SinL y para mejorar la estimación de su MG. El objetivo de este trabajo fue determinar qué hace que el SinL, SinCS y SinCD puedan diferir. Con L líneas no emparentadas cuyo CE es FL se formaron L/2 cruzas simples (CSs) y L/4 cruzas dobles (CDs); cada línea y cada híbrido se representó por m plantas, y por apareamiento aleatorio de los progenitores correspondientes se formó el SinL, SinCS y SinCD. Se encontró que para valores fijos de m y de FL, en general, los arreglos genotípicos de las tres VSs difieren porque con las mismas cuatro líneas progenitoras de una CD se aporta a la VS un número más grande de genes no idénticos por descendencia en la medida en que menos líneas integren un progenitor [línea (1), cruza simple (2), cruza doble (4)]. Así, el SinL y el SinCD tienen el menor y mayor CE, respectivamente, y se espera que las MGs tengan el orden de magnitud opuesto. Además, los arreglos genotípicos de las VSs difieren más a medida que FL y m sean más pequeños. Por el contrario, cuando las líneas son puras, los del SinL y el SinCS son iguales y los coeficientes de endogamia deben ser iguales también. Para que el SinCD también sea igual al SinL se requiere, además de líneas puras, un valor grande de m. Sin embargo, aun con FL < 1, en la medida en que m es más grande el SinCS y el SinCD también tienden a SinL, aunque más lentamente, en particular en el SinCD.
Palabras clave adicionales: Allium cepa L., Zea mays L., endogamia, variedades sintéticas, híbridos, arreglo genotípico.
Abstract
The use of synthetic varieties (SVs) derived from single (SynSC) or double (SynDC) crosses is a way of avoiding the high seed cost of hybrid maize (Zea mays L) and onion (Allium cepa L.) varieties, among others. In addition, these SVs could be used to more economically predict the genotypic means (GMs) of the SVs derived from the hybrid parental lines (SynL). It is known, however, that the inbreeding coefficients (ICs) of the SynL, SynSC, and SynDC derived from the same lines can differ, but the reason why is unknown. This lack of knowledge makes it difficult for the breeder to improve the representation of the SynL through the SynDC and SynSC and thereby to improve the estimation of the SynL GM. The objective of this study was to determine the reason why the SynL, SynSC, and SynDC can differ. From unrelated L lines whose IC is FL, L/2 single (SCs) and L/4 double crosses (DCs) were derived; and each line, SC, and Dc were represented by m individuals. By randomly mating each set of parents, the SynL, SynSC, and SynCD were formed. It was found that the genotypic arrays of the three SVs differ because with the same 4 initial lines the number of non–identical by descent genes contributed to the genotypic array is larger as fewer lines are involved in a parent [lines (1), single crosses (2), and double crosses (4)]. Thus, the ICs of SynL and SynDC are the smallest and largest, respectively; and it is expected that the GMs show the opposite ordering. In addition, as m and FL are smaller the differences among the genotypic arrays of the SVs are larger. On the contrary, when the L lines are pure, the genotypic arrays of SynL and SynSC are equal, and their IC must be equal as well. To be equal to SinL, SynDC requires, in addition, a large m. However, even with FL < 1, as m is larger the genotypic arrays of SynSC and SynDC tend to the genotypic array of SynL, although slowly, particularly for SynDC.
Additional keywords: Allium cepa L., Zea mays L., inbreeding, synthetic varieties, hybrids, genotypic array.
INTRODUCCIÓN
Recientemente, las variedades sintéticas (VSs) de cultivos como la cebolla (Allium cepa L.) y el maíz (Zea mays L.), cuyos progenitores son cruzas simples (Sahagún y Villanueva, 1997) o cruzas dobles (Márquez–Sánchez, 2008), han sido tema de estudio. Entre los argumentos para justificar estos estudios se encuentran los siguientes: i) que las VSs resultantes deben ser poblaciones muy similares a las que se formarían con las líneas que generan los híbridos progenitores del sintético resultante (Sahagún, 2011); ii) que algunos agricultores en México forman variedades sintéticas para evitar el alto costo de la semilla de las variedades híbridas, ya que la semilla cosechada de una VS regenera la misma variedad sin cambio en la media genotípica, lo que sí sucede en los híbridos (Villanueva et al., 1994), y iii) que la reducción del número de progenitores de la VS que se genera con dos, tres o cuatro líneas para formar un solo híbrido reduce los costos y trabajo necesarios para predecir el rendimiento de las VSs posibles de ser formadas con las líneas progenitoras de tales híbridos. Cuando el número de progenitores potenciales se reduce en una unidad, el número de poblaciones que se tiene que formar y evaluar experimentalmente para efecto de predicción según el método de Wright (1981), disminuye en un 50 %, aproximadamente.
Con el apareamiento aleatorio de L/2 cruzas simples se forma una variedad sintética que tiene el mismo coeficiente de endogamia que el de la VS que se formaría con el apareamiento aleatorio de las L líneas progenitoras de esos L/2 híbridos (Márquez–Sánchez, 2007). Sin embargo, la VS formada con el apareamiento aleatorio de L/4 cruzas dobles tiene un coeficiente de endogamia que difiere del de la VS formada con L/2 cruzas simples construidas con todas las mismas L líneas (Sahagún, 2011). Para generar información que contribuya a determinar el origen y la magnitud de tal diferencia, debe ser útil determinar qué es la VS derivada con L líneas en relación a las VSs cuyos progenitores son L/2 cruzas simples o L/4 dobles formadas, en ambos casos, con la participación de todas las L líneas. La información que se obtenga debe ser útil para evaluar el potencial de las VSs construidas con híbridos y para hacer predicciones más precisas y económicas, y debe contribuir al desarrollo de mejores variedades sintéticas. El objetivo de este estudio es determinar por qué las VSs formadas con las líneas con que se formaría un conjunto de híbridos de cruza simple o doble puede diferir del de las VSs que se formarían con tales híbridos.
MÉTODOS Y MARCO TEÓRICO
En este estudio se emplearon la notación y el enfoque utilizados por Sahagún (2011) para el modelo de un locus de una especie diploide que se reproduce por apareamiento aleatorio. Brevemente, en este enfoque se utilizaron dos conceptos: si en una población el gen Ai y el genotipo Ai Aj tienen frecuencias Pi y Pij, los arreglos gamético (AGA) y genotípico (AGE) de tal población se definen como:

Si en particular la reproducción de una población es por apareamiento aleatorio, como en el caso de las VSs, el arreglo genotípico de su progenie (AGEP) es igual al cuadrado del AGA de tal población. Por ejemplo, para una población cuyo AGA es el de la Ecuación 1, el apareamiento aleatorio debe producir la progenie cuyo AGEP es:

Además, si los arreglos gaméticos de las poblaciones P1 y P2 (AGAP1 y AGAP2) son ΣiPiAi y ΣjqjAj, su cruza produce una progenie cuyo arreglo genotípico [AGE (P1 x P2)] es:

Para la derivación de las variedades sintéticas (VSs) se partió de L líneas, formadas comúnmente por autofecundación, no emparentadas y con un coeficiente de endogamia FL. Con la participación de todas las L líneas en cada tipo de híbridos, se consideró la formación de L/2 cruzas simples (CSs) y de L/4 cruzas dobles (CDs), y cada línea y cada híbrido se representó por m plantas. Luego, por el apareamiento aleatorio de las líneas, de las CSs y de las CDs por separado, se formaron los tres tipos de sintéticos. Las diferencias entre las VSs se expresaron en términos de sus arreglos genotípicos y sus consecuencias en términos de sus coeficientes de endogamia.
DERIVACIÓN DE RESULTADOS
En la VS formada con las L líneas, si Aip1 Aip2 es el genotipo del p–ésimo individuo (p = 1,2,…,m) de la línea i (i = 1,2,…,L), la frecuencia del gameto Aipk (k=1,2) debe ser (2mL)–1, y el arreglo gamético de los mL individuos que representan a los L progenitores (AGAPL) de la VS es:

El arreglo genotípico de la VS que resulta del apareamiento aleatorio de los mL individuos que representan a los L progenitores (AGESL) es, por lo tanto,

A continuación se investigará con detalle en qué medida la variedad sintética (VS) cuyos progenitores son las L líneas, puede ser visualizada como una población que resulta del apareamiento aleatorio de un grupo de híbridos (L/2 cruzas simples o L/4 cruzas dobles).
Cruzas simples
Supóngase que L, el número de líneas iniciales, es un múltiplo de 4 (aunque los resultados para el sintético de cruzas simples son igualmente válidos para números pares de líneas). Supóngase ahora que en forma aleatoria con muestreo iterativo sin reemplazo de tamaño 2 se forman L/2 subconjuntos de 2 líneas cada uno. Supóngase, también, sin pérdida de generalidad, que el s–ésimo par de líneas es el conjunto {Ls–1, Ls}, s = 2,4,…,L; y que el genotipo del p–ésimo individuo (p = 1,2,…,m) de la línea s–1 es Ajp1Ajp2 (i = s–1); similarmente, el genotipo del q–ésimo individuo (q = 1,2,…,m) de la línea s es Ajq1Ajq2 (j = s). Así, de acuerdo con la Ecuación 5, el arreglo genotípico de la población producida por el apareamiento aleatorio de los 2m individuos que representan a las líneas Ls–1 y Ls [(AGES)s] debe ser:

(AGES)s (Ecuación 6) produce a su vez el arreglo gamético [(AGAS)s]:
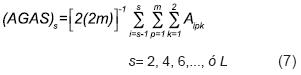
Por otra parte, los arreglos gaméticos de las líneas Ls–1 y Ls [(AGAL)s–1 y (AGAL)s] deben ser:

y

Según las Ecuaciones 8 y 9, el arreglo genotípico de la cruza simple Ls–1 X Ls, s = 2,4,…, ó L [(AGECS)s] es:

Evidentemente, (AGECS)s (Ecuación 10) produce el arreglo gamético [(AGACS)s]:

Como se puede observar, los arreglos gaméticos de una VS generada por el apareamiento aleatorio de los 2m individuos que representan dos líneas (Ecuación 7) y el de la progenie que produce la cruza entre ellas (Ecuación 11) son iguales, no obstante que provienen de poblaciones diferentes. Son diferentes en su arreglo genotípico, sí, pero sus frecuencias génicas son iguales. En primera instancia, esto significa que el apareamiento aleatorio entre los integrantes de cada una de las dos poblaciones por separado descritas en las Ecuaciones 6 y 10 produce un sintético cuyo arreglo genotípico es el mismo [(AGESCS)s]:

En segunda instancia, la mencionada igualdad de arreglos gaméticos (Ecuaciones 7 y 11) implica que la cruza entre dos poblaciones cuyos arreglos genotípicos son (AGES)s y (AGES)r (r ≠ s, Ecuación 6) debe producir la misma progenie que generaría la cruza entre las subpoblaciones cuyos arreglos genotípicos son (AGECS)s y (AGECS)r [Ecuación 10]. En particular, de la Ecuación 11, para r ≠ s:
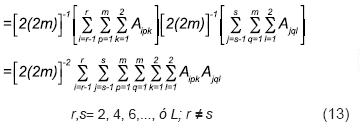
Con base en las Ecuaciones 12 y 13, se mostrará cómo el arreglo genotípico de la VS cuyos progenitores son L líneas (Ecuación 5) puede ser visualizado como el de la VS generada por el apareamiento aleatorio de L/2 cruzas simples formadas cada una con una pareja de líneas definida por muestreo aleatorio de tamaño 2 sin reemplazo a partir del conjunto inicial de L líneas. En efecto, con sólo argumentos algebraicos, de la Ecuación 5 se obtiene, en principio, que:

Además, cuando r = s en la Ecuación 14, se puede visualizar que con cada valor de s (s = 2,4,…, ó L) se definen los elementos del arreglo genotípico que produciría el apareamiento aleatorio de la progenie de la cruza simple Ls–1 X Ls, representable como [(AGACSs)]2 (Ecuación 11). Por otra parte, para cada caso en que r ≠ s, los genotipos de la Ecuación 14 que satisfacen esta condición pueden ser visualizados como los de la progenie que produciría la cruza entre las cruzas simples cuyos arreglos genotípicos son: (AGECS)r y (AGECS)s, r ≠ s; ó (AGESCS)r y (AGESCS)s, r ≠ s (Ecuación 13). En resumen, de la Ecuación 14 y de estas últimas dos consideraciones, resulta que:

De las Ecuaciones 12 y 13, la ecuación anterior para AGESL se puede escribir como
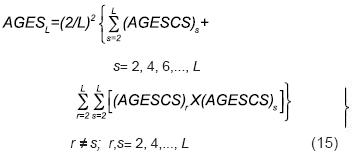
La Ecuación 15 y su origen implican que el AGESL es igual al arreglo genotípico de la VS cuyos progenitores son las L/2 cruzas simples hasta ahora consideradas (AGESXCS) ; es decir:

Por su origen, la Ecuación 16 debe ser válida para cualquier conjunto de L/2 pares de líneas que pueda producir el muestreo aleatorio sin reemplazo considerado para formar las L/2 cruzas simples. Sin embargo, debe tenerse en cuenta que las cruzas simples del AGESXCS son virtuales; es decir, aparentes, no se hicieron realmente. Además, en la Ecuación 14, el término 2mL de la expresión inicial se escribió posteriormente como 2(2m)L/2, en congruencia con la idea de representar el arreglo genotípico de un sintético formado con L/2 cruzas simples, cada una representada por 2m genotipos, que a su vez están formados por sendos pares de genes.
Cruzas dobles
En el segundo término de la Ecuación 15 se puede visualizar la presencia de los genotipos que produciría la cruza entre los híbridos simples r y s (r ≠ s). El arreglo genotípico de tal cruza doble [(AGECD) rs], de acuerdo con la Ecuación 13, es:

Para simplificar, sin pérdida de generalidad, supóngase que las líneas involucradas en la formación de la cruza doble d (d = 4,8,…,L) se representan por Ld–3 y Ld–2 si son las que forman la r–ésima cruza simple y por Ld–1 y Ld a las que forman la s–ésima cruza simple (r ≠ s; r, s = 2,4,6,…, ó
L). Con esta notación, el arreglo genotípico de la Ecuación 17 [(AGECDx)d] se puede expresar como:

El arreglo genotípico (AGECDx)d (Ecuación 18) produce el arreglo gamético [(AGACD)d] siguiente:

Por analogía (Ecuación 19), el arreglo gamético de la cruza doble e (e ≠ d) es de la forma:
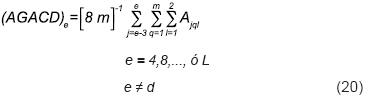
De acuerdo con las Ecuaciones 19 y 20, el arreglo genotípico de la cruza entre las cruzas dobles d y e (d ≠ e), [(AGECD)Xde] producido a su vez por el producto de sus arreglos gaméticos, es:

Evidentemente, si e = d,

Del arreglo genotípico de la VS formado por las L líneas iniciales (Ecuación 5), de la consideración de las Ecuaciones 21 y 22, y en congruen cia con el álgebra, se genera un resultado de importancia:

Evidentemente, la Ecuación 23, de acuerdo con las Ecuaciones 21 y 22, está formada por dos grupos de genotipos representados en sendos paréntesis rectangulares por: 1) el que contiene los genotipos que resultan del apareamiento aleatorio entre los individuos aquí visualizados como los representantes de la cruza doble d (Ecuación 22), y 2) el que contiene los genotipos que resultan del cruzamiento entre las cruzas dobles d y e (d≠e, Ecuación 21). Esto permite concluir que la VS formada por L líneas también puede ser visualizada como una VS formada por L/4 cruzas dobles, construidas con la participación de todas las L líneas, cada una representada a su vez por m plantas. Resumiendo, si el arreglo genotípico de esta VS se denota con AGESXCD, se ha encontrado (Ecuación 23) que

Además, de acuerdo con las Ecuaciones 16 y 24,

Como en el caso del AGESXCD , el resultado de la Ecuación 24 sería el mismo para cualquier conjunto de L/4 cruzas de este tipo que se pueda generar con la participación de todas las L líneas. Sin embargo, similar al caso del AGESXCs , las cruzas dobles del AGESXCD son virtuales; cada cruza doble es en realidad un conjunto de cuatro líneas.
DISCUSIÓN
Considérese la variedad sintética (VS) formada por L líneas no emparentadas de una o más autofecundaciones (SinL). Si cada línea se representa por m individuos y su coeficiente de endogamia es FL, el coeficiente de endogamia de tal VS (FSinL) es igual a (1 + FL)/(2L). Además, de acuerdo con Sahagún (2011) la VS cuyos progenitores son las m plantas de cada una de las L/2 cruzas simples convencionales (SinCS) formadas con la participación de todas las líneas con que se formó el SinL tiene un coeficiente de endogamia (FSinCS) igual a (1 + FL)/(2L) + (1 – FL)/(2mL). Finalmente, según Sahagún (2011), la VS cuyos L/4 progenitores son cruzas dobles convencionales, cada una representada por m plantas, tiene el coeficiente de endogamia (FSinCD):

Los resultados previos sobre el coeficiente de endo–gamia (CE) parecen contradecir a los generados en este estudio para los sintéticos cuyos arreglos genotípicos son AGESxCS y AGESxCD y que son iguales al del SinL (Ecuaciones 16 y 24). Por esto y por la consideración de que FSinL = (1 + FL)/(2L), debe ocurrir que respecto a sus CEs (FSx yFSxinCD ):

Sin embargo, FSinCS y FSinCD difieren entre si y de (1 + FL)/(2L). Diferencias importantes entre coeficientes de endogamia (CEs) en este estudio son:

y

Las Ecuaciones 27 y 28 implican que cuando FL es menor que 1, los coeficientes de endogamia de los sintéticos cuyos progenitores son cruzas simples convencionales (FSinCS) o cruzas dobles convencionales (FSinCD) son mayores que los de las VSs aquí denominadas SxinCS y SxinCD (FSxin y FSxin , respectivamente). Evidentemente, las diferencias en las Ecuaciones 27 y 28 se reducen a medida que FL se acerca a 1 y m es mayor. Con FL = 1 se reducen a 0 y 1/(mL), respectivamente; y cuando m es "grande", las diferencias (Ecuaciones 27 y 28) son despreciables (Cuadro 1), y SinL, SinCS y SinCD son prácticamente iguales. En estas condiciones, el fitomejorador puede válidamente optar por el uso de cruzas simples o dobles convencionales para formar las VSs que se producirían con las líneas progenitoras de tales híbridos (o viceversa). Con esta estrategia se lograría una reducción de trabajo y recursos para efectos de predicción, en caso de disponer de tales híbridos. Los problemas de esta estrategia con relación al uso de VSs convencionales son la reducción de la precisión en la estimación de la media de la VS (Sahagún y Villanueva, 1997; Sahagún, 2011) y de la flexibilidad en términos del número y de la elección de progenitores.
A continuación se hará un análisis del origen de las diferencias que muestran las Ecuaciones 27 y 28 en términos de arreglos genotípicos. De las reflexiones que en este contexto se hagan, comúnmente se encontrarán evidencias concretas en términos de coeficientes de endogamia en el Cuadro 1. Según estas diferencias, cuando FL < 1, y los valores de m son pequeños, estrictamente ni el arreglo genotípico de un sintético generado con L /2 cruzas simples (SinCS) ni el de uno generado con L/4 cruzas dobles (SinCD) pueden ser iguales al del sintético cuyos progenitores son las L líneas iniciales, que son las mismas con que se forman el SinCS y el SinCD. En el caso del SinCS, para que el apareamiento aleatorio de los m(L/2) individuos progenitores permita que todos los genes que no son idénticos por descendencia (NIPD) de las L líneas iniciales formen parte del arreglo genotípico del SinCS es necesario que FL = 1. Cuando FL < 1 y m es pequeño, esto puede no ser posible; por ejemplo, con m = 1, mientras que la planta que representa una cruza simple aporta dos genes NIPD, las dos plantas que representan a las líneas progenitoras de tal cruza simple pueden aportar conjuntamente dos, tres o hasta cuatro genes NIPD al SinL, dependiendo del valor de FL. Y esto hace que los arreglos genotípicos del SinL y del SinCS sean diferentes. Es claro, sólo si FL = 1, en ambos casos, el número de genes NIPD aportados al arreglo genotípico debe ser dos.
En el caso del SinCD, puede no ser posible que todos los genes NIPD de las cuatro líneas progenitoras de una cruza doble se muestren en su arreglo genotípico, cualquiera que sea el valor de FL y un valor pequeño de m. Con m = 1, por ejemplo, sólo dos de los cuatro o más genes NIPD que portan las cuatro líneas progenitoras pueden formar parte del arreglo genotípico del SinCD, puesto que tales líneas no tienen parentesco. Así, con valores pequeños de m el SinCS y el SinCD no pueden tener arreglos genotípicos iguales.
Evidentemente, las diferencias de los arreglos genotípicos de los sintéticos formados con cruzas simples y con cruzas dobles convencionales (SinCS y SinCD, respectivamente) respecto al del SinL (y consecuentemente a los del SxinCS y SxinCS ) cuando FL < 1 y m es pequeño, son consistentes con lassdiferencias entre los coeficientes de endogamia correspondientes. Los resultados del Cuadro 1 ejemplifican estas tendencias generales con números realistas de líneas para maíz (Kutka y Smith, 2007).
Finalmente, los resultados que se han generado en este estudio en términos de coeficientes de endogamia y de arreglos genotípicos permiten establecer que las variedades sintéticas cuyos progenitores son L líneas puras (FL = 1) son iguales a las variedades sintéticas construidas con L/2 cruzas simples formadas con la participación de todas las L líneas, independientemente del tamaño de m y de la forma de integración de las parejas de líneas que forman las L/2 cruzas simples. Cuando FL < 1, la diferencia entre los coeficientes de endogamia de los sintéticos hechos uno con cruzas dobles convencionales y con cruzas dobles virtuales, FSinCD –FSxinCD (Ecuación 27), depende de la magnitud de FL y de m; a medida que éstos sean más grandes, se reduce según la expresión (3 – FL)/(2mL). Por otra parte, el apareamiento aleatorio de las L/4 cruzas dobles que se puedan definir mediante muestreo sin reemplazo de tamaño 2 tomadas del conjunto de L/2 cruzas simples hasta agotar las L/2 cruzas simples genera un arreglo genotípico que, aun con líneas puras (FL = 1), difiere del SinL. En términos del CE, la diferencia es 1/(mL), que tiende a cero cuando m es más grande. Si FL < 1, la diferencia FSinCD – FSinL es mayor y la velocidad con que tiende a cero es más lenta que en el caso de las cruzas simples.
CONCLUSIONES
Se encontró que la variedad sintética (VS) producida por el apareamiento aleatorio de L líneas no emparentadas cuyo coeficiente de endogamia (CE) es FL, y son representadas por sendos conjuntos de m plantas, tiene un CE (FSinL) que nunca es mayor que el de la VS derivada del apareamiento aleatorio de L/2 cruzas simples (FSinCS) o L/4 cruzas dobles (FSinCD) generadas con las mismas L líneas. Sólo con líneas puras (FL = 1), FSinL = FSinCS. Cuando FL < 1, las diferencias FSinL – FSinCS y FSinL – FSinCD, que son (1 – FL)/(2mL) y (3 – FL)/(2mL), respectivamente, se vuelven marginales y los arreglos genotípicos de los tres tipos de VSs son prácticamente iguales cuando FL y mL son grandes. Las diferencias entre las VSs se deben a que cada conjunto de cuatro líneas iniciales aporta al arreglo genotípico de la VS un número más grande de genes no idénticos por descendencia en la medida en que menos líneas integren cada tipo de progenitor [líneas (1), cruzas simples (2) o cruzas dobles (4)]. Esto es lo que hace que las tres VSs puedan diferir.
LITERATURA CITADA
KUTKA F., J.; SMITH M.E. 2007. How many parents give the highest yield in predicted synthetic and composite populations of maize? Crop Science 47: 1907–1913. [ Links ]
MÁRQUEZ–SÁNCHEZ, F. 2007. Predicción de sintéticos de maíz hechos con cruzas simples. Agrociencia 41: 271–274. [ Links ]
MÁRQUEZ S., F. 2008. Endogamia y predicción de sintéticos de maíz de cruzas dobles. Revista Fitotecnia Mexicana 31: 1–4. [ Links ]
SAHAGÚN C., J. 1994. Sobre el cálculo de coeficientes de endogamia de variedades sintéticas. Agrociencia Serie Fitociencia 5(2): 67–78. [ Links ]
SAHAGÚN C., J.; VILLANUEVA V., C. 1997. Teoría de las variedades sintéticas formadas con híbridos de cruza simple. Revista Fitotecnia Mexicana 20: 69–79. [ Links ]
SAHAGÚN C., J. 2011. Inbreeding and yield of synthetic varieties derived from single and double cross hybrids. Maydica 56(3): 265–272. [ Links ]
VILLANUEVA V., C.; CASTILLO G., F.; MOLINA G., J. D. 1994. Aprovechamiento de cruzamientos dialélicos entre híbridos comerciales de maíz: Análisis de progenitores y cruzas. Revista Fitotecnia Mexicana 17: 175–185. [ Links ]
WRIGHT A., J. 1981. The quantitative genetics of diploid synthetic varieties. Proc. Fourth meeting of the section biometrics in plant breeding. Poitiers, France, Sept. 2–4. pp. 137–157. [ Links ]

