










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Chapingo. Serie horticultura
versión On-line ISSN 2007-4034versión impresa ISSN 1027-152X
Rev. Chapingo Ser.Hortic vol.17 no.2 Chapingo may./ago. 2011
Crecimiento de plántulas de estátice (Limonium sinuatum) y viola (Viola cornuta) en ambientes contrastantes
Growth of statice (Limonium sinuatum) and viola (Viola cornuta) seedlings in contrasting environments
Oscar Javier Ayala–Garay1*; José Alfredo Carrillo–Salazar1; Evelia Hernández–García1; Elizabeth Díaz–Martínez2; Manuel Livera–Muñoz1, Gustavo Almaguer–Vargas3
1 Recursos Genéticos y Productividad, Colegio de Postgraduados. km 36.5 Carretera México–Texcoco. Montecillo, Estado de México. C. P. 56230. MÉXICO. Tel. 01 (595) 952–0200 Ext. 1594. Correo–e: oayala@colpos.mx (*Autor para correspondencia).
2 Licenciatura en Agronomía. División de Ciencias Biológicas y de la Salud. Unidad Xochimilco, Universidad Autónoma Metropolitana. México D.F.
3 Departamento de Fitotecnia, Universidad Autónoma Chapingo. km 22.5 Carretera México–Texcoco. Chapingo, Estado de México. C.P. 56200. MÉXICO.
Recibido: 10 de octubre, 2010.
Aceptado: 23 de mayo, 2011.
Resumen
En producción rústica de ornamentales se requiere conocer el efecto medioambiental en la emergencia y crecimiento de plántulas para optimizar el proceso de producción. El objetivo de esta investigación fue estudiar el efecto de tres condiciones de producción: un túnel cubierto por polietileno, malla–sombra de 50 % de transmisión de radiación solar y a la intemperie, en el crecimiento de plántulas de estátice y viola, durante el invierno en el Valle de México. Se midieron la temperatura del aire, porcentaje de emergencia (E%), área foliar por planta (AF) y biomasa seca por planta (PS) a los 10,18, 24, 31 y 35 días después de siembra (dds) en estátice, y 21, 28, 35, 43 y 52 dds en viola; se calcularon las tasas absoluta de crecimiento (TAC), relativa de crecimiento (TRC) y de asimilación neta (TAN). En túnel de polietileno y malla–sombra se tuvieron valores de E % más altos: 95 y 93 % para estátice y 97 y 95 % para viola, respectivamente. En ambas especies existieron dos cinéticas sigmoidales de crecimiento en PS: la cinética de crecimiento en el túnel de polietileno fue la mayor (P<0.05) con temperatura media de 19.5 °C y alta oscilación térmica, pero sin heladas. En el túnel de polietileno, las máximas PS, AF y TAC (88 mg·plántula–1,15.1 cm2·plántula1 y 7.4 mg·d–1, respectivamente) se observaron a los 35 dds en estátice y a los 52 en viola (43.6 mg·plántula1, 7.6 cm2·plántula1 y 2.4 mg·d1, respectivamente) antes del trasplante. En el túnel de plástico no se presentaron temperaturas inferiores a 0 °C, lo que explica el mayor crecimiento de las plántulas.
Palabras clave: Emergencia de plántulas, temperatura del aire, tasa absoluta y relativa de crecimiento.
Abstract
In order to grow ornamental plants in non–controlled environments, the effect of temperature on seedling emergence and growth must be known in order to optimize plant production. The aim of this research was to compare the growth of statice and viola seedlings, during the winter in the Valley of Mexico, under three production conditions: a polyethylene–covered tunnel, shade netting that cuts light transmission by 50 %, and in the open. We measured air temperature (0C), emergence percentage (E%), leaf area (LA) per plant and dry weight biomass (DWB) per plant at 10, 18, 24, 31 and 35 days after planting (dap) in statice seedlings, and at 21, 28, 35, 43 and 52 dap in viola seedlings. In addition, the absolute growth rate (AGR), the relative growth rate (RGR) and the net assimilation rate (NAR) were calculated. Compared to the outdoor treatment (control), The polyethylene tunnel and shade netting produced higher E% values: 95 and 93 % for statice and 97 and 95 % for viola, respectively. Two different sigmoid curves of dry matter accumulation were observed in both species. The polythene tunnel produced higher DWB than the other conditions (P<0.05) with an average temperature of 19.5 0C and high thermal oscillation, but without frost. In the polyethylene tunnel, maximum DWB, LA and AGR (88 mg·seedling–1, 15.1 cm2seedling–1, and 7.4 mg·d–1, respectively) were observed at 35 dap in statice and 52 dap in viola (43.6 mg·seedling–1, 7.6 cm2 seedlingH and 2.4 mg·d–1, respectively) before transplanting. The polythene cover prevented temperatures below 0 °C, which explains the greater seedling growth in this environment.
Key words : Seedling emergence, air temperature, absolute and relative growth rate.
INTRODUCCIÓN
Las especies ornamentales registran altos niveles de producción por hectárea y representan una derrama económica significativa debido a la infraestructura, insumos y mano de obra que necesitan para su cultivo (Flores–Amaraz y Lagunes–Tejeda, 1998). Dos de estas especies cultivadas son el estátice (Limonium spp.), ofrecido en el mercado deshidratado en arreglos florales, con 80 ha cosechadas y un valor de la producción de 27 millones de pesos, y la viola o pensamiento (Viola spp.), con 180 mil macetas producidas con un valor de la producción de 1.1 millones de pesos en el 2008 (Anónimo, 2010). El estátice es originario del Mediterráneo (Larson, 1992), mientras que la viola es encontrada en forma silvestre en zonas montañosas de la frontera de España y Francia (Kelly et al., 2006), por lo que ambas son tolerantes a temperaturas bajas. Estátice requiere de 10 a 13 °C de temperatura media para florecer más rápido, y viola tolera bien el frío y se recomienda cultivarla con 13 a 18 °C de temperatura media (Anónimo, 2005), por lo que son opciones viables de cultivo en condiciones invernales en el Valle de México, donde se tienen temperaturas promedio de entre 12 y 18 °C (García, 1981).
Las especies ornamentales en México se cultivan bajo alguna de las siguientes modalidades: intemperie, invernadero o malla–sombra, condiciones en las que se producen cambios en temperatura del aire y planta, humedad relativa y nivel de radiación que afectan el rendimiento, calidad del producto y tiempo de producción. En la producción de plántulas, es deseable el cultivo bajo un ambiente microclimático que favorezca su producción rápida, con la obtención de plantas sanas y vigorosas, lo que es importante económicamente en un proceso de producción intensivo (Ayala–Sierra y Valdez–Aguilar, 2008). Para investigar el efecto ambiental en el crecimiento y desarrollo, se tiene la herramienta del análisis de crecimiento vegetal, que permite entender la naturaleza de la interacción planta–ambiente (Hunt et al., 2002; Mora–Aguilar et al., 2005). Esta técnica de análisis calcula índices de eficiencia de producción de biomasa, como son la tasa relativa de crecimiento (TRC), la tasa absoluta de crecimiento (TAC) y la tasa de asimilación neta (TAN), entre otras (van der Werf, 1996).
La producción de plántulas de ornato puede ser redituable (Moran, 2004), pero en México aún es incipiente; las variedades y recomendaciones técnicas de manejo provienen del extranjero. Sin embargo, las condiciones de producción son diferentes a las recomendadas en los catálogos de las empresas semilleras; aquí la infraestructura es diferente, y se desconocen las técnicas apropiadas para producir dichas plántulas (Ayala–Sierra y Valdez–Aguilar, 2008). El objetivo de esta investigación fue estudiar el efecto del ambiente de producción en túnel de plástico, malla–sombra o intemperie, en el crecimiento de plántulas de estátice y viola en el Valle de México durante el invierno.
MATERIALES Y MÉTODOS
El estudio se realizó en Texcoco, Estado de México (19° 29' N, 98° 51' O y 2,250 m de altitud). Se utilizó semilla de estátice (Limonium sinuatum) cv. Fortress y Viola cornuta cv. Sorbet de la compañía Ball Horticultural Company, Estados Unidos de América. Las semillas fueron sembradas el 6 de febrero de 2009, en charolas de 200 cavidades rellenas con turba (Premier PRO–MIX® PGX, Canadá); cada charola se dividió en tres secciones, para obtener tres repeticiones por especie. Se colocó una charola de cada especie en tres ambientes: microtúnel de polietileno blanco lechoso 30 % de sombreo (1.5 m de ancho x 3 m de largo), microtúnel de malla–sombra 50 % de sombreo (1.5 m de ancho x 3 m de largo) y a la intemperie (testigo).
En cada tratamiento se colocó un data logger (HOBO H8–032–08, Onset Computer Corporation, Estados Unidos de América) para registrar la temperatura del aire (°C) cada 30 min. La irradiancia fotosintética incidente (tratamiento a la intemperie o testigo) fue calculada como el 50 % de la radiación global (Brown, 2010) medida en la estación agrometeorológica del Colegio de Postgraduados, en Montecillo, Estado de México, a 200 m del sitio experimental. La irradiancia fotosintética en el microtúnel de polietileno y de malla–sombra fue de 30 y 50 % de la irradiancia fotosintética incidente en el testigo, respectivamente, debido a las especificaciones de transmisividad de los materiales usados.
Se determinó el porcentaje de emergencia (E %) de plántulas sobre el sustrato de germinación a los 18 y 28 días después de la siembra (dds) para estátice y viola, respectivamente.
Para evaluar el crecimiento, se hicieron cinco muestreos de cinco plantas seleccionadas al azar por repetición, a los 10, 18, 24, 31 y 35 dds para estátice, y a los 21, 28, 35, 43 y 52 dds para viola. Las plántulas se separaron en hojas y tallos, se secaron en una estufa con aire forzado (modelo 31480, Thelco, Estados Unidos de América) a 70 °C hasta alcanzar peso constante, y posteriormente se registró la biomasa seca total (PS) con una balanza Ohaus (Pine Brook, China). Antes de secar las plántulas, se estimó el área foliar (AF) por planta. Para esto, primero se digitalizaron las imágenes de las hojas por planta con un escáner (HP PSC 1210. HP Company, Estados Unidos de América), y se calculó la superficie relativa cubierta de píxeles (unidades que componen una imagen) verdes de cada imagen con un programa en Microsoft Visual Basic® ver. 6.0 basado en el método propuesto por Otha et al. (1980). Con base en una superficie de referencia conocida (1 cm2), se calculó el área foliar por planta (AF).
Con la biomasa seca promedio y el área foliar promedio por planta, se calcularon los siguientes índices de eficiencia fisiológica (Hunt, 1990): tasa absoluta promedio de crecimiento (TAC), tasa relativa promedio de crecimiento (TRC) y tasa promedio de asimilación neta (TAN), con las fórmulas siguientes:

donde PS2 y PS1: biomasa seca de la plántula al final y al inicio de un periodo determinado; t1 y t2: tiempo transcurrido entre dos muestreos; AF2 y AF1: área foliar por plántula al final y al inicio del mismo periodo y ln: logaritmo natural.
Se utilizó un diseño experimental completamente al azar con tres tratamientos y tres repeticiones de cinco plantas (unidad experimental), donde las comparaciones fueron entre tratamientos para la misma especie.
La captura de datos y las curvas de crecimiento se hicieron en una hoja de cálculo. El análisis de varianza de E %, PS y AF se hizo con el programa Statistical Analysis System versión 8.2e (SAS Institute, 2003). Para las variables que mostraron efecto de tratamiento se hizo la prueba de comparación de medias de Tukey (P<0.05).
RESULTADOS Y DISCUSIÓN
Variables climáticas en los ambientes de producción
Durante el periodo experimental, las temperaturas del aire promedio en los tratamientos de túnel de polietileno e intemperie resultaron similares (Cuadro 1), existiendo sólo 0.4 °C de diferencia entre ellos. En cambio, en la condición del túnel de malla–sombra, la temperatura promedio del aire fue 3.5 °C menor que los otros tratamientos. El intervalo diario de la temperatura del aire (diferencia entre las temperaturas máxima promedio y mínima promedio) fue menor en el túnel de malla–sombra (27.1 °C) que bajo el túnel de polietileno (35.6 °C) o a la intemperie (29.3 °C). La temperatura mínima promedio del aire (Tmin) más baja fue registrada a la intemperie (2.6 °C). Tanto en el tratamiento a la intemperie (testigo) como en el túnel con malla–sombra se registraron ocho y tres días con heladas, respectivamente. En el tratamiento de túnel con malla–sombra, las temperaturas por debajo de 0 °C se presentaron a los 18, 24 y 25 días después de la siembra (dds), mientras que a la intemperie se observaron durante todo el periodo experimental.
El promedio diurno de la irradiancia fotosintética incidente durante el periodo experimental en el túnel de malla–sombra fue de 7.8 ± 0.94 MJ m–2·d–1, en el de polietileno 5.6 ± 0.67 MJ m–2·d–1 y en el testigo de 11.2 ± 1.3 MJ m–2·d–1.
Emergencia de plántulas
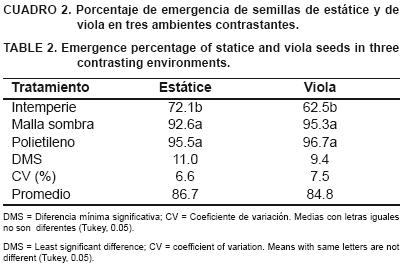
El porcentaje de emergencia de estátice (E %) en túnel plástico (95 %), fue estadísticamente igual a lo observado bajo malla–sombra (93 %), y ambos superaron al obtenido a la intemperie (72 %) (Cuadro 2). En viola se observaron resultados similares de E % en los ambientes evaluados (97, 95 y 63 %, respectivamente), siendo la germinación a la intemperie inferior estadísticamente a los otros dos tratamientos. Las condiciones climáticas prevalecientes (Cuadro 1), específicamente la presencia de ocho d de heladas en el tratamiento de crecimiento a la intemperie y una temperatura del aire mínima promedio más baja (2.6 °C), provocaron una germinación estadísticamente inferior en ambas especies.
Dinámica del crecimiento
La dinámica de acumulación de biomasa de la planta de estátice fue de tipo sigmoidal en los tres tratamientos estudiados (Figura 1A). Esta dinámica de crecimiento es típica de órganos e individuos vegetales (Sedano et al., 2005; Taiz y Zeiger, 2006). La fase de crecimiento lenta fue desde la siembra hasta los 24 dds. De acuerdo a las diferencias estadísticas, a partir de los 18 dds se observaron dos patrones de crecimiento: las plántulas cultivadas en el túnel de polietileno tuvieron mayor materia seca que bajo las otras condiciones de cultivo (Figura 1A).
Después de 24 dds se observó una fase de acumulación logarítmica de la biomasa seca en los tres tratamientos, y a partir de 31 dds fue evidente una fase estacionaria del crecimiento de las plántulas bajo el túnel de malla–sombra y el testigo; en cambio, el crecimiento rápido continuó bajo el túnel de polietileno (Figura 1A). A los 35 dds, las plántulas de este tratamiento habían alcanzado las características recomendadas para ser trasplantadas (Anónimo, 2005), pero las plántulas de los otros dos tratamientos no alcanzaron el tamaño recomendado durante el periodo de experimentación.
El proceso de emergencia de las plántulas de viola, fue más lento; se observaron dos tendencias de crecimiento, como en estátice, pero menos evidentes, desde los 28 dds (Figura 1B). A partir de 35 y 43 dds, las plántulas en los túneles de plástico y de malla–sombra, respectivamente, iniciaron su crecimiento exponencial, lo que llevó a las plántulas del primer tratamiento a alcanzar el tamaño recomendado para el trasplante (Anónimo, 2005) a los 52 dds; las plántulas del túnel de malla–sombra y a la intemperie tampoco alcanzaron el tamaño recomendado durante el periodo de experimentación. La cinética de crecimiento de las plántulas del testigo (intemperie) se mantuvo en la fase inicial de una curva sigmoide sin alcanzar la fase de crecimiento exponencial (Figura 1B).
La cinética de crecimiento del área foliar por planta (AF) en ambas especies fue similar al comportamiento de biomasa seca (Figuras 1C y D). Esto es, que en el túnel de polietileno se produjeron las plantas con mayor área foliar o capacidad fotosintética para abastecer de fotoasimilados a las raíces y permitir un crecimiento rápido después del trasplante (Larson, 1992). A la intemperie, la superficie foliar de las plántulas de estátice disminuyó a los 35 dds debido a la senescencia prematura de hojas.
En el túnel de polietileno, la plántula de estátice podría trasplantarse a los 35 dds con biomasa seca de 88 mg·plántula–1 y 15.1 cm2·plántula–1 de área foliar, lo cual fue el doble del tamaño de las plántulas de viola a los 52 dds, cuando tenían un peso de 43.6 mg·plántula–1 y AF de 7.6 cm2·plántula–1, lo que muestra una mayor velocidad de crecimiento de las plántulas de estátice.
Tanto en el tratamiento a la intemperie como bajo malla–sombra se registraron menores acumulaciones de biomasa seca en ambas especies (Figura 1A y 1B), siendo en estos tratamientos donde se observaron ocho y tres días con heladas, respectivamente, así como las más bajas temperaturas mínimas promedio (Cuadro 1). Además, probablemente las heladas presentes a lo largo del periodo experimental en el tratamiento a la intemperie fueron la causa de la disminución de la AF en estátice a los 35 dds, pues es considerada una especie menos tolerante al frío que viola (Anónimo, 2005).
Índices de eficiencia fisiológica
La tasa absoluta de crecimiento (TAC) que se considera como una medida de la fuerza de la demanda fisiológica (Ho et al., 1989) de estátice, presentó una fase de crecimiento lenta al principio, y fue hasta los 24 dds cuando se inició una fase de crecimiento rápido (Figura 2). Las plántulas en intemperie y en el túnel con malla–sombra alcanzaron una TAC máxima de 0.9 y 1.6 mg·d–1 a los 31 dds, pero tuvieron TAC cero a los 35 dds, cuando se detuvo el crecimiento con base en su contenido de biomasa (Figura 2A), mientras que las plántulas en el túnel de plástico alcanzaron 7.4 mg·d–1.
Los valores de TAC de las plántulas de viola creciendo en el túnel de polietileno, tuvieron una fase de crecimiento mayor a partir de los 35 dds (Figura 2B); en ese muestreo el valor alcanzado fue de 0.81 mg·d–1, mientras que a los 52 dds la TAC fue 2.4 mg·d–1; superando los valores de TAC de los otros tratamientos, que no alcanzaron en ningún periodo valores superiores a 0.5 mg·d–1.
Los máximos valores de TAC de ambas especies se observaron en las plántulas creciendo dentro del túnel con cubierta de polietileno; muy probablemente esto se debió a la ausencia total de temperaturas por debajo de 0 °C y una temperatura mínima promedio más alta que en los otros tratamientos (Cuadro 1).
La tasa relativa de crecimiento (TRC), considerada como la actividad de la demanda fisiológica o velocidad con la que se acumula carbono en la plántula (Ho et al., 1989), aumentó en estátice hasta los 31 dds bajo malla–sombra y a la intemperie, pero a los 35 dds ésta fue negativa; en túnel de polietileno no hubo una tendencia clara, aunque alcanzó el máximo valor de TRC a los 31 dds (Figura 2C).
En las plántulas de viola, de los tres tratamientos estudiados, los valores de TRC alcanzaron sus valores máximos a los 28 dds (Figura 2D), disminuyendo a lo largo del periodo de medición. La máxima TRC registrada se observó en las plantas del túnel plástico (0.14 mg·mg–1·d–1), disminuyendo de manera constante hasta alcanzar los 52 dds 0.7 mg·mg–1·d–1. En los otros dos tratamientos, los valores mínimos se alcanzaron a los 43 dds, aumentando al siguiente muestreo.
De acuerdo con Faust et al. (2005) y Adams et al. (1997), la TRC disminuye cronológicamente, pero aumenta linealmente al aumentar la irradiancia fotosintética incidente; y con altas temperaturas, esta variable es inicialmente alta hasta un límite en el que declina rápidamente, causando el cese del crecimiento en una etapa relativamente temprana; mientras que a más bajas temperaturas, esta TRC es baja inicialmente, declinando más lentamente conforme el cultivo se desarrolla que a altas temperaturas, lo que permite que las plantas crezcan durante un periodo mayor con la acumulación de más biomasa, similar a lo observado principalmente en viola (Figura 2D). Esto permite contemplar la opción de cambiar estratégicamente las plántulas en ambientes diferentes, dependiendo de su etapa de crecimiento.
La tasa de asimilación neta (TAN), como estimador de la actividad fotosintética (Ho et al., 1989), fue más alta en plántulas de estátice en el túnel plástico que en los otros tratamientos (Figura 2E). En los tres tratamientos, la TAN fue máxima a los 31 dds y mostró una disminución a los 35 dds. En esta fase, la TAN de los tratamientos a la intemperie y túnel de malla–sombra disminuyeron rápidamente, como consecuencia de una disminución del área foliar (Figura 1C), provocada a su vez por la presencia de temperaturas por debajo de 0 °C (Cuadro 1).
En viola (Figura 2F), la TAN fue máxima a los 28 dds en todos los tratamientos; posteriormente hubo una disminución. En esa edad de la plántula, la TAN de las plántulas bajo el túnel con cubierta plástica y malla–sombra (1.6 y 1.3 mg·cm–2·día–1, respectivamente) fue superior a la máxima TAN encontrada en estátice durante la fase experimental, la cual se observó a los 31 dds en la condición de túnel con cubierta plástica (0.8 mg·cm–2·día–1). Los valores de TAN de viola nunca fueron negativos, lo que puede estar relacionado con su mayor tolerancia al frío.
De acuerdo con Van der Werf (1996), los principales parámetros del análisis del crecimiento vegetal pueden ser usados para entender las relaciones entre la fuente de fotosintatos y la demanda fisiológica. Durante la fase inicial del desarrollo y hasta antes de la floración, la fuerza de la demanda la constituye el crecimiento de hojas, tallos y raíces.
Determinación del crecimiento vegetal
En esta investigación, fue posible hacer germinar y producir plántulas de estátice y viola en condiciones invernales. El ambiente menos favorable fue a la intemperie, en donde se observaron temperaturas mínimas más bajas y mayor número de heladas (Cuadro 1) que provocaron daños irreversibles en el aparato fotosintético de las plantas de estátice, además de observarse senescencia prematura de hojas; el crecimiento de plántulas fue bajo, y al final del periodo de evaluación se observaron TRC y TAN negativas. En viola, la TRC y la TAN fueron cercanas a cero. De acuerdo con la literatura, esta especie tiene una temperatura base de 4.1 °C y es considerada una especie tolerante al frío (Blanchard y Runkle, 2011), por lo que la reducción en el crecimiento fue provocada posiblemente por otros factores, como estrés hídrico o irradiancias altas. Es claro que mientras estátice tiene una tasa de crecimiento mayor, al mismo tiempo es más sensible a las temperaturas bajas y de congelamiento que viola.
El ambiente más favorable fue bajo cubierta plástica, que permitió obtener la mayor biomasa seca, área foliar, TRC, TAN y TAC en ambas especies (Figuras 1 y 2). Bajo este ambiente, la temperatura promedio del aire fue 19.5 °C (Cuadro 1), la que, considerando la magnitud de la desviación estándar de los valores de esta condición, podría ser considerada similar a la temperatura del aire medida a la intemperie, siendo además superior en 3.7 °C al registro en el túnel de malla–sombra. A pesar de existir una alta oscilación térmica de 35.6 °C entre las temperaturas máxima promedio y mínima promedio (Cuadro 1) y a que la temperatura máxima del aire fue 10 °C mayor bajo el túnel de polietileno que en los otros tratamientos lo que fue superior en 6.5 °C a lo indicado por Taiz y Zeiger (2006) como temperatura máxima óptima de crecimiento para especies originarias de climas templados, como las especies estudiadas, no se registraron temperaturas menores a 0 °C, beneficiando a ambas especies. Se ha encontrado que la mayor biomasa seca de la parte aérea de viola se obtiene a 20 °C (Adams et al., 1997), similar al promedio de temperatura a la intemperie y bajo cubierta plástica, mientras que la acumulación de materia seca es más lenta a temperaturas más calientes o más frías; la temperatura óptima estimada por Adams et al. (1997) para la TRC instantánea fue 25.3 °C y fue constante a través del tiempo. Sin embargo, estos autores señalan que la máxima acumulación de biomasa seca se presenta a altas temperaturas durante el crecimiento inicial y a bajas temperaturas durante el crecimiento posterior, lo cual implica la disminución del potencial fotosintético, un aumento del grado de autosombreo o la disminución de la relación de área foliar con la edad biológica de la planta. En viola Warner y Erwin (2006) encontraron disminución de biomasa seca hasta en 54 % (de 101 a 46 mg·d–1) de ocho cultivares cuando la temperatura aumentó de 20 a 30 °C, mientras que la TRC disminuyó con el tiempo, más que por efecto de la temperatura.
Bajo malla–sombra, la temperatura media fue 15.8 °C (Cuadro 1), lo cual fue 4 °C inferior a la temperatura óptima sugerida por Adams et al. (1997) para viola, y en la presente investigación se observó que ambas especies tuvieron menores valores en las variables respuestas medidas. Esto implica que las ventajas relativas de una malla–sombra, como son el aumento de la radiación difusa, aumento de la humedad relativa y la dispersión de la radiación ultravioleta (Stamps, 2009), no compensaron la disminución de la temperatura media. También la disminución de la irradiancia fotosintética, que integrada fue de 5.6 MJ·m–2·d–1, aunque este nivel de irradiancia fotosintética lo consideran Nemali y Van Iersel (2004) como un valor alto en especies como petunia y begonia, ya que encontraron que la biomasa de la parte aérea fue dos a tres veces mayor en plantas que crecieron a 5 MJ·m–2·d–1 que a 2.1 MJ·m–2·d–1. Sin embargo, estas plantas que crecieron en bajo nivel de radiación, aumentaron la relación de área foliar, aunque las que crecieron a altas irradiancias tuvieron las más altas tasas de crecimiento (Nemali y Van Iersel, 2004). En varias investigaciones es consistente el efecto del incremento de la irradiancia en el aumento de la biomasa seca de plántulas o estacas (Faust et al., 2005; Lopez y Runkle, 2008). Esto hasta cierto límite, en nuestra investigación, el mejor ambiente, con cubierta plástica con 30 % de sombreo, permitió ingresar 7.8 MJ·m–2·d–1 y el peor ambiente (testigo) hasta 11.2 MJ·m–2·d–1 en promedio, lo cual confirma que el efecto limitante fue la temperatura mínima y la presencia de temperaturas menores a 0 °C, y que en este caso no hubo un efecto proporcional en el incremento de biomasa conforme aumentó la irradiancia fotosintética.
En las condiciones en que se hizo el presente estudio, la irradiancia fotosintética no fue una limitante para el crecimiento, pues inclusive en invierno se alcanzan niveles de esta variable que en latitudes altas son consideradas como altas. Sin embargo, la limitación es, con frecuencia, la ocurrencia de heladas invernales.
CONCLUSIONES
Las condiciones ambientales generadas por el túnel con cubierta de polietileno, permitieron un mayor porcentaje de emergencia y mayor crecimiento de plántulas de estátice y viola, con lo que se obtuvieron plántulas para el trasplante a los 35 y 52 días después de la siembra (dds), respectivamente. El mayor crecimiento de las especies estudiadas en el túnel cubierto con polietileno se atribuye a la ausencia de temperaturas iguales o inferiores a 0 °C.
LITERATURA CITADA
ADAMS, S. R.; PEARSON, S.; HADLEY, P. 1997. An analysis of the effects of temperature and light integral on the vegetative growth of pansy cv. Universal Violet (Viola x wittrockiana Gams.). Annals of Botany 79: 219–225. [ Links ]
ANÓNIMO. 2005. Catalog of Ball Horticultural Company. Panamerican Seed Company, PanAmSeed.com. California, USA. 326 p. [ Links ]
ANÓNIMO. 2010. Anuario del Sistema Integral de Información Agroalimentaria y Pecuaria. Secretaria de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación (SAGARPA). www.siap.sagarpa.gob.mx/ar_comdeanuadin.html. Consultado el 25 de julio de 2010. [ Links ]
AYALA–SIERRA, A.; zVALDEZ–AGUILAR, L. A. 2008. El polvo de coco como sustrato alternativo para la obtención de plantas ornamentales para trasplante. Revista Chapingo Serie Horticultura 14: 161–167. [ Links ]
BLANCHARD, M. G.; RUNKLE, E. S. 2011. Quantifying the thermal flowering rates of eighteen species of annual bedding plants. Scientia Horticulturae 128: 30–37. [ Links ]
BROWN, R. 2010. Design with Microclimate: The Secret to Comfortable Outdoor Space. Island Press. Washington D.C. USA. 192 p. [ Links ]
FAUST, J. E.; HOLCOMBE, V.; RAJAPAKSE, N. C.; LAYNE, D. R. 2005. The effect of daily light integral on bedding plant growth and flowering. HortScience 40: 645–649. [ Links ]
FLORES–ALMARAZ, F.; LAGUNES–TEJEDA, A. 1998. La Horticultura Ornamental en México. Instituto Nacional de Estadística, Geografía e Informática–Colegio de Postgraduados. Aguascalientes, Ags. México. 88 p. [ Links ]
GARCÍA, E. 1981. Modificaciones al Sistema de Clasificación Climática de Köppen. UNAM. México, D.F. 168 p. [ Links ]
HO, L. C.; GRANGE, R. I.; SHAW, A. F. 1989. Source/sink regulation. In: Transport of Photoassimilates. p: 306–343. BAKER, D.; MILLBURN, J. A. (eds.). Longman Scientific and Technical. Essex, U.K. [ Links ]
HUNT, R. 1990. Basic Growth Analysis. Unwing Hyman. London, U. K. 112 p. [ Links ]
HUNT, R.; CAUSTON, D. R.; SHIPLEY, B.; ASKEW, A .P. 2002. A modern tool for classical plant growth analysis. Annals of Botany 90: 485–488. [ Links ]
KELLY, R. O.; DENG, Z.; HARBAUGH, B. K. 2006. Evaluation of viola cultivars as bedding plants and establishment of the best–of–class. HortTechnology 16: 167–171. [ Links ]
LARSON, R. A. 1992. Introduction to Floriculture. 2nd Edition. Academic Press, Inc. San Diego, California, USA. 407 p. [ Links ]
LÓPEZ, R. G.; RUNKLE, E. S. 2008. Photosynthetic daily light integral during propagation influences rooting and growth of cuttings and subsequent development of New Guinea impatiens and petunia. HortScience 43: 2052–2059. [ Links ]
MORA–AGUILAR, R.; ORTÍZ–CERECERES J.; MENDOZA–CASTILLO M. C.; RIVERA–PEÑA A.; LOZOYA–SALDAÑA H.; COLINAS–LEÓN M. T. 2005. Cinética de la acumulación y distribución de biomasa en genotipos de papa establecidos en condiciones de secano. Revista Chapingo Serie Horticultura 11: 135–142. [ Links ]
MORÁN, M. F. 2004. Producción de Plantas Ornamentales en Maceta en Invernadero. Centro de Agronegocios Tezoyuca. FIRA Banco de Tezoyuca, Morelos. 55 p. [ Links ]
NEMALI, K. S.; VAN IERSEL, W. 2004. Light intensity and fertilizer concentration: II. Optimal fertilizer solution concentration for species differing in light requirement and growth rate. HortScience 39: 1293–1297. [ Links ]
OTHA, Y. I.; KANADE T.; SAKAI, T. 1980. Color information for region segmentation. Computer Graphics and Image Processing 13: 222–241. [ Links ]
SAS INSTITUTE. 2003. The Analyst Application. 2nd ed. SAS Institute, Inc. Cary, NC, USA. 500 p. [ Links ]
SEDANO–CASTRO, G.; GONZÁLEZ–HERNÁNDEZ, V. A.; ENGLEMAN, E. M.; VILLANUEVA–VERDUZCO C. 2005. Dinámica del crecimiento y eficiencia de la planta de calabacita. Revista Chapingo Serie Horticultura 11: 291297. [ Links ]
STAMPS, R. H. 2009. Use of colored shade netting in horticulture. HortScience 44: 239–241. [ Links ]
TAIZ, L.; ZEIGER, E. 2006. Plant Physiology. 4a ed. Sinauer. Sunderland, Massachusetts, USA. 690 p. [ Links ]
VAN DER WERF, A. 1996. Growth analysis and photoassimilate partitioning. In: Photoassimilate Distribution in Plants and Crops. p: 1–20. ZAMSKI, E.; SHAFFER, A. A. (eds.) Marcel Dekker, Inc. New York, USA. [ Links ]
WARNER, R. M.; ERWIN, J. E. 2006. Prolonged high–temperature exposure differentially reduces growth and flowering of 12 Viola x wittrockiana Gams. cvs. Scientia Horticulturae 108: 295–302. [ Links ]

