










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Chapingo. Serie horticultura
On-line version ISSN 2007-4034Print version ISSN 1027-152X
Rev. Chapingo Ser.Hortic vol.16 n.3 Chapingo Sep./Dec. 2010
Colecta y caracterización morfológica de 'chile de agua' (Capsicum annuum L.) en Oaxaca, México
Collection and morphological characterization of 'water chilli' pepper (Capsicum annuum L.) in Oaxaca, Mexico
David Martínez–Sánchez1; Mario Pérez–Grajales2*; Juan Enrique Rodríguez–Pérez2; Esaú del Carmen Moreno Pérez2
1 Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional (CIIDIR). Unidad Oaxaca, Instituto Politécnico Nacional. Hornos 1003, Santa Cruz Xoxocotlán, Oaxaca. C. P. 71230. MÉXICO Correo–e: perezgm@hotmail.com (*Autor Responsable).
2 Departamento de Fitotecnia, Universidad Autónoma Chapingo. km 38.5 Carretera México–Texcoco. Chapingo, Estado de México, C. P. 56230. MÉXICO.
Recibido: 29 de septiembre, 2009.
Aceptado: 25 de octubre, 2010.
Resumen
El estado de Oaxaca, México, tiene zonas productoras de 'chile de agua' (Capsicum annuum L.) que se diferencian por sus características fisiográficas y agroecológicas. Con el objetivo de generar conocimiento que contribuya a la conservación y mejoramiento genético del potencial productivo de esta especie, en ocho zonas se colectó germoplasma y caracterizó in situ durante 2006 y 2007 bajo un esquema de muestreo completamente aleatorio, con dos localidades por zona y diez plantas por localidad, para lo cual se empleó la guía del Instituto Internacional de Recursos Fitogenéticos. Durante 2008, para disminuir el efecto ambiental, se llevó a cabo una caracterización morfológica ex situ en condiciones de invernadero; para ello las ocho colectas (360 plantas) fueron cultivadas bajo un sistema intensivo en hidroponía, en un diseño experimental completamente al azar; la información se obtuvo de ocho muestreos efectuados cada quince días, en los cuales se identificaron descriptores de raíz, tallo, flor, fruto y semilla, que permitieron diferenciar (selección de variables) las colectas e identificarlas en tres grupos (análisis de agrupamiento de mínima varianza de Ward) y describir las características responsables de la agrupación (análisis de componentes principales). El germoplasma de la zona de Ocotlán, Oaxaca, resultó promisorio para un proceso de mejoramiento genético por presentar los frutos con los caracteres más deseables para el mercado.
Palabras clave: morfología, descriptores de Capsicum, germoplasma, análisis multivariado.
Abstract
Oaxaca state of Mexico has a number of zones with 'water chilli' pepper, (Capsicum annuum L.) which differ in their physiographic and agro–ecological characteristics. With the objective of to generate knowledge that contributes to the conservation and genetic improvement of the productive potential of this species of chilli, in eight zones, germplasm was collected and characterized in situ during 2006 and 2007 under a completely random sampling using two locations and ten plants per location, which used the guide to the International Plant Genetic Resources Institute. To decrease the environmental effect of the collection sites, was carried out ex situ morphological characterization, during 2008, under greenhouse conditions, therefore the eight collections (360 plants) were grown under an intensive system in hydroponics, in a design completely randomized. The information was extracted from eight samples taken every fifteen days. From analysis of variance and covariance were identified descriptors root, stem, flower, fruit and seed, to distinguish collections. In the cluster analysis of cluster (Ward minimum variance) and describe the characteristics of the grouping responsible (principal components analysis) were identified three groups and Ocotlán germplasm as promising for a process of genetic improvement by to present the fruits with the characteristics most desirable for the market.
Key words: Morphology, Capsicum descriptors, germplasm, multivariate analysis.
INTRODUCCIÓN
La falta de conocimiento de la diversidad genética que conservan los productores en sus parcelas de cultivo, conlleva a un limitado conocimiento sobre la distribución geográfica del género Capsicum en México y de la variabilidad de las especies silvestres cultivadas (Latournerie et al., 2002). Es poco el trabajo que se ha hecho en la protección de las especies silvestres, o domesticadas en sus hábitats naturales. Algo muy importante en los procesos de conservación in situ es conocer y comprender los factores que influyen en la toma de decisiones de los campesinos para seguir conservando sus recursos genéticos (Latournerie et al., 2002), entre los cuales se encuentra el 'chile de agua' (Capsicum annuum L.) cultivado en el estado de Oaxaca.
Esta hortaliza tiene importancia económica y social en los Valles Centrales de Oaxaca, donde se le cultiva y consume ampliamente (López y Castro, 1999; Velasco et al., 1998). Los sistemas agrícolas tradicionales han permitido la diversidad morfológica de este tipo de chile, el cual tiene un ciclo vegetativo de aproximadamente cuatro meses y su fruto es apreciado por su sabor y picor en la preparación de platillos típicos. No obstante, el rendimiento y calidad de fruto es limitado por la falta de variedades mejoradas y el bajo desarrollo tecnológico. El mejoramiento de este tipo de chile ha sido a través de la selección que los productores han llevado a cabo en sus propios cultivos; así, los materiales obtenidos presentan alta variabilidad aún no caracterizada, condición necesaria para iniciar un proceso formal de mejoramiento genético, de mediano y largo plazos, que genere variedades propias para la región (López y Castro, 1999).
Debido a las situaciones mencionadas yala necesidad de contar con información de los caracteres morfológicos que expliquen la variabilidad presente en los ecotipos de 'chile de agua' en las diferentes zonas donde se cultivan, Tapia et al. (2005) consideran la utilidad de una caracterización morfológica de los genotipos de los recursos vegetales, pues permiten definir una serie de descriptores útiles por su importancia agronómica, o por su valor para clasificar e identificar grupos genéticos. Las características morfológicas han sido usadas para la identificación de especies, familias y géneros de plantas, las cuales involucran factores de resistencia a enfermedades y plagas y de rendimiento, entre otros (Falconer y Mackay, 1996).
Por la importancia de contar con patrones de identificación, caracterización y evaluación de la mayoría de las plantas cultivadas, se han realizado estudios básicos para conocer la variabilidad de las características dentro y entre plantas y seleccionar las características cualitativas o cuantitativas que sean útiles para la descripción (Pérez et al., 2004). Los órganos más importantes para la descripción morfológica son aquellos que están menos influidos por el ambiente, como la flor y el fruto, así como las hojas, tallos, raíces y los tejidos celulares (Raven et al., 2005).
Con el objetivo de generar conocimiento que contribuya a la conservación y mejoramiento genético del chile de agua, se colectó y caracterizó morfológicamente in situ el germoplasma de diferentes localidades de los Valles Centrales de Oaxaca; para evitar el efecto ambiental de los sitios de colecta, también se hizo una caracterización morfológica ex situ en condiciones de invernadero, bajo la hipótesis de que existe variabilidad genética que permite la distinción entre colectas.
MATERIALES Y MÉTODOS
Colecta de germoplasma
Durante 2006 y 2007 se efectuaron recorridos de campo en localidades con tradición en el cultivo de chile de agua, las cuales, por sus características fisiográficas y agroecológicas, se agruparon en diferentes zonas (INEGI, 2008). En dos localidades de cada zona elegida se colectaron frutos maduros de diez plantas por localidad, seleccionadas de manera aleatoria. La ubicación geográfica de las zonas de colecta, así como sus características de clima, se presenta en el Cuadro 1.
Caracterización morfológica in situ
Las colectas fueron caracterizadas in situ de acuerdo con los descriptores de Capsicum spp (IPGRI, 1995), donde se consideraron 70 caracteres entre cuantitativos y cualitativos. Los caracteres constantes no fueron incluidos en el análisis de caracterización, por lo que sólo se analizaron 46 variables.
Caracterización morfológica ex situ
Con el propósito de evitar el efecto ambiental se hizo caracterización morfológica ex situ, donde se empleó la mezcla de las semillas colectadas de las dos localidades, de cada una de las ocho zonas. El estudio se llevó a cabo de junio a septiembre de 2008 en un invernadero de estructura metálica y vidrio, en la Universidad Autónoma Chapingo, donde la condición promedio de temperatura, humedad relativa y radiación fue 21 °C, 69.5 %, y 950 emoles de fotones·m–2·s–1, respectivamente. El cultivo se manejó en bolsas de polietileno negro calibre 600, con espuma volcánica (tezontle) como sustrato, sistema de tutoreo con rafia, sistema de fertirrigación con solución nutritiva de Steiner (1984), control de plagas con trampas adherentes de color amarillo y azul, y el de enfermedades con fungicidas preventivos a base de oxicloruro de cobre y azufre (Ruíz y Medina, 2001).
Cada una de las ocho colectas manejadas como tratamientos, fueron establecidas bajo un diseño completamente al azar con cuatro repeticiones, cada una con once plantas. Los descriptores empleados para é caracterización también fueron 46, y para su descripción se siguió la guía del IPGRI (1995). Se realizó una selección de caracteres que permitió la discriminación de colectas mediante análisis de varianza y covarianza sucesivas; posteriormente, con las variables elegidas se efectuó un análisis de agrupamiento mediante el uso de distancia euclidiana al cuadrado y la técnica de mínima varianza de Ward (Johnson, 2000); la altura de corte fue determinada mediante el criterio cúbico de agrupamiento (CCC), la pseudoestadística T cuadrada de Hotelling (PST2 ) y la pseudo F (Johnson, 2000). Un análisis de Componentes Principales (CP) se aplicó para corroborar la agrupación obtenida y determinar los caracteres responsables de ésta; esto con el apoyo del paquete estadístico SAS V. 9.0.
RESULTADOS Y DISCUSIÓN
Caracterización morfológica in situ
Según el IPGRI (1995) y González (2001), en la caracterización morfológica se deben registrar aquellos caracteres altamente heredables, que pueden ser fácilmente evaluados a simple vista y se expresan en todos los ambientes. En chile de agua se encontraron 21 caracteres constantes (Cuadro 2), los cuales pueden ser considerados de herencia simple (Azofeifa, 2006) y, por lo tanto, empleados como marcadores en este tipo de chile.
Se encontraron frutos con ápice redondo, agudo o hendido incluso en la misma planta, así como frutos con pedúnculos con inclinación desde 0 hasta 90°, descripción que coincide con López y Castro (1999).
Caracterización morfológica ex situ
Mediante análisis de covarianza sucesivos fueron seleccionados los caracteres que presentaron variabilidad (Johnson, 2000). El criterio de selección se basó en conservar los caracteres que presentan variabilidad que permita la discriminación efectiva de colectas, y eliminar aquéllos cuya variación esté altamente correlacionada con los que son constantes. Así, de 46 caracteres, once tuvieron significancia estadística y contribuyeron ampliamente en la caracterización del chile de agua (Cuadro 3).

Los descriptores seleccionados muestran que existe una distribución de la variabilidad en la planta, con un carácter asociado a la raíz, tres al tallo, dos a la hoja, cuatro al fruto y cinco a la flor; asimismo, en forma general se observó que dentro de las ocho colectas caracterizadas hubo poca variabilidad entre ellas, no así entre plantas donde los principales órganos que mostraron mayor variabilidad fueron flor y fruto.
De acuerdo con el criterio cúbico de agrupamiento (CCC) y la pseudoestadística T cuadrada de Hotelling (PST2) (Johnson, 2000), a partir de 15 variables seleccionadas (Cuadro 3) se formaron tres grupos.
El Análisis de Componentes Principales (ACP), indicó que los cuatro componentes principales (CP) explicaron 85 % de la variabilidad total (Cuadro 4).

El primero CP1(32 % de la variabilidad total) representa a los caracteres días a la floración, días a fructificación y densidad de ramificación; al CP2 (27 % de variación) corresponden las variables: longitud de la hoja cotiledón, la relación diámetro del fruto/grosor del pericarpio y longitud de la corola; el CP3 (15 % de la variabilidad), las variables ancho de la corola, longitud de la antera y longitud de fruto, mientras que el CP4 (11 % de variación) representa a longitud del hipocótilo y la excersión del estigma (Cuadro 5). (Figura 1)
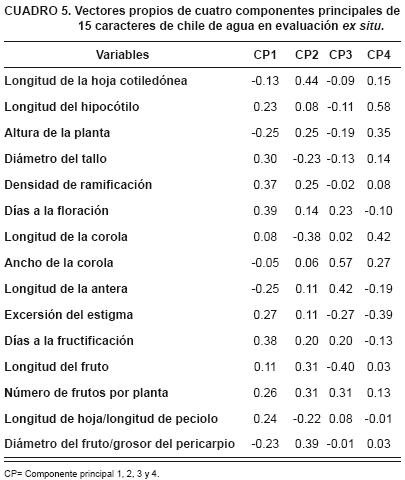
Los caracteres seleccionados por el análisis (CP) son importantes en el proceso de mejoramiento de esta especie, el cual consiste en seleccionar aquellos individuos que dispongan de caracteres deseables en el mayor grado posible, desechando los de grado menor, para que en programas de mediano y largo plazos y a través de selecciones repetidas se puedan alcanzar las expectativas de la mejora deseada (Poehlman y Allen, 2003).
Por medio de la mejora vegetal utilizando la información de los caracteres o descriptores que mostraron la mayor variabilidad en el análisis (CP), y de acuerdo a los principios de la genética, es posible producir variedades hortícolas con características más deseables, tales como mayor resistencia a las enfermedades, mejores valores nutricionales, sabores más agradables e intensos, mayor rendimiento, etc. Investigadores dentro del campo del mejoramiento de plantas y con la información de los caracteres morfológicos y agronómicos a su alcance, han conseguido acelerar el proceso de mejora y selección natural mediante selección artificial y orientando los resultados hacia la obtención de plantas deseables por determinadas características (Howell, 1998).
La dispersión de las poblaciones se representa en un plano bidimensional formado por los componentes principales 1 y 2 (Figura 2); se aprecia que las colectas de chile de agua corresponden a los tres grupos generados por el análisis de agrupamiento. El grupo I incluye las colectas 1, 7 y 8 (de diferentes zonas de Oaxaca, Tlacolula y Yautepec, respectivamente), que se distinguen por ser precoces, con poca densidad de ramificación, hojas cotiledonares pequeñas, pericarpio grueso y corolas largas. El grupo II contiene cuatro colectas de las cuales la 4 y la 6 (colectadas en Etla y Sola de Vega) son tardías, con mayor ramificación, hojas cotiledonares largas, pericarpio delgado y corola corta, y la colecta 5 (proveniente de Zaachila) es tardía, con mayor ramificación, hoja cotiledónea pequeña, pericarpio grueso y corola larga. La colecta 2 (de Zimatlán) no presenta los caracteres distintivos de este grupo, debido a que los caracteres longitud del hipocótilo, longitud del fruto y la relación hoja del fruto/longitud del peciolo presentan valores muy altos en alguno de los grupos, que los alejan de la media. En el grupo III, la colecta 3 (procedente de Ocotlán) es precoz, con poca ramificación, hoja de cotiledón larga, pericarpio grueso y corola corta. Las características ambientales de las zonas de origen de las colectas, no influyeron sobre la caracterización morfológica realizada con las variables analizadas en la presente investigación.
Estos resultados se asemejan a los reportados por Latournerie et al. (2002), Chávez y Castillo (1999), González y Bosland (1991), quienes para diversas especies de Capsicum encontraron que los tres primeros componentes explicaron el mayor porcentaje de la variabilidad, y los descriptores de fruto fueron los más importantes debido principalmente a las prácticas tradicionales de cultivo, a la recombinación genética y a la selección natural. López y Castro (1999), en colectas nativas de Oaxaca, determinaron que los caracteres longitud del limbo, ancho del limbo, longitud del pedúnculo de la hoja, longitud del fruto, ancho del fruto y el diámetro de la semilla presentaron una mayor contribución a la varianza global.
Con base en descriptores cualitativos y agronómicos, Sudre et al. (2010) caracterizaron 26 especies de Capsicum y mencionan que los genotipos observados presentan potencial para su uso en distintos sectores de producción; asimismo, encontraron que en las especies estudiadas cinco fue el número ideal de grupos por los criterios pseudo–F y pseudo–t2. El procedimiento de Ward–MLM permitió la diferenciación de las especies C. annuum, C. frutescens, C. baccatum y C. chinense, y mostró cierto grado de eficiencia en el agrupamiento.
En la caracterización morfológica de 93 accesiones de Capsicum spp, Palacios y García (2008) confirmaron la variabilidad dentro del género, en especial los descriptores de arquitectura de planta, estructuras reproductivas y producción, que explicaron el 78 % de la variabilidad total, con base en componentes principales; las distancias genéticas posibilitaron formar grupos con base en el origen de las accesiones y caracteres de flor y fruto, pero no discriminaron entre especies; por lo demás, las cortas distancias genéticas resultantes del análisis discriminante entre C. annuum, C. frutescens y C. chinense indicaron que conforman un solo grupo morfológico.
En el Cuadro 6 se presentan las medias de los caracteres por grupo, el nivel de significancia de las variables dentro de cada grupo, medias generales y los coeficientes de variación por descriptor.
Los caracteres longitud del hipocótilo con valores entre 2.61 y 3.50 mm, densidad de ramificación con variación entre 5.24 y 6.00 (intermedia), el ancho de la corola con fluctuación entre 1.96 y 2.14 mm, y el número de frutos por planta que osciló entre 17.24 y 18.84, no presentaron diferencia significativa entre grupos (Tukey, P<0.05). Los valores obtenidos para estos caracteres concuerdan con los reportados por López y Castro (1999), donde mencionan que la densidad de ramificación predominante para este cultivo es intermedia y que está relacionada con su hábito de crecimiento erecto, tallos de forma cilíndrica y dicotómica.
La longitud de la hoja de cotiledón con valores que variaron entre 9.0 y 16 mm, el diámetro del tallo con valores entre 8.54 y 10.13 mm, la longitud de la corola con fluctuaciones entre 0.88 y 1.03 mm y la relación diámetro de fruto/grosor de pericarpio, entre 13.32 y 18.57, mostraron diferencia significativa (Tukey P<0.05) del grupo 3 contra los grupos 1 y 2; entre estos últimos no hubo diferencias. De alguna manera estos resultados contrastan con algunos de los obtenidos por López y Castro (1999) para diversas colectas de chiles nativos de Oaxaca, en los que mencionan que los caracteres cuantitativos: longitud de limbo, ancho de limbo, longitud hasta la primera ramificación, longitud del pedúnculo de la hoja y la longitud del fruto, presentaron los coeficientes de variación de mayor magnitud e intervalos de 4.6 a 11 cm, 2.8 a 8.5 cm., 1 a 14 cm, 2 a 10.5 cm y de 1 a 8.5 cm, respectivamente.
La altura de la planta con valores que fluctuaron entre 108 y 118 cm, la excersión del estigma donde los valores cambiaron entre 5.08 y 6.60, la longitud del fruto de 9.14 a 10.40 cm y en la relación longitud de hoja/longitud de peciolo entre 1.66 y 2.41, mostraron diferencia significativa (Tukey P<0.05) entre los tres grupos. Los valores de altura de la planta son mayores a los reportados por López y Castro (1999), quienes encontraron en condiciones de campo promedios de altura de 64.53, 64.31, 56.55 y 56.93 cm. La diferencia entre alturas de la planta se debe a que en condiciones de cultivo en invernadero existe un mayor control en los factores ambientales y manejo del cultivo que influye en el crecimiento y desarrollo. Perera y Poulus (1993) al evaluar la posición del estigma en 17 colectas de chile encontraron el estigma arriba, abajo y al mismo nivel de las anteras, lo cual atribuyeron a la heterogeneidad de las plantas; mencionan también que en la longitud del pistilo y los estambres influye el efecto del ambiente.
Para el carácter días a la floración donde los valores fluctuaron entre 43.20 y 47.20, la longitud de la antera con valores que oscilaron entre 1.84 y 2.00 mm y en días a la fructificación el rango de variación estuvo entre 63.48 y 67.90, hubo diferencia significativa (Tukey P<0.05) entre el grupo 2 respecto a los grupos 1 y 3 que no mostraron variación entre ellos. Estos datos concuerdan con los reportados por Ruíz y Medina (2001) en diversas colectas de chile de agua donde obtuvieron, para días a floración, un promedio de 37.8 para colectas precoces y 49.5 para colectas tardías, y también mencionan un promedio de 75.5 días para cosecha en diversas colectas de chile de agua.
CONCLUSIONES
Los resultados de la caracterización morfológica in situ y ex situ mostraron que en las colectas bajo estudio, hubo 21 caracteres sin variación y 46 que presentaron variabilidad. De éstos, se identificaron cuatro componentes principales (CP) que explicaron 85 % de la variación total, atribuida a 15 caracteres morfológicos.
El germoplasma de Ocotlán es el más promisorio para un proceso de mejoramiento genético, por presentar frutos con caracteres deseables para el mercado.
LITERATURA CITADA
AZOFEIFA D., A. 2006. Uso de marcadores moleculares en plantas; aplicaciones en frutales del trópico. Agronomía Mesoamericana 17(2): 221–242. [ Links ]
CHAVÉZ, S. J. L.; CASTILLO, G. F. 1999. Variabilidad en caracteres morfológicos de colectas de chile manzano (Capsicum pubescens R y P). Rev. Fitotecnia Mexicana. 22: 27–41. [ Links ]
FALCONER, D. S.; MACKAY T., F. C. 1996. Introduction to Quantitative Genetics, Ed. 4. Longmans Green, Harlow, Essex, UK. pp 2235–240. [ Links ]
GONZÁLEZ, A. F. 2001. Caracterización morfológica. In: Conservación y caracterización de recursos fitogenéticos. GONZÁLEZ–ANDRÉS, F.; PITA–VILLAMIL, J. M. (Eds). Publicaciones I. N. E. A. Valladolid, España. 279 p. [ Links ]
GONZÁLEZ, M. M.; BOSLAND, P. W. 1991. Strategies for stemming genetic erosion of Capsicum germoplasm in the Americans Diversity 7: 52–53. [ Links ]
HOWELL, S. H. 1998. Genética de plantas y su desarrollo. Cambridge Univ. Press. M. A. Estados Unidos. pp. [ Links ]
INEGI. 2008. Sistema para la consulta del Anuario estadístico del estado de Oaxaca. México en: http.//www.inegi.org.mx/est/contenido/español/sistemas/aeeo3/estatal/oax/index.htm. Consultado: 10 de marzo de 2009. [ Links ]
IPGRI. 1995. Descriptors for Capsicum spp. Instituto Internacional de Recursos Fitogenéticos (IPGRI), Secretariat, Rome, Italy. 32 p. [ Links ]
JOHNSON D. E. 2000. Métodos multivariados aplicados al análisis de datos. Traducido por Pérez Castellanos H. Internacional Thomson Editores. México. 566 pp. [ Links ]
LATOURNERIE, M. L.; CHÁVEZ, S. J. L.; PÉREZ, P. M.; CASTAÑÓN, N. G.; RODRÍGUEZ, H. S. A.; ARIAS, R. L. M.; RAMÍREZ, V. P. 2002. Valoración in situ de la diversidad morfológica de chiles (Capsicum an–nuum L. y Capsicum chinense Jacq) en Yaxcaba, Yucatán. Rev. Fitotecnia Mexicana 25 (1): 25–33. [ Links ]
LÓPEZ, L. P. S.; CASTRO, G. F. H. 1999. Al rescate de la diversidad del chile (Capsicum spp) en Oaxaca. Publicación especial. Campo Experimental Valles Centrales de Oaxaca. INIFAP, Oaxaca. México. 7 pp. [ Links ]
PALACIOS, C. S.; GARCÍA, D. M. A. 2008. Caracterización morfológica de 93 accesiones de Capsicum spp del banco de germoplasma de la Universidad Nacional de Colombia Sede Palmira. Acta agronómica. 57 (4): 247–252. [ Links ]
PERERA, O.; POULUS. J. M. 1993. Studies on stigma position of seventeen pepper accessions. Capsicum and Eggplant Newsletter 12: 44–46. [ Links ]
PÉREZ, G. M.; GONZÁLEZ, H. V. A.; MENDOZA, C. M. C. ; PEÑA, V. C. 2004. Physiological characterization of manzano hot pepper (Capsicum pubences R & P) landraces. J. Amer. Soc. Hort. Sci. 129(1): 88–92. [ Links ]
POELHMAN, J., ALLEN, D. 2003. Mejoramiento genético de las cosechas. Edit. Limusa. 2da Edición. México, D. F. 511 p. [ Links ]
RAVEN, P. H.; R. F. VUELQUE; S. E. EICHHORN, 2005. Biología de plantas. 7ma. ed., Nueva York: W. H. Freeman. Book News, Inc., Portland, OR. pp 359–464. [ Links ]
RUÍZ, V. J.; MEDINA, Z. J. 2001. Avances en el manejo integrado de Bemisia tabaci en tomate y chile en Oaxaca, México. Manejo Integrado de plagas (Costa Rica) 59: 32–38. [ Links ]
STEINER, A. 1984. The Universal Nutrient Solution. In: Proceedings of the 6th International Congress on Soilles Culture. ISOSC. Wageningen, The Netherlands. pp. 633–649. [ Links ]
SUDRÉ P.C.; GONCALVES A. S. L.; R. RODRIGUEZ, R.; A. T. DO AMARAL JUNIOR A. T.; RIVA–SOUZA E. M; BENTO, C. DOS S. 2010. Genetic variability in domesticated Capsicum spp as assessed by morphological and agronomic data in mixed statistical analysis. Genet. Mol. Res. 9 (1): 283–294. [ Links ]
TAPIA, C. E.; GUTIÉRREZ, E. M. A.; WARBURTON, M. L.; SANTACRUZ, V. A.; VILLEGAS, M. A. 2005. Characterization of mandarin (Citrus spp.) using morphological and AFLP markers. Interciencia 30 (11): 687–693. [ Links ]
VELASCO, V. V. A.; TRINIDAD S. A.; TIRADO, T. J. L.; TÉLIZ, O. D.; MARTÍNEZ, G. A.; CADENA, H. M. 1998 Efecto de algunos nutrimentos en plantas de chile de agua infectadas con virus. Terra Latinoamericana. 16(4): 317–324. [ Links ]

