










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista Chapingo. Serie horticultura
versão On-line ISSN 2007-4034versão impressa ISSN 1027-152X
Rev. Chapingo Ser.Hortic vol.16 no.3 Chapingo Set./Dez. 2010
Efecto de NaCl y CaCl2 en el desarrollo de ejes embrionarios de Persea americana Mill. criollo y 'Hass' cultivados in vitro
Effect of NaCl and CaCl2 on embryonic axes development of criollo and 'Hass' of Persea Americana Mill. growing in vitro
Leobarda Guadalupe Ramírez–Guerrero1*; Héctor González–Rosas1; Guillermo Calderón–Zavala1; Juan Velázquez Mendoza2; Victor Manuel Cetina–Alcalá2; Ana María Castillo–González3; Adriana Delgado–Alvarado4
1 Recursos Genéticos y Productividad, Colegio de Postgraduados, Campus Montecillo, km 36.5. Carretera México–Texcoco, Montecillo, Estado de México. C. P. 56230, MÉXICO.
2 Forestal, Colegio de Postgraduados, Campus Montecillo, km 36.5. Carretera México–Texcoco, Montecillo, Estado de México. C. P. 56230, MÉXICO. Correo–e: leoramirez@colpos.mx (Autor responsable).
3 Departamento de Fitotecnia, Universidad Autónoma Chapingo. km 38.5 Carretera México–Texcoco. Chapingo, Estado de México. C. P. 56230. MÉXICO.
4 Botánica, Colegio de Postgraduados, Campus Puebla. km 125.5 Carretera Federal México–Puebla Colonia La Libertad. C. P. 72130, Puebla, Pue. MÉXICO.
Recibido: 30 de marzo, 2010.
Aceptado: 17 de septiembre, 2010.
Resumen
El objetivo de este estudio fue identificar materiales de aguacate tolerantes a salinidad, útiles como portainjertos. Se sembraron in vitro ejes embrionarios de aguacate criollo y 'Hass' en medio Murashige y Skoog al 100 % de su composición nutrimental en condiciones salinas inducidas con NaCl y CaCl2. Se emplearon cinco niveles de cada sal (0.1, 0.3, 0.5, 0.7, 0.9 %), así como la combinación de éstas en proporción 1:1. Se estudió germinación, emergencia, enraizamiento y supervivencia de brotes de aguacate. El criollo superó al 'Hass' con el 21 % de germinación, 36 % de emergencia, con aproximadamente el 50 % en cada una de las variables de crecimiento y el 10 % de supervivencia de plántulas. El desarrollo de los ejes embrionarios fue más limitado en los tratamientos con NaCl en comparación a los establecidos en CaCl2 y NaCl:CaCl2. Con la técnica in vitro se determinó a los 65 días de cultivo que el material criollo fue más tolerante al estrés salino con respecto al 'Hass'.
Palabras clave: Aguacate, germinación, emergencia, sobrevivencia, salinidad.
Abstract
The aim of this study was to identify salinity tolerant avocados materials useful as rootstocks. Embryonic axes of criollo and 'Hass' avocados were planted in vitro on Murashige and Skoog medium at 100 % of its nutritional composition in induced saline environment with NaCl and CaCl2. Five levels of each salt were tested (0.1, 0.3, 0.5, 0.7, 0.9 %), as well as their combination at 1:1 ratio. The germination, seedling emergency, rooting plant growth and plant surviving of avocados were studied. Criollo achieved 21 % of seed germination, and 26 % of seedling emergency, higher than 'Hass' avocados, with 50 % in each one of plant growth variables and 10 % of plantlets survival. The embryonic axes were more deteriorated with NaCl treatments than with CaCl2 treatments and NaCl:CaCl2 treatments. The in vitro technique helped to determine, after 65 days of culture than criollo avocados were more stress saline tolerant than Hass.
Key words: Avocado, germination, emergency, plant surviving, salinity.
INTRODUCCIÓN
Los suelos afectados por sales se localizan en prácticamente todas las altitudes y regiones climáticas, por lo cual se considera a nivel mundial uno de los principales problemas que limitan la producción agrícola, que combinado con la pobre calidad del agua, restringen severamente la productividad de los cultivos (Singh y Chatrath, 2001). En las zonas productoras de aguacate, especialmente en las áreas áridas y semiáridas con irrigación, como California, Texas e Israel (Gazit y Kadman, 1976), los suelos están en mayor o menor grado afectados por procesos de salinización y donde las aguas utilizadas incrementan cada vez más su concentración salina, disminuyendo el potencial productivo de este frutal (Mickelbart et al., 2007).
En México, el aguacate (Persea americana Mill.) es un cultivo de importancia económica alta, de acuerdo a la superficie cultivada (133,441 ha), localizada principalmente en los estados de Michoacán, Jalisco, Morelos, México, Nayarit y Guerrero (Anónimo, 2010). Su distribución es afectada por ser una de las especies frutales más sensibles a la salinidad (conductividad eléctrica menor a 3 dS·m–1) principalmente por la presencia de NaCl en el suelo y en el agua de riego (Bernstein et al, 2001; Mickelbart y Arpaia, 2002). El uso irracional de los fertilizantes es una práctica agrícola que eleva transitoriamente la conductividad eléctrica o salinidad del suelo, y con ello las plantas aumentan la absorción de iones tóxicos, como el cloro y sodio, situación trascendental en la época de precipitación pluvial baja y más cuando el árbol está en crecimiento activo, limitando sus posibilidades de desarrollo, ocasionando daños y pérdidas en los huertos existentes (Salazar–García y Lazcano–Ferrat, 1999; Ben–Ya'acov y Zilberstaine, 1999; Mickelbart y Arpaia, 2002 ). La acumulación de sodio en las hojas ocasiona la muerte de los brotes, lo cual está asociado con el deterioro del sistema radical, debido a que pierde su capacidad para retener este ión, lo que facilita su distribución en varias partes de la planta. El cloro genera necrosis que puede cubrir más del 50 % de la hoja, causando su caída prematura y afectando los procesos fisiológicos como la actividad fotosintética y, en consecuencia, la producción (Salazar–García et al, 1987; Bernstein et al., 2001; Castro et al., 2009); sin embargo, la sensibilidad o tolerancia al exceso de cloro y sodio varían entre cultivares y entre portainjertos.
La relevancia de la producción mundial y nacional de aguacate, las condiciones de salinidad en los suelos y el agua de riego en las zonas agrícolas, enfatizan la necesidad del establecimiento de programas de mejoramiento genético para este cultivo. En México existe poca investigación dirigida a la obtención de materiales tolerantes o a la evaluación mediante la selección de portainjertos tolerantes a salinidad, que es una práctica para superar este problema (Ben–Ya'acov y Zilberstaine, 1999; Pire et al., 2007). Sin embargo, la producción de materiales y la evaluación de la tolerancia en experimentos de campo implican periodos mayores a 10 años, y generalmente, los resultados son afectados por muchas variables ambientales (Salazar–García et al., 1987; Ben–Ya'acov y Zilberstaine, 1999). Ante esta situación, se requiere de trabajos interdisciplinarios que, apoyados por sistemas eficientes, permitan obtener resultados a corto plazo. La investigación in vitro puede ser una opción para seleccionar materiales tolerantes a sales (Zhang, 1994). En este sentido, González–Rosas et al. (2003) y Fuentes et al. (2004) mencionan que el cultivo in vitro de embriones cigóticos maduros es una técnica útil con propósitos de selección de portainjertos tolerantes a sales en corto tiempo en relación a la variación genética natural de los embriones y su capacidad de ajuste osmótico por la acumulación de solutos. Por lo anterior, este estudio tuvo como objetivo identificar in vitro materiales de aguacate tolerantes a salinidad.
MATERIALES Y MÉTODOS
Este estudio se realizó en dos experimentos: uno con aguacate criollo y otro con aguacate 'Hass'. En ambos experimentos se utilizaron ejes embrionarios extraídos de semillas de frutos maduros de aguacate. Las semillas se lavaron con agua corriente y detergente suave; se sometieron a tratamiento desinfestante superficial con solución de etanol al 70 % durante 1 min y a enjuagues con agua destilada estéril. A la semilla se le cortaron los cotiledones que cubren el eje embrionario hasta dejarlos en forma de cubo de 1 cm por lado; a este cubo se le dio otro tratamiento desinfestante sumergiéndolo en solución de hipoclorito de sodio al 20 % durante 15 min y a enjuagues sucesivos con agua destilada estéril (González–Rosas et al., 2003); por último, se sometieron por 1 hora en solución al 2 % de KH2PO4 para minimizar la oxidación.
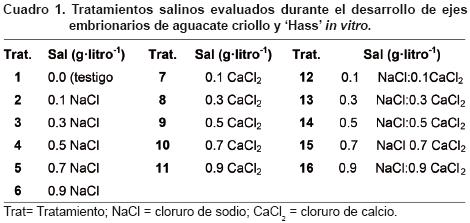
Los ejes embrionarios se sembraron en tubos de ensaye (25 x 2 cm) con 20 ml de medio de cultivo (Murashige y Skoog, 1962) al 100 % de sus sales, suplementado con 2 mg·litro–1 de glicina, 10 mg·litro–1 de mio–inositol, 4 mg·litro–1 de tiamina–HCl, 30 g·litro–1 de sacarosa y 8 g·litro–1 de agar, y la adición de los tratamientos salinos inducidos con NaCl, CaCl2 y la combinación de ambas sales en proporción 1:1, como se muestra en el Cuadro 1. El pH del medio de cultivo se ajustó a 5.7±0.1 utilizando NaOH (1N) y HCl (1N), y se esterilizó en autoclave a 121° C y 1.5 kg·cm–2 por 15 min. Después de la siembra, los tubos se colocaron en una sala de incubación a 27±1° C y 16 h de luz (50 μmol·m–2–s–1 de intensidad de luz proporcionada por lámparas fluorescentes).
Las variables evaluadas fueron: días a inicio de germinación y porcentaje de germinación, los cuales fueron contabilizados a partir de la aparición de la raíz; días a inicio de emergencia y porcentaje de emergencia registrados al momento de la aparición de la plúmula. A los 65 días de cultivo se midieron: longitud de raíz y de tallo, considerados desde la base de inicio de cada uno de ellos hasta su ápice; diámetro de raíz y de tallo, evaluados en la parte media de su longitud; número de hojas formadas y porcentaje de supervivencia de ejes embrionarios.
En los dos experimentos, el diseño experimental fue completamente al azar con cinco repeticiones. La unidad experimental fue una plántula por tubo. A los datos obtenidos se les realizó un análisis de varianza y la prueba de medias de Tukey (P<0.05) usando el paquete SAS (Statistical Analysis System) versión 9.2. Además, se realizó una prueba de t para comparar el grado de tolerancia a salinidad entre criollo y 'Hass'.
RESULTADOS
Días a inicio de germinación y porcentaje de germinación. La germinación de ejes embrionarios de aguacate 'Hass' y criollo varió significativamente (P<0.01) entre tratamientos (Cuadro 2). En comparación al testigo, tanto en criollo como en 'Hass', la germinación de los ejes embrionarios disminuyó, principalmente en el grupo de tratamientos con NaCl y en menor grado con CaCl2 y NaCl:CaCl2. En el criollo, la germinación inició a los 22 días mientras que en 'Hass' fue a los 20 días. Del total de ejes embrionarios cultivados in vitro, en criollo se logró el 60 % de germinación y en 'Hass' el 39 %.
Días a inicio de emergencia y porcentaje de emergencia. En 'Hass' y criollo, el inicio de emergencia de la plúmula fue marcadamente diferente (P<0.01) entre tratamientos (Cuadro 2). En el testigo, la emergencia de criollo fue más rápida, aunque en porcentaje similar que 'Hass' (Cuadro 3). En ambos materiales de aguacate, la emergencia fue menor en el grupo de tratamientos compuestos por NaCl y, fue favorecida con la presencia de CaCl2 en el medio. En 'Hass', la adición de NaCl detuvo tanto la germinación como la emergencia, pues en tres de las cinco concentraciones empleadas la respuesta fue limitada (Cuadro 3) y en NaCl:CaCl2, los ejes embrionarios no germinaron ni emergieron en dos de las cinco concentraciones, específicamente a altas concentraciones.
Longitud de raíz y tallo. En criollo hubo efecto significativo de tratamientos sólo para longitud de tallo; en 'Hass'; la diferencia entre tratamientos se observó en ambas variables (Cuadro 2). Tanto en 'Hass' como en criollo, la longitud de raíz y tallo fue estadísticamente superior en el testigo (Cuadro 4); sin embargo, 'Hass' logró 1.47 cm más longitud de raíz y 4 cm más de tallo que el criollo. Ambos órganos crecieron más en CaCl2 y fueron principalmente dañados por NaCl. En criollo se encontró, una proporción tallo:raíz de 2:1 y en 'Hass' fue aproximadamente 1:1.
Diámetro de raíz y tallo. Entre tratamientos, el análisis arrojó diferencias en el diámetro de raíz y tallo, en los dos materiales de aguacate (Cuadro 2). En promedio, en las plántulas de criollo, el diámetro de tallo fue ligeramente mayor que el de la raíz y en 'Hass', el diámetro de raíz fue levemente superior que el del tallo (Cuadro 4). En el testigo, la raíz de criollo fue 0.22 mm más gruesa que en 'Hass', pero en este cultivar, el diámetro de tallo fue ligeramente superior (0.06 mm) al del criollo. Las raíces y los tallos fueron más gruesos en el grupo de tratamientos compuestos por CaCl2 tanto en plántulas de criollo como de 'Hass', en comparación a los obtenidos con NaCl:CaCl2 y NaCl (Cuadro 4).
Número de hojas. En criollo, el análisis entre tratamientos fue estadísticamente diferente (P<0.05) y en 'Hass' fue (P<0.01) (Cuadro 2). El criollo formó más hojas por plántula en el conjunto de tratamientos con CaCl2 en relación al testigo, a NaCl:CaCl2 y a NaCl (Cuadro 5). En cuanto a 'Hass', la mayor formación de hojas se dio en la combinación de NaCl:CaCl2; sin embargo, la formación de hojas fue mayor en el testigo con respecto a los tratamientos con sales.
Supervivencia. El análisis entre tratamientos mostró una diferencia significativa en la supervivencia de ambos materiales de aguacate (Cuadro 2). La supervivencia de criollo y 'Hass' fue estadísticamente igual tanto en el testigo como en algunos tratamientos con NaCl, CaCl2 y NaCl:CaCl2 (Cuadro 5). En los dos materiales vegetales, la supervivencia fue mayor (Cuadro 5) que la germinación y emergencia (Cuadro 3). En promedio, criollo superó la supervivencia de 'Hass' en aproximadamente 10 %.
La prueba de t para la comparación de medias de criollo y 'Hass' indicó que el criollo fue estadísticamente superior (P<0.01) en la mayoría de las variables evaluadas excepto en días a inicio de germinación, días a inicio de emergencia y porcentaje de supervivencia (Cuadro 2).
DISCUSIÓN
El estrés por sales es un factor abiótico que ocasiona cambios en el crecimiento y morfología de las plantas (Bernstein et al., 2001). Una forma de comparar la tolerancia entre materiales vegetales es evaluar la germinación y la respuesta de crecimiento ante condiciones de estrés (Pire et al., 2007) y en el cultivo in vitro, el eje embrionario de aguacate es una opción para seleccionar portainjertos tolerantes a salinidad en corto tiempo (González–Rosas et al., 2003; Fuentes et al., 2004). En este estudio, los materiales de aguacate criollo y 'Hass' variaron su respuesta a las condiciones salinas inducidas con NaCl, CaCl2 y NaCl:CaCl2. Según Hadas (1977), el proceso de germinación se inhibe desde el momento que la semilla no es capaz de adquirir agua por el proceso físico de imbibición, ocasionando retardo en la activación metabólica necesaria para la emisión radical y la posterior movilización de las reservas de las diferentes partes de la semilla, útiles para el crecimiento del eje embrionario (Jamil et al., 2006) y por la absorción excesiva de iones tóxicos, específicamente Na+ y Cl– (Prisco y O'Leary, 1970). En este estudio, tanto los días a inicio de germinación y emergencia como el porcentaje de germinación y emergencia de criollo y 'Hass', fueron marcadamente diferentes entre tratamientos. La germinación y emergencia en los tratamientos con NaCl como único componente en el medio de cultivo, fue más perjudicial que los compuestos con CaCl2 y NaCl:CaCl2, específicamente en 'Hass'. El NaCl, no sólo inhibe la germinación debido a la sequía fisiológica ocasionada por la disminución del potencial hídrico en el medio, sino también al aumento de la concentración de iones tóxicos en el embrión (Prisco y O'Leary, 1970).
En los dos materiales de aguacate se observó que no todos los ejes embrionarios que germinaron lograron la emergencia y de igual forma no todos los que emergieron consiguieron la emisión radical, por lo que se puede considerar que no existió relación positiva entre la germinación y la emergencia de las plántulas. La germinación fue más sensible a la salinidad que la emergencia de las plántulas; procesos que no fueron uniformes, al existir ejes embrionarios que adelantaron o retrasaron tanto la emisión de la raíz como de la plúmula; se cree que los factores que pueden marcar esta diferencia son el grado de madurez de los embriones o si los frutos son producto de la primera o segunda floración (variable no considerada en este estudio); no obstante, se comprueba que desde el inicio de su desarrollo las plantas muestran sensibilidad a la salinidad (Chang, 1961) y los materiales de aguacate criollo y 'Hass' no son la excepción. Por su parte, Norlyn y Epstein (1984) opinan que la capacidad de las semillas para germinar y posteriormente emerger en condiciones de estrés salino puede ser indicador del potencial genético que contienen para tolerarlo.
A los 65 días de cultivo, la longitud de raíz y tallo de las plántulas obtenidas de aguacate criollo y 'Hass' fueron mayores en el tratamiento 1. En condiciones salinas, la primer respuesta al estrés es la inhibición de crecimiento del tallo y las raíces (Lazof y Bernstein, 1998); en aguacate, las raíces son más sensibles al estrés por sales que los tallos (Bernstein et al., 2004). La formación y longitud de estos órganos se redujeron en presencia de NaCl, debido a su efecto tóxico, originando la absorción desbalanceada de nutrimentos, posiblemente porque el sistema radical pierde habilidad para controlar la entrada de iones y la capacidad de absorción de agua, factores que son cruciales para la supervivencia de las plantas (Hajibagheri et al., 1989;). La tolerancia de las plantas a las sales está en función de la habilidad del sistema radical para excluir o limitar el transporte de iones tóxicos como Na+ y Cl– a los tallos (Renault et al., 2001; Singh et al., 2003) y restablecer su balance hídrico para que su crecimiento prosiga (Maurel y Chrispeels, 2001).
Durante la formación y expansión de la hoja, en condiciones salinas, se encontró que el criollo desarrolló hojas con lámina totalmente expandida, algunas de ellas con la punta necrosada, síntoma que puede ser debido al exceso de Cl– más que de Na+ en el medio de desarrollo (Salazar–García et al., 1987). En 'Hass', el crecimiento de yemas foliares fue escaso y apenas visible; en consecuencia, la formación de hojas se redujo y la expansión foliar fue mínima. A pesar de que en aguacate la raíz es el órgano inmediato que sufre la salinidad más que las hojas (Bernstein et al., 2004), el primer síntoma visible es la reducción de la expansión de la hoja en la mayoría de las plantas (Parida y Das, 2005) como una reacción ante el estrés para conservar energía (Jaleel et al., 2008) o a la inhibición de crecimiento por la acumulación de sales (Prisco y O'Leary, 1970).
Los efectos de la salinidad en el desarrollo de estos materiales varió dependiendo del tipo y concentración de las sales, como lo mencionan Singh et al. (2003) y Ye et al. (2005). El crecimiento fue detenido más por NaCl que por CaCl2; con NaCkCaCl2 fue notorio que el Na+ y Cl–compitieron con Ca2+ restringiendo su efecto de protección; sin embargo, ni la germinación, la emergencia y ni el crecimiento fueron tan limitados en comparación al efecto de NaCl, por lo que, se considera que la presencia de Ca+2 inhibió el efecto dañino del NaCl al favorecer el crecimiento tanto de raíz como de tallo en ambos materiales. De acuerdo con Parida y Das (2005), estos resultados pueden indicar que en aguacate el desarrollo de los ejes embrionarios es afectado cuando son expuestos a salinidad y especialmente a NaCl, puesto que el crecimiento se detuvo y la producción de plántulas se redujo (Lazof y Bernstein, 1998), además se confirma que el crecimiento de raíces se limita más que el del tallo como lo manifiestan Salazar–García et al. (1987) y Bernstein et al. (2004).
El desarrollo escaso de las plantas que crecieron en estas condiciones, y aun cuando la germinación fue cercana al 60 % en criollo y 39 % en 'Hass', la supervivencia fue mayor a los 65 días de cultivo y diferente entre los tratamientos en los dos materiales (79 % en criollo y 69 % en 'Hass'). La diferencia entre estos procesos pudo deberse a que algunos ejes embrionarios germinaron, emergieron y permanecieron vivos, pero su crecimiento se detuvo; la de otros, a que únicamente acrecentaron su volumen sin mostrar síntomas de oxidación ni deshidratación. La supervivencia fue afectada negativamente por la oxidación y deshidratación tanto de ejes embrionarios como de plántulas. La oxidación y deshidratación se consideran respuestas asociadas a la concentración alta de sales en el medio y debido a esta condición se producen efectos hiperiónicos e hiperosmóticos (Hasegawa et al., 2000) que inducen la disminución del potencial hídrico e incrementan tanto la síntesis de compuestos fenólicos como de especies reactivas al oxígeno (Ayaz et al., 2000; Jaleel et al., 2007).
La tolerancia a sales no es un carácter constante en las plantas, sino que varía con el ambiente, estado de desarrollo (Zekri, 1994) y origen de la planta (Delesalle y Blum, 1994). La tolerancia durante la germinación, emergencia de la plántula y las etapas de crecimiento pueden ser afectadas de manera diferente (Zekri, 1994). Se puede asumir que la respuesta de criollo y 'Hass' sometidos a estrés salino, pudo deberse a las características intrínsecas de cada material de aguacate o de cada eje embrionario. En 'Hass', la germinación y emergencia fueron más rápidos, pero presentaron mayor afectación en condiciones salinas, lo que permite suponer que en una plantación comercial el nivel de tolerancia es dependiente del portainjerto utilizado (Salazar–García et al., 1987; Mickelbart et al., 2007).
La reducción y retraso en la germinación y emergencia, longitud de raíz y brotes, así como la supervivencia, pudieron deberse a la presencia de NaCl, tal como lo manifiestan Jamil et al. (2006) en remolacha, coliflor y amaranto, y Delesalle y Blum (1994) en los resultados obtenidos en ocho familias de Sagitaria latifolia. Adicionalmente, la disminución de la germinación y el retraso de la emergencia en los tratamientos con NaCl puede ser la evidencia de su efecto tóxico, ya que al adicionar Ca2+ se incrementó la germinación y emergencia. Esto concuerda también con los resultados señalados en cítricos por Zekri (1994).
En este estudio, la tendencia general de efecto negativo ocasionado por la salinidad en la germinación, emergencia, desarrollo y supervivencia de aguacate criollo y 'Hass' fue: NaCl > NaCkCaCl2 CaCl2 Testigo.
CONCLUSIONES
El aguacate criollo fue más tolerante al estrés por sales con respecto a 'Hass'. La adición del NaCl al medio de cultivo MS (1962) al 100 % de su composición mineral, en comparación al CaCl2, tuvo un efecto negativo mayor en los procesos de germinación y emergencia de los ejes embrionarios, el desarrollo y supervivencia de las plántulas especialmente de 'Hass'. El cultivo in vitro de ejes embrionarios maduros de aguacate permitió determinar que es posible identificar materiales tolerantes y sensibles al estrés salino a los 65 días después de su establecimiento.
BIBLIOGRAFÍA
ANÓNIMO. 2010. Anuario estadístico de la producción agrícola. Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación (SAGARPA). Sistema de Información Agroalimentaria y Pesquera (SIAP). Situación Nacional hasta abril de 2010. En www.siap.gob.mx. Consultado en mayo de 2010. [ Links ]
AYAZ, F. A.; KADIOGLU, A.; TURGUT, R. 2000. Water stress effects on the content of low molecular weight carbohydrates and phenolic acids in Ctenanthe setosa (Rose) Eichler. Canadian Journal Plant Science 80:373–378. [ Links ]
BEN–YA'ACOV, A. D.; ZILBERSTAINE, A. M. 1999. The possible use of avocado (Persea americana Mill) germoplasm material as rootstocks for soil stress conditions. Revista Chapingo Serie Horticultura 5:25–28. [ Links ]
BERNSTEIN, N.; IOFFE, M.; ZILBERSTAINE, M. 2001. Salt–stress effects on avocado rootstock growth I. Establishing criteria for determination of shoot growth sensitivity to the stress. Plant and Soil 233:1–11. [ Links ]
BERNSTEIN, N., MEIRI, M.; ZILBERSTAINE, M. 2004. Root growth of avocado is more sensitive to salinity than shoot growth. Journal of the American Society for Horticultural Science 129:188–192. [ Links ]
CASTRO, V. M.; ITURRIETA, E. R.; FASSIO, O. C. 2009. Rootstock effect on the tolerance of avocado plants cv. Hass to NaCl stress. Chilean Journal of Agricultural Research 69:316–324. [ Links ]
CHANG, C.W. 1961. Effects of saline irrigation water and exchangeable sodium on soil properties and growth of alfalfa. Soil Science 91:29–37. [ Links ]
DELESALLE, V. A.; BLUM, S. 1994. Variation germination and survival among families of Sagitaria latifolia in response to salinity and temperature. International Journal of Plant Sciences 155:187–195. [ Links ]
FUENTES,J. L.; RODRIGUEZ, N. N.; SANTIAGO, L.; VALDÉS, Y.; RAMIREZ, I. M.; RODRIGUEZ, J. A. 2004. Zygotic embryo culture in avocado (Persea americana Mill). Cultivos Tropicales 25:73–76. [ Links ]
GAZIT, S.; KADMAN, A. 1976. Growing avocados in areas of high salinity. Proceedings of the first International Tropical Fruit Short Course: The avocado. Institute of Food and Agricultural Sciences, University of Florida 58–60 [ Links ]
GONZÁLEZ–ROSAS, H; SALAZAR–GARCÍA, S.; RAMÍREZ–REYES, G.; RODRÍGUEZ–ONTIVEROS, J. L.; RAMOS–VILLASEÑOR, A. C. 2003. Preliminary results on in vitro selection for tolerant to chloride excess in avocado. Revista Chapingo Serie Horticultura 9:39–43. [ Links ]
HADAS, A. 1977. Water uptake and germination of leguminous seeds in soils of changing matrix and osmotic water potential. Journal Experimental Botanic 28: 977–985. [ Links ]
HAJIBAGHERI, M. A.; YEO, A. R.; FLOWER, T. J.; COLLINS, J. C. 1989. Salinity resistance in Zea mays fluxes of potassium, sodium an chloride, cytoplasmic concentrations and mi–crosomal membrane lipids. Plant, Cell and Environment 12: 753–757. [ Links ]
HASEGAWA, P. M.; BRESSAN, R. A.; ZHU, J. K.; BOHNERT, H. J. 2000. Plant cellular and molecular responses to high salinity. Annual Review Plant Physiology and Plant Molecular Biology 51:463–499. [ Links ]
JALEEL, CH. A.; GOPI, R.; MANIVANNAN, P.; PANNEERSELVAM, R. 2007. Antioxidative potentials as a protective mechanism in Catharanthus roseus (L.) G. Don. Plants under salinity stress. Turk Journal Botanic 31:245–251. [ Links ]
JALEEL, CH. A.; SANKAR, B.; SRIDHARAN, R.; RANNEERSELVAM, R. 2008. Soil salinity alters growth, chlorophyll content, and secondary metabolite accumulation in Catharanthus roseus. Turk Journal Biology 32:1–5. [ Links ]
JAMIL, M., LEE, D. B.; JUNG, K. Y.; MUHAMMAD, A.; LEE, S. CH.; RHA, E. S. 2006. Effect of salt (NaCl) stress on germination and early seedling growth of four vegetables species. Journal Central European Agriculture 7(2):273–282. [ Links ]
LAZOF, D. B.; BERNSTEIN, N. 1998. The NaCl–induced inhibition of shoot growth: The case for disturbed nutrition with special consideration of calcium nutrition. Advances in Botanical Research 29:115–190. [ Links ]
MAUREL, C.; CHRISPEELS, M.J. 2001. Aquaporins. A molecular entry into plant water relations. Plant Physiology 125:135–138. [ Links ]
MICKELBART, M. V.; ARPAIA, M. L. 2002. Rooststock influences in ion concentration, growth, and photosynthesis of 'Hass' avocado trees in response to salinity. Journal of the American Society for Horticultural Science 127:649–655. [ Links ]
MICKELBART, M.V.; MELSER, S.; ARPAIA, M.L. 2007. Salinity–induced changes in ion concentration of 'Hass' avocado trees on three rootstocks. Journal of Plant Nutrition 30:105–122. [ Links ]
MURASHIGE, T.; SKOOG, F. 1962. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiology Plant. 15:473–497. [ Links ]
MYERS, B.A.; MORGAN, W.C. 1989. Germination of the salt–tolerant grass Diplachne fusca. II. Salinity responses. Australian Journal of Botanic 37:239–251. [ Links ]
NORLYN, J.D.; EPSTEIN, E.. 1984. Variability in salt tolerance of four Triticale lines at germination and emergence. Crop Science 24:1090–1092. [ Links ]
PARIDA, A.K.; DAS, A.B. 2005. Salt tolerance and salinity effects on plants: a review. Ecotoxicology Environmental Safety. 60:324–349. [ Links ]
PIRE, R.; PEREIRA, A.; DIEZ, J.; FERERES, E. 2007. Evaluación de la tolerancia a la sequía de un portainjerto venezolano de vid y posibles mecanismos condicionantes. Agrociencia 41:435–446. [ Links ]
PRISCO, J.T.; O'LEARY, J.W. 1970. Osmotic and "toxic" effects of salinity on germination of Phaseolus vulgaris L. seeds. Turrialba 20:177–184. [ Links ]
RENAULT, S.; CROSER, C.; FRANKLIN, A.J.; ZWIAZEK, J.J. 2001. Effects of NaCl and Na2So4 on red–osier dogwood (Cornus stolonifera Michx) seedlings. Plant and Soil 233:261–268. [ Links ]
SALAZAR–GARCÍA, S.; CORTÉS–FLORES, J.I.; ALCALDE–BLANCO, S.; ZÁRATE–DE LARA, G.P. 1987. Daños por salinidad en árboles de aguacate fuerte en Atlixco, Puebla. Agrociencia 68:115–134. [ Links ]
SALAZAR–GARCÍA, S.; LAZCANO–FERRAT, I. 1999. Diagnóstico nutrimental del aguacate 'Hass' bajo condiciones de temporal. Revista Chapingo. Serie Horticultura 5:173–184. [ Links ]
SINGH, K.N.; CHATRATH, R. 2001. Salinity tolerance. Pp 101–110. In: Application of physiology in wheat breeding. REYNOLDS, M.P., ORTIZ–MONASTERIO, J.I., McNAB, A. (eds.). CIMMYT. México, D.F. [ Links ]
SINGH A.; SAINI, M.L.; BEHL, R.K. 2003. Screening of citrus rootstocks for salt tolerance in semi–arid climates–a review. Topics. 13:53–66. [ Links ]
YE, Y.; FUNG–YEE, T.; CHANG–YI, L.; YUK–SHAN, W. 2005. Effects of salinity on germination, seedlings growth and physiology of three salt–secreting mangrove species Aquatic Botanic 83:193–205. [ Links ]
ZHANG, Y. 1994. Development of in vitro bioassays for determination of salinity tolerance in potato (Solanum spp). Ph.D. Dissertation, Department of Plant Science. McGill University. Montreal, Canada. 138 p [ Links ]
ZEKRI, M. 1994. Effects of salinity and calcium on seedling emergence, growth and sodium and chloride concentrations of citrus rootstocks. Proceeding of Florida State Horticulture Society 106:18–21. [ Links ]

