










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista Chapingo. Serie horticultura
versão On-line ISSN 2007-4034versão impressa ISSN 1027-152X
Rev. Chapingo Ser.Hortic vol.16 no.2 Chapingo Mai./Ago. 2010
Relaciones injerto–interinjerto y características anatómicas y fisiológicas de la hoja de cuatro genotipos de aguacate
Scion–interstock relationships and anatomical and physiological leaf characteristics of four avocado genotypes
J. Ayala–Arreola1*; A. F. Barrientos–Priego1; M. T. Colinas–León1; J. Sahagún–Castellanos1; J. C. Reyes–Alemán2
1 Posgrado en Horticultura, Departamento de Fitotecnia, Universidad Autónoma Chapingo. km 38.5 carretera México–Texcoco, Chapingo, Estado de México. C. P. 56230, MÉXICO. Correo–e: ayala67755@hotmail.com (*Autor responsable).
2 Fundación Salvador Sánchez Colín, CICTAMEX, S. C. Ignacio Zaragoza Núm. 6 Coatepec Harinas Estado de México, C. P. 51700. MÉXICO.
Recibido: 8 de abril, 2009.
Aceptado: 23 de junio, 2010.
Resumen
Se evaluó la influencia del interinjerto en las características fisiológicas y anatómicas de la hoja de aguacate 'Hass', 'Fuerte', 'Colín V–33' y 'Duke 7' utilizados como interinjertos e injertos finales. En la lámina foliar se determinó la densidad estomática, densidad de células epidérmicas, índice estomático y diámetro polar del estoma. En cortes transversales de la vena central y del pecíolo de la hoja se evaluó: área, perímetro, longitud del eje mayor y menor, índice de redondez y diámetro Feret en vasos del xilema. En cortes transversales de hoja, se evaluó: el grosor de la epidermis superior e inferior y del parénquima en empalizada I. Las variables fisiológicas evaluadas fueron: tasa de asimilación de CO2, conductancia estomática, concentración interna de CO2 resistencia estomática, tasa de transpiración y temperatura de la hoja. 'Colín V–33' como interinjerto promovió la mayor densidad estomática, mayor densidad de células epidérmicas y grosor de la capa de parénquima en empalizada I. En la comparación intra genotipos se encontró que 'Hass' y 'Fuerte' con interinjerto incrementaron en 18.8 y 17.8 % el número de estomas, mientras que en 'Duke 7' disminuyó en un 25.4 %. La densidad de células epidérmicas se incrementó de manera significativa en 'Fuerte' (23.3 %) con la presencia del interinjerto. 'Fuerte' y 'Duke 7' con interinjerto incrementaron significativamente el grosor de la epidermis inferior en 14.9 y 12.1 %, respectivamente, mientras que 'Duke 7', 'Hass' y 'Colín V–33' con interinjerto disminuyeron el grosor de epidermis inferior en 19.5 y 1.0 %. Se encontró que 'Hass' como interinjerto promovió la tasa de asimilación neta de CO2 más alta, siendo diferente 'Duke 7'. Para resistencia estomática le correspondió el menor valor a 'Colín V–33' y presentó diferencias con 'Duke 7'.
Palabras clave: Asimilación de CO2, resistencia estomática, Persea americana, densidad de estomas, sistema vascular.
Abstract
The influence of interstock in anatomical and physiological leaf characteristics were evaluated in 'Hass', 'Fuerte', 'Colín V–33' and 'Duke 7' used as interstocks and final grafts. Stomatal density, epidermal cell density, stomatic index and stomata polar diameter were assessed in leaf blade. In cross sections of the central vein and petiole of the leaf: area, perimeter, length of the minimum and maximum axis, roundness index and Feret diameter of xylem vessels were determined. Thickness of the upper and lower epidermis and thickness of palisade parenchyma I layer were measured in leaf cross sections. Physiological variables were CO2 assimilation rate, stomatic conductance, CO2 internal concentration, stomatic resistance, transpiration rate and leaf temperature. 'Colín V–33' as interstock induced higher stomatal and epidermal cell density and thicker palisade parenchyma I. In the intra genotype comparison, 'Hass' and 'Fuerte' with interstock increased stomatal density 18.8 and 17.8 %, respectively, while 'Duke 7' caused a 25.4 % reduction. Epidermal cell density increased in 'Fuerte' (23.3 %) with the presence of the interstock. 'Fuerte' and 'Duke 7' with interstock increased the thickness of the lower epidermis in 14.9 and 12.1 %, respectively, while 'Hass' and 'Colín V–33' with interstock reduced epidermis thickness by 19.5 and 1.0 %, respectively. 'Hass' as interstock promoted a higher CO2 assimilation rate than that promoted by 'Duke 7'. 'Colín V–33', as a rootstock, showed lower stomatic resistance than 'Duke 7'.
Key words: CO2 assimilation, stomatal resistance, Persea americana, stomatal density, vascular system.
INTRODUCCIÓN
El cultivo del aguacate en México comprende una superficie de 122,186 ha en producción, las cuales generan 1,120 000 t de fruta (Anónimo, 2009). Este cultivo es un producto de alto valor nutritivo y de importancia tradicional en la dieta mexicana, así como para el mercado de exportación y es una fuente de ingreso de divisas para el país (Santacruz, 2004). La principal forma de propagación del aguacate es la injertación de cultivares comerciales sobre portainjertos francos (originados por semilla), mismos que presentan una alta heterocigosis que resulta en huertos heterogéneos en cuanto a comportamiento y productividad. Por tal motivo, reviste vital importancia la elección de un buen portainjerto, ya que de él puede depender el éxito o fracaso de una plantación (Barrientos–Priego et al., 2000).
La investigación y selección de posibles portainjertos comerciales se ha ampliado de la resistencia a Phytophtho–ra cinnamomi Rands para incluir a otras enfermedades, así como a la tolerancia a la salinidad, sequía, pH alcalino, mayor productividad y precocidad en la producción, calidad, tamaño de fruto y un efecto enanizante. Las nuevas características adquiridas en la planta son el resultado de una intensa interacción entre el portainjerto o interinjerto y el injerto, aportando diferente información genética cada uno (Lockard y Schneider, 1981; Lacono et al., 1998). Sin embargo, el uso de interinjertos en la producción de planta y en plantaciones comerciales de aguacate, como una alternativa para solucionar los problemas anteriormente mencionados, ha sido poco estudiado (Ben–Ya'acov y Michelson, 1995). Debido a la poca información sobre las relaciones injerto/interinjerto, esta investigación planteó como objetivo el evaluar si existen modificaciones por influencia del interinjerto en las características fisiológicas y anatómicas de la hoja del injerto Anal en cuatro genotipos de aguacate.
MATERIALES Y MÉTODOS
El experimento se realizó en un invernadero ubicado en el Campo Agrícola Experimental San Martín, del Departamento de Fitotecnia de la Universidad Autónoma Chapingo, en Chapingo, Estado de México, situado a 19° 29' N y a 98° 53' O, a una altitud de 2,240 m.
Material vegetal
Las varetas que se utilizaron para generar los interin–jertos y los injertos finales se obtuvieron de árboles en producción establecidos en la Fundación Salvador Sánchez Colín CICTAMEX, S. C., en Coatepec Harinas, Estado de México, en México. Como interinjertos e injertos Anales utilizaron 'Hass', 'Fuerte', 'Colín V–33' y 'Duke 7'. Como portainjertos se usaron plantas de la raza Guatemalteca, los cuales fueron obtenidos a partir de semilla. La injertación para generar los interinjertos se llevó a cabo de junio a diciembre de 2006 y la injertación sobre los interinjertos se realizó de marzo a mayo de 2007. El tipo de injerto utilizado fue de hendidura.
Diseño experimental y diseño de tratamientos
El diseño experimental utilizado fue un completamente al azar con un arreglo de tratamientos factorial (4x4), en donde el factor 1 fue el interinjerto ('Hass', 'Fuerte', 'Colín V–33' y 'Duke 7') y el factor 2 fue el injerto final ('Hass', 'Fuerte', 'Colín V–33' y 'Duke 7'). Ambos factores contaron con cuatro niveles (Los cultivares anteriormente mencionados). La unidad experimental consistió en una planta y se tuvieron cuatro repeticiones de cada tratamiento; adicionalmente, se consideró un testigo sin injertar para cada cultivar, con cuatro repeticiones, utilizando un total de 80 unidades experimentales (Cuadro 1).

Variables de estomas y células epidérmicas
En cada uno de los injertos finales se seleccionó y se etiquetó una hoja madura y completamente expandida; de ésta se obtuvo una impresión con silicón para huellas dentales (Exactodent®). De la impresión se obtuvo, con barniz de uñas, la impresión positiva y después se fijó en portaobjeto y cubreobjeto. En la impresión se determinó la densidad estomática, densidad de células epidérmicas, índice estomático y diámetro polar de estoma (10 estomas por campo). El índice estomático se calculó a partir de la siguiente expresión: IE = [DE / (DE + DCE)] x 100; donde IE = índice estomático, DE = densidad estomática, y DCE = densidad de células epidérmicas.
Los caracteres de estomas fueron evaluados en cinco campos (40X de objetivo y 10X de ocular), con un microscopio Motic B3 Professional Series, con adaptación de una cámara de alta resolución Moticam 480, con un adaptador de 12 mm. Se utilizó el programa Imagen Plus 2000 para darle formato digital.
Cortes anatómicos en hoja
Las hojas de donde se extrajeron las réplicas para la evaluación de las variables estomáticas se colectaron y se fijaron en una solución FAA (50 % etanol 96° + 5 % ácido acético glacial + 10 % de formaldehído con 40 % de pureza + 35 % agua destilada). Después, se procesaron en un cambiador automático de tejidos (Tissuematon Fisher) con cellosolve y xileno, para transferirse a parafina (55 °C), posteriormente permanecieron 72 horas dentro de una estufa.
Se elaboró el taquete y pirámide de parafina de acuerdo a la metodología propuesta por Sass (1968). En un micrótomo rotatorio (American Optical Modelo 820) se realizaron cortes transversales en la hoja abarcando la vena central con un grosor de 10 |jm, los cuales se montaron con adhesivo Haup y formol al 10 % y su tinción se hizo con safranina y verde fijo. Se evaluaron las variables: área, perímetro, longitud del eje mayor y menor, índice de redondez (4 x PI x área) e índice Feret [(4.área/n)0,5], todo en vasos del xilema a 40X de objetivo y 10X de ocular. Las mediciones fueron cuantificadas en cinco campos por planta, con base en el estudio de Sperry y Saliendra (1994).
En cortes de la lámina foliar se evaluó: el grosor de la epidermis superior e inferior y grosor de parénquima en empalizada I. Los cortes transversales en pecíolos se realizaron a 10 |m mediante un micrótomo de congelamiento de la marca Heindelberg modelo 880 (American Optical Corp). Se evaluaron las variables: área, perímetro, longitud del eje mayor y menor, índice de redondez e índice Feret, todo en vasos del xilema a 40X de objetivo y 10X de ocular. La medición de las variables fue a partir de imágenes digitales, con el programa Image Tool versión. 3.00 (Wilcox et al., 2002).
Variables fisiológicas
Antes de realizar los cortes anatómicos en la hoja seleccionada y previamente identificada mediante una etiqueta en cada uno de los injertos finales y en las plantas testigos, se evaluaron mediante un analizador de gases infrarrojo (IRGA; modelo LI–COR 6200) las variables tasa de asimilación de CO2, conductancia estomática, concentración interna de CO2. Las variables resistencia estomática, tasa de transpiración y temperatura de la hoja se evaluaron con un porómetro (modelo LI–COR 1600). Se realizaron tres mediciones semanales, entre las 9:30 a 14:30 horas, antes del corte de la hoja.
Análisis estadístico
Se realizó un análisis de varianza por factor (inte–rinjerto e injerto final) y se llevaron a cabo las pruebas de comparación de medias (Tukey P<0.05). También se realizaron contrastes ortogonales para el factor injerto final y así comparar con el testigo sin interinjerto. Además, se realizó un análisis de correlación de Pearson.
RESULTADOS
Factor interinjerto
La prueba de comparación de medias para el factor interinjerto (Cuadro 2) mostró que 'Colín V–33' promovió la mayor densidad estomática, al igual que mayor densidad de células epidérmicas siendo diferente a los demás interinjertos para ambas variables (P<0.05). Sin embargo, no existió diferencia entre cultivares para el índice estomático. Para diámetro del estoma, 'Fuerte' indujo mayor valor y 'Hass' menor valor (P<0.05).
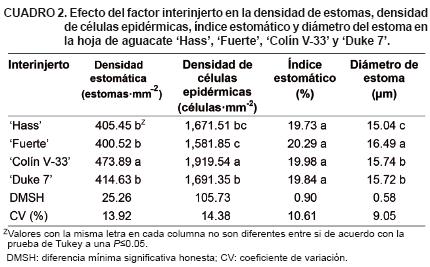
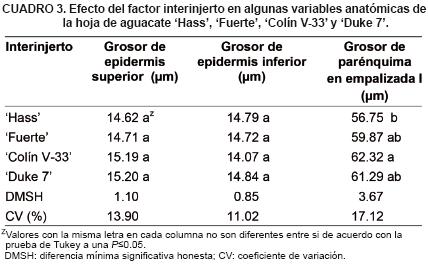
Para el grosor del parénquima en empalizada I se observaron diferencias (P<0.05) entre 'Colín V–33' y 'Hass' (Cuadro 3). No se encontraron diferencias (P<0.05) para los grosores de la epidermis superior e inferior entre interinjertos. El grosor de parénquima en empalizada presentó una correlación positiva y significativa con la mayoría de las variables anatómicas del vaso del xilema tanto en la vena central como en el pecíolo de la hoja (valores no mostrados).
En la tasa de asimilación neta se observaron diferencias (P<0.05) entre 'Hass' y 'Fuerte' con 'Duke 7'. En tanto que para resistencia estomática hubo diferencias entre 'Colín V–33' y 'Duke 7' (Cuadro 4). En los vasos del xilema de la vena central de la hoja 'Duke 7' provocó mayores valores que 'Hass' para área, perímetro, longitud del eje mayor, longitud del eje menor e índice Feret. En tanto que, para estas mismas variables en el pecíolo de la hoja se encontró que 'Fuerte', como interinjerto, provocó mayores valores que 'Colín V–33' (Cuadro 5).

Factor injerto final
El injerto final de 'Duke 7', con interinjerto, presentó la mayor densidad de estomas, mayor densidad de células epidérmicas y el menor diámetro del estoma, mostrando diferencias con 'Hass', 'Fuerte' y 'Colin V–33' (Cuadro 6). 'Duke 7' sin interinjerto tuvo los mayores valores para la densidad estomática y para la densidad de células epidérmicas siendo diferente (P<0.05) a los demás cultivares.
En la comparación intra cultivares mediante contrastes ortogonales se encontró que la presencia del interinjerto en 'Hass' y 'Fuerte' incrementó significativamente (18.8 y 17.8 %, respectivamente) la densidad de estomas. La densidad de células epidérmicas se incrementó significativamente en 23.3 % con la presencia del interinjerto en 'Fuerte'. En 'Hass', con interinjerto, el índice estomático se incrementó significativamente en 7.3 %. El diámetro de estoma fue mayor en 'Hass', 'Fuerte' y 'Duke 7' sin interinjertos.
El grosor de epidermis superior fue mayor en 'Hass' y Colín 'V–33', con interinjerto, presentando diferencias con 'Fuerte' y 'Duke 7'. En tanto que, para el grosor de epidermis inferior 'Duke 7' con interinjerto incrementó significativamente el grosor y fue diferente a 'Hass', 'Fuerte' y 'Colín V–33'. En cuanto al comportamiento de los injertos finales, sin interinjerto se observaron diferencias (P<0.05) para el grosor de epidermis superior e inferior entre 'Hass' y 'Fuerte' (Cuadro 7). Para el grosor de parénquima en empalizada I 'Colin V–33' sin interinjerto tuvo el mayor valor y fue diferente (P<0.05) a los demás cultivares. En la comparación intra cultivares se encontró que 'Duke 7' con interinjerto disminuyó 9.2 % el grosor de la epidermis superior y fue diferente a cuando se estableció sin interinjerto. 'Fuerte' y 'Duke 7', con interinjerto, incrementaron el grosor de la epidermis inferior en 14.9 y 12.1 %, respectivamente siendo diferentes al que no tenía interinjerto.
La prueba de comparación de medias para el factor injerto final con interinjerto mostró que 'Hass' 'Fuerte' y 'Colín V–33' presentaron la mayor tasa de asimilación neta de CO2, siendo diferente a 'Duke 7'; este último presentó la mayor tasa de asimilación neta de CO2 pero sin interinjerto, siendo diferente a 'Fuerte'. Con respecto a la tasa de transpiración y conductancia estomática no se encontraron diferencias entre genotipos (Cuadro 8). Para resistencia estomática 'Hass', sin interinjerto, presentó el mayor valor y 'Colín V–33' el menor.
Mediante contrastes ortogonales se encontró que 'Fuerte', con interinjerto, incrementó significativamente en 76.7 % la tasa de asimilación neta comparado con el que no tenía interinjerto (Cuadro 8). La resistencia estomática se vio disminuida significativamente (P<0.05) con el interinjerto para 'Hass' y 'Fuerte' en 38.46 y 26.31 %, respectivamente. Para las variables concentración interna de CO2 y temperatura de la hoja no hubo diferencias significativas intra cultivares (Cuadro 9).

En las características anatómicas de vasos de xilema en la vena central de la hoja para el factor injerto final con interinjerto se encontró diferencias (P<0.05) para el área, perímetro, longitud del eje mayor, longitud del eje menor e índice Feret, de 'Duke 7' con el resto de los cultivares (Cuadro 10). Cuando se evaluó en injerto final, sin interinjerto, se observó que la vena central de la hoja de 'Colín V–33' fue diferente (P<0.05) a los demás genotipos en: área, perímetro, longitud del eje mayor y menor, índice Feret (Cuadros 10 y 11).

En las características anatómicas de las células de xilema en el pecíolo de la hoja para el factor injerto final con interinjerto se encontró que 'Colín V–33' y 'Hass' presentaron diferencias (P<0.05) con 'Duke 7' en la mayoría de las variables anatómicas evaluadas, excepto para la variable índice de redondez, donde 'Duke 7' presentó el mayor valor (Cuadros 10 y 11). 'Colín V–33' sin interinjerto mostró los mayores valores para el área, perímetro, longitud del eje mayor y menor e índice Feret, siendo diferente (P<0.05) a todos los demás cultivares (Cuadros 10 y 11).
DISCUSIÓN
Para 'Fuerte' como injerto final, la densidad estomática encontrada coincide con el intervalo reportado por Blanke (1992). Sin embargo, en general los datos no coinciden con los reportados por Macías y Borys (1980). Esto, debido al efecto de las condiciones ambientales prevalecientes en el invernadero y a que estos autores estudiaron plantas francas de aguacate de diversas procedencias.
La presencia del interinjerto en 'Hass', 'Fuerte' y 'Colín V–33' tendió a incrementar la densidad de estomas y de células epidérmicas. Probablemente lo anterior se deba al efecto de injertación, coincidiendo parcialmente con lo encontrado por Páres et al. (2004) en anonas. Esto tal vez se pueda interpretar como un aspecto positivo ya que se esperaría un incremento en conductancia estomática, y en consecuencia se aumentaría la capacidad de transpiración. Sin embargo, no se encontró tal relación en el presente estudio, sólo reducción en la resistencia estomática, en algunos genotipos, lo cual puede ser de importancia a la adaptación en condiciones de estrés.
El alto grado de asociación entre el índice estomático y la densidad estomática reportada por Verdugo et al. (1999) no fue muy claro en el presente estudio. Esto puede ser que se deba a que el índice estomático depende de la densidad de estomas y células epidérmicas que pueden estar fuertemente influenciadas por las condiciones ambientales y por el genotipo que se trate (Wilkinson, 1979; Roth et al., 1986; Rubino et al., 1989; Bethke y Drew, 1992 y Salas et al., 2001). También, se puede atribuir a que tanto la frecuencia como el tamaño de los estomas pueden variar en función de la posición foliar, las condiciones de crecimiento y el cultivar (Sánchez–Díaz y Aguirreola, 2008).
'Colín V–33' como injerto Anal, con y sin interinjerto presentó el mayor grosor de parénquima en empalizada I y valores muy similares para el grosor de epidermis superior e inferior lo que podría indicar que es más eficiente desde el punto de vista fotosintético y que posiblemente tenga un comportamiento más constante (Wilson y Cooper, 1970 y Barden, 1978). En este genotipo fue donde se apreció más claramente la presencia de dos capas de parénquima en empalizada, lo cual parcialmente se reflejó en la asimilación de CO2 presentando valores altos al igual que 'Duke 7' que a su vez tenía un mayor contraste en densidad estomática.
En cuanto a las variables fisiológicas, tasa de asimilación neta de CO2, tasa de transpiración, conductancia estomática y temperatura de la hoja, se observó que con la presencia del interinjerto tendieron a incrementarse en 'Hass' y 'Fuerte', principalmente. Esto puede estar relacionado con lo encontrado en las dimensiones de los vasos del xilema del pecíolo y de la vena central de la hoja, donde se vieron favorecidas por la presencia del interinjerto, lo cual se relaciona con el transporte del agua en aguacate (Reyes et al., 2002), y esto al parecer podría haber influido en la eficiencia fisiológica de la hoja. La resistencia estomática disminuyó con la presencia del interinjerto en 'Hass', 'Fuerte' y 'Duke 7'. Para este último genotipo se tuvieron los menores valores en la mayoría de las variables fisiológicas evaluadas, con excepción de la concentración interna de CO2 y la temperatura de la hoja, esto quizá sea debido a que éste no respondió adecuadamente a las condiciones de invernadero ya que se manifestaron precozmente síntomas de deficiencia nutrimental y los brotes presentaron un bajo crecimiento, efecto más acentuado cuando se combinó con 'Colín V–33' como interinjerto. De hecho 'Duke 7' en campo también presenta lento crecimiento y es frecuente la deficiencia de nitrógeno (Alejandro F. Barrientos Priego, comunicación personal).
Experimentalmente en 'Colín V–33', como interinjerto, se ha encontrado que existe una disminución de la auxina sintetizada en la copa; esta misma auxina tiene flujo polar basipétalo a través del floema y células cambiales, donde el ABA tiene una función importante en el balance ABA/ AIA (Barrientos–Villaseñor et al., 1999), esta característica juega un papel importante en la regulación de la forma de la planta y también en la formación de los tejidos vasculares; por lo que se podría inferir que en 'Colín V–33', como interinjerto, repercute en una menor dimensión de la célula y en el efecto sobre la asimilación de CO2 y resistencia estomática, tal como se encontró en el presente estudio. Lo anterior hace suponer que la cantidad de auxina que llega a la raíz, es menor y en consecuencia se tiene poco desarrollo radical y por lo tanto, la cantidad de citocininas sintetizadas, que son translocadas por el xilema a los puntos de crecimiento, también es menor; esto implica necesariamente que la molécula de la auxina sufre una rápida e irreversible destrucción durante la estimulación del crecimiento (Lockard, 1981), lo cual trae indudablemente cambios anatómicos y como consecuencia respuestas fisiológicas contrastantes.
CONCLUSIONES
La presencia del interinjerto modificó de manera significativa algunas características anatómicas de elementos del vaso de la vena central y del pecíolo de la hoja, así como algunos aspectos del intercambio de gases de la hoja.
El genotipo 'Duke 7' usado como interinjerto influyó en la mayoría de las características anatómicas de elementos de vaso de la vena central y el pecíolo de la hoja, así como en la tasa de asimilación de CO2 y resistencia estomática.
En general 'Hass' y 'Fuerte' presentaron mayores modificaciones en la anatomía y fisiología por efecto del interinjerto.
LITERATURA CITADA
ANÓNIMO. 2009. Dirección General Adjunta de Planeación Estratégica y Análisis Sectorial, Financiera Rural. Monografía Aguacate. D.F., México. [ Links ]
BLANKE, M. M. 1992. Photosynthesis of avocado fruit. Proc. Second World Avocado Congress. Orange, California, USA. pp. 179–189. [ Links ]
BARRIENTOS–PRIEGO, A. F.; MUÑOZ–PÉREZ, R. B.; BORYS, M. W.; MARTÍNEZ–DAMIÁN, MA. T. 2000. Cultivares y portainjertos del aguacate, pp. 35–54. In: El Aguacate y su Manejo Integrado. TÉLIZ, D.; GONZÁLEZ, H.; RODRÍGUEZ, J.; DROMUNDO, R. (eds.). Mundi–Prensa México, S.A. de C.V. D.F., México. [ Links ]
BARRIENTOS–VILLASEÑOR, A.; BARRIENTOS–PRIEGO, A. F.; RODRÍGUEZ–PÉREZ, J. E.; PEÑA–LOMELI, A.; MUÑOZ–PÉREZ, R. 1999. Influencia del interinjerto cv. Colín V–33 sobre algunos aspectos fisiológicos en aguacatero (Persea americana Mill.). Revista Chapingo Serie Horticultura 5: 103–116. [ Links ]
BARDEN, J. A. 1978. Apple leaves, their morphology and photosynthetic potential. HortScience 13(6): 644–645. [ Links ]
BEN–YA'ACOV, A. D.; MICHELSON, E. 1995. Avocado rootstocks. Horticultural Reviews 17: 381–429. [ Links ]
BETHKE, C. P.; DREW, C. M. 1992. Stomatal and nonstomatal components to inhibition of photosynthesis in leaves of Capsicum annuum during progressive exposure to NaCl salinity. Plant Physiology 99(1): 219–226. [ Links ]
LACONO, F.; BUCCELLA, A.; PETERLUNGER, E. 1998. Water stress and rootstock influence on leaf gas exchange of grafted and ungrafted grapevines. Scientia Horticulturae 75(1): pp 27–39, [ Links ]
LOCKARD, R. G.; SCHNEIDER, G. W. 1981. Stock and scion relationships and the dwarfing mechanism in apple. Horticultural Reviews 3: 315–375. [ Links ]
MACÍAS G., J. L.; BORYS, M. W. 1980. Distribución de estomas en una población de Persea americana var. drymifolia Mill. Programa de Actividades y Resúmenes de Ponencias del Congreso Nacional de la Sociedad Mexicana de Fitogenética (SOMEFI). Uruapan, México. p. 82. [ Links ]
PÁRES–MARTÍNEZ, J.; ARIZALETA, M.; SANABRIA, M. E.; BRITO, L. 2004. Características de los estomas, densidad e índice estomático y su variación en función de la injertación en Annona muricata. L. y Annona montana MADFAC. Bioagro 16(3): 213–218. [ Links ]
REYES–SANTAMARÍA, I; TERRAZAS, T; BARRIENTOS–PRIEGO, A. F; TREJO, C. 2002. Xylem conductivity and vulnerability in cultivars and races of avocado. Scientia Horticulturae 92(2): 97–105. [ Links ]
ROTH, I.; MÉRIDA, T.; LINDORF, H. 1986. Morfología y anatomía foliar de plantas de la selva nublada de Rancho Grande, pp. 205–241. In: La Selva Nublada de Rancho Grande. Parque Nacional 'Henry Pittier". El Ambiente Físico, Ecología General y Anatomía Vegetal. JUBER, O. (ed). Fondo Editorial Acta Científica Venezolana. Caracas, Venezuela. [ Links ]
RUBINO, P.; TARANTINO, E.; REGA, F. 1989. Relationship between soil water status and stomatal resistance of tomatoes. Irrigazione e Drenaggio 36: 95– 98. [ Links ]
SALAS, J. A; SANABRIA, M. E; PIRE, R. 2001. Variación en el índice y densidad estomática en plantas de tomate (Lycopersicon esculentum Mill.) sometidas a tratamientos salinos. Bioagro 13(3): 99–104. [ Links ]
SÁNCHEZ–DÍAZ, M.; AGUIRREOLEA, J. 2008. Transpiración y movimientos estomáticos, pp. 41–56. In: Fundamentos de Fisiología Vegetal. AZCÓN–BIETO, J.; TALÓN, M. (eds). McGraw–Hill–Interamericana. Barcelona, España. [ Links ]
SANTACRUZ U., H. 2004. Actividad productora del aguacate en México y Michoacán. Tecnoagro 5(13): 13–18. [ Links ]
SASS, J. E. 1968. Botanical Microtechnique. 3rd. Edition. The Iowa State University Press. Iowa, USA. 227 p. [ Links ]
SPERRY, J. S.; SALIENDRA, N. Z. 1994. Intra and inter–plant variation in xylem cavitation in Betula occidentalis. Plant, Cell and Environment 17(11): 1233–1241. [ Links ]
VERDUGO, V.; ROJAS, A.; DE LEÓN, A.; ZAMBRANO, B.; BARRIOS, S.; LEÓN, E.; RIOS, B.; BENAVIDES, A. 1999. Estimación del índice estomático y la frecuencia estomática en cuatro variedades de ajo (Allium sativum L.). Curso de Fisiología de hortalizas. Maestría en Horticultura.UAAAN, Saltillo. Coahuila, México. [ Links ]
WILCOX C., D.; DOVE, S. B.; DOSS, W. M.; GREER, D. B. 2002. UTHSCSA Image Tool. IT Version 3.0 Departament of Dental Diagnostic Science. University of Texas Health Science Center, San Antonio, Texas, USA. 57 p. [ Links ]
WILKINSON, H. 1979. The plant superface (mainly leaf), pp. 97–165 In: Anatomy of Dicotyledons. METCALFE, C. R.; CHALK, L. (eds). Clarendon Press. Oxford, UK. [ Links ]
WILSON, D.; COOPER, J. P. 1970. Effect of selection for mesophyll cell size on growth and assimilation in Lolium perenne L. NewPhytologist. 69(2): 233–245. [ Links ]

