










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Chapingo. Serie horticultura
versión On-line ISSN 2007-4034versión impresa ISSN 1027-152X
Rev. Chapingo Ser.Hortic vol.16 no.2 Chapingo may./ago. 2010
Prohexadiona–Ca, una alternativa en la producción de tomate de cáscara (Physalis ixocarpa Brot.)
Prohexadione–Ca, an alternative in the production of husk–tomato (Physalis ixocarpa Brot.)
H. Ramírez1*; C. E. Rivera–Cruz1; A. Benavides–Mendoza1; V. Robledo–Torres1; G. Reyna–Sustaita2
1 Departamento de Horticultura, Universidad Autónoma Agraria Antonio Narro, Buenavista, Saltillo, Coahuila. MÉXICO.
2 Departamento de Ciencias Básicas. Universidad Autónoma Agraria Antonio Narro, Buenavista, Saltillo, Coahuila. MÉXICO. Correo–e: homeror@terra.com.mx (Autor responsable).
Recibido: 1 de diciembre, 2009.
Aceptado: 2 de junio, 2010.
Resumen
El tomate de cáscara, por su superficie cultivada y su constante aumento en el consumo per capita, se considera una de las principales hortalizas en México. Se cultiva en cualquier región del país, aunque con una producción media nacional por debajo de su potencial. Prohexadiona de Ca (P–Ca), es un retardante del crecimiento señalado recientemente como una alternativa prometedora en la horticultura moderna, principalmente para mejorar la producción y calidad del producto cosechado. Con base en lo anterior, se evaluaron los efectos de P–Ca sobre el crecimiento vegetativo y productivo; así como las propiedades químicas del fruto en tomate de cáscara. Se realizaron dos aplicaciones de P–Ca a diferentes dosis (0, 125, 175 y 200 mg·litro–1). La primera se realizó cuando las plantas presentaron primordios florales y la segunda 20 días después. Con un diseño en bloques completos al azar y una comparación de medias con la prueba DMS a una P<0.05, se demostró que P–Ca redujo significativamente la altura de la planta, aumentó notablemente el número de frutos, el rendimiento por planta (200 mg·litro–1), el contenido de vitamina C en frutos (175 y 200 mg·litro–1) y la actividad enzimática catalasa (175 mg·litro–1). Las variables de tamaño y peso de fruto, el contenido de proteína, la actividad enzimática peroxidasa y el contenido mineral en fruto no fueron afectados por P–Ca.
Palabras clave: P–Ca, vitamina C, retardante de crecimiento, tomate de cáscara.
Abstract
Husk tomato, because of its cultivated area and its constant increase in per capita consumption, is considered one of Mexico's main horticultural species. It is grown in almost all agriculture regions of the country, but the mean national yield per hectare of this crop is below its potential. Prohexadione–Ca (P–Ca) is a growth retardant recently named as a promising alternative for improving production and produce quality in modern horticulture. In this study, the effects of P–Ca on vegetative and productive growth as well as on the chemical properties of husk tomato fruits were assessed. The growth retardant was applied twice at 0 (control), 125, 175 and 200 m·gL–1. The first application was done when plants showed the first flower buds; the second application was 20 days later. The study was established in open field using a randomized block design and a DMS (Pd"0.05) statistical test. It was demonstrated t·hat P–Ca significantly reduced plant height, increased number of fruits and yield (200 m·gL–1) per plant, vitamin C content in fruits (175 and 200 m·gL–1) and catalase activity (175 m·gL–1). Other parameters such as weight and size of fruits, protein content, peroxidase activity and nutrient content were not affected by P–Ca.
Key words: P–Ca, vitamin C, growth retardant, husk–tomato.
INTRODUCCIÓN
El tomate de cáscara (Physalis ixocarpa Brot.) es la quinta hortaliza de mayor importancia en México por su superficie cultivada (SIAP, 2009). En años recientes, el consumo per cápita de tomate de cáscara en México ha ido en aumento. Esta especie se localiza en diferentes regiones del país, ya sea en forma silvestre, cultivada o mejorada; y se cultiva desde los 10 a los 2,600 m de altitud. En México se reporta una media de producción de 12 t·ha–1, cuando su potencial productivo alcanza en trabajos experimentales hasta las 40 t·ha–1 (Peña, 2001). Por lo tanto, es oportuno considerar nuevas alternativas de producción.
El uso de biorreguladores que actúen en armonía con la naturaleza y no causen efectos adversos en la salud humana, abren la posibilidad de ser utilizados en horticultura para aumentar y mejorar la producción. El uso de estas substancias tienen la ventaja de producir efectos benéficos en hortalizas de acuerdo a las necesidades del productor (Rojas y Ramírez, 1993). Los retardantes de crecimiento vegetativo favorecen el cuajado de frutos, debido a que inhiben la síntesis de giberelinas. Prohexadiona de Calcio (P–Ca) es un retardante de crecimiento que bloquea la biosíntesis de giberelinas en ápices, reduce el crecimiento vegetativo, induce la formación de yemas florales e incrementa el cuajado de fruto (Unrath, 1999; Ramírez et al., 2005).
Con la visión de obtener un aumento y mejora en la producción de tomate de cáscara y de analizar otros posibles efectos del retardante vegetativo P–Ca en el cultivo, se realizó este estudio planteando como objetivos, el evaluar los efectos de este retardante de crecimiento en parámetros hortícolas en planta y en fruto de tomate de cáscara, y conocer los posibles cambios en propiedades bromatológicas en el fruto.
MATERIALES Y MÉTODOS
La presente investigación se desarrolló en la UniversidadAutónoma AgrariaAntonio Narro, Unidad Saltillo, durante el periodo 2008 – 2009. Se utilizaron semillas certificadas de tomate de cáscara (Physalis ixocarpa Brot.) variedad Rendidora. Las semillas fueron sembradas en charolas de poliestireno de 200 cavidades. Cuando las plántulas presentaron 15 cm de altura en promedio, se trasplantó a campo abierto con 1.20 m entre surcos y 30 cm entre plantas (una densidad de 22,222 plantas·ha–1).
Se evalúo la aplicación de tres dosis de P–Ca, 125, 175, 200 mg·litro–1 y un testigo (agua). Se realizaron dos aplicaciones de P–Ca; la primera cuando las plantas presentaban primordios florales y la segunda 20 días después. La aplicación de P–Ca se realizó por la mañana sobre el follaje de las plantas a punto de goteo. Para el análisis estadístico de los datos, se utilizó un diseño en bloques completos al azar y la prueba de DMS (P<0.05) para la comparación de medias. Las variables evaluadas fueron: altura final de la planta, número de frutos por planta, tamaño de frutos (diámetro polar y ecuatorial de fruto), peso promedio de fruto, rendimiento, contenido de vitamina C, proteína, actividad enzimática (catalasa y peroxidasa) y el contenido mineral en frutos.
Parámetros hortícolas
La altura final de la planta se determinó midiendo con un cinta métrica escala 0 a 5 m desde la base del tallo hasta el ápice de la planta al final del ciclo. Se realizaron tres cortes, en 15 plantas de cada tratamiento para determinar el número de frutos por planta. El tamaño y el peso promedio de fruto se determinó tomando como muestra tres frutos por planta. El rendimiento por planta se determinó con el peso del número total de frutos por planta, utilizando una báscula Ohaus modelo 3729 con capacidad máxima de 3,000 gramos y resolución de 0.1 gramos.
Propiedades químicas del fruto
El contenido de vitamina C en fruto, se determinó por el método de titulación (Padayatt et al., 2001). Se tomaron muestras de fruto en fresco de 10 g, se trituró juntamente con 10 ml de ácido clorhídrico 2 %, se filtró y se aforó a 100 ml con agua destilada en un matraz Erlenmeyer. Con 10 ml del diluido, se tituló con el 2,6 diclorofenolindofenol (1x10–3 N) y se determinó el contenido de vitamina C con la fórmula:

El contenido de proteína en fruto, se determinó por el método Kjeldahl (A.O.A.C., 1980), el cual consiste en un proceso de digestión y destilación; un matraz Kjeldahl con 1.5 g de fruto seco y molido, una cucharada de mezcla catalítica (sulfato de potasio+sulfato de cobre), 25 ml de ácido sulfúrico (H2SO4 98 %), y cuatro perlas de vidrio, se coloca en una estufa para la digestión hasta tener un líquido de color claro. La muestra digerida, mezclada con 250 ml de agua destilada, 110 ml de hidróxido de sodio (NaOH 45 %) y gránulos de Zinc, se destila; recibiendo el destilado en matraces con 50 ml de ácido bórico (H3BO3 4 %) y colorante mixto. El destilado colectado se titula con H2SO4 (0.1N) y se calculó el porcentaje de proteína en frutos mediante la fórmula:
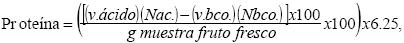
donde, v. ácido es el volumen gastado de H2SO4 (0.1N) para titular la muestra, Nac. y Nbco. es la normalidad del H2SO4 (0.1N), v.bco. es el volumen gastado de H2SO4 (0.1N) para titular el blanco y 6.25 es el factor de conversión generalizado, en este caso para tomate de cáscara.
Para la determinación de la actividad enzimática catalasa (CAT), la extracción se realizó de 0.5 g de pulpa de tomate de cáscara en 5 ml de buffer de fosfatos 100 mM (pH 7), 50 mg de polivinilpirrodiona, en un mortero enfriado a 4 °C, se centrifugó a 183. 3 Hz por 11 min a 4 °C. Del sobrenadante se obtuvo la enzima (Masia, 1998). Luego se prepararon 5 ml de la mezcla de reacción que contenía: 300 μM de buffer fosfatos 100 Mm (pH 6.8), 100 μM de H2O2 y 1 ml de sobrenadante con la enzima diluido 1:20 (v/v). La mezcla de reacción se incubó por 1 min a temperatura constante de 25 °C, la reacción fue detenida al agregar 10 ml de H2SO4 al 2 % (v/v). El H2O2 residual se tituló con una solución de KMnO4 (0.2 M) hasta obtener un color púrpura débil que resistió al menos 15 segundos. La actividad de la enzima se calculó con la fórmula: 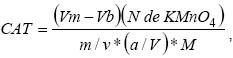 donde, Vm es el volumen gastado de KMnO4 para titular la muestra, Vb es el volumen gastado de KMnO4 para titular el blanco, N de KMnO4 es la normalidad de KMnO4, m son los gramos de pulpa de fruto, v es el volumen de dilución de la pulpa, a/V es la dilución de la enzima extraída (1:20) y M es el volumen de la mezcla de reacción.
donde, Vm es el volumen gastado de KMnO4 para titular la muestra, Vb es el volumen gastado de KMnO4 para titular el blanco, N de KMnO4 es la normalidad de KMnO4, m son los gramos de pulpa de fruto, v es el volumen de dilución de la pulpa, a/V es la dilución de la enzima extraída (1:20) y M es el volumen de la mezcla de reacción.
La actividad enzimática peroxidasa (PX) se determinó de la siguiente manera, se homogenizaron 0.5 g de pulpa de tomate de cáscara con 5 ml de buffer fosfatos 100 mM (pH 6.8) en un mortero enfriado a 4 °C. Se centrifugó a 216.66 Hz por 15 min a 4 °C. El sobrenadante que contiene la enzima peroxidasa se preparó y diluyó en proporción 1:20 (v/v). La actividad enzimática se determinó con 125 μM de buffer fosfatos 100 mM (pH 6.8), 50 μM de pirogagol, 50 μM de H2O2 y 1 ml de extracto de enzima diluido 1:20 para obtener 5 ml de volumen. La mezcla de reacción se incubó por 1 min a 25 °C, se añadieron 0.5 ml de H2SO4 al 5 % (v/ v) para detener la reacción. La concentración de purpurogalina se mide a 420 nm de absorbancia (A420) (Kar y Mishra, 1976). La actividad de la enzima se calculó con la fórmula: 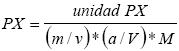 donde, unidad PX es, por cada 0.1 absorbancia equivale a la unidad de peroxidasa, m son los gramos de pulpa de fruto, v es el volumen de dilución de la pulpa, a/V es la dilución de la enzima extraída (1:20) y M es el volumen de la mezcla de reacción.
donde, unidad PX es, por cada 0.1 absorbancia equivale a la unidad de peroxidasa, m son los gramos de pulpa de fruto, v es el volumen de dilución de la pulpa, a/V es la dilución de la enzima extraída (1:20) y M es el volumen de la mezcla de reacción.
El contenido de elementos minerales se determinó como sigue: El nitrógeno (N) fue determinado en el mismo proceso en que se determinó proteína, con el método de Kjeldahl (A. O. A. C., 1980), y se utilizó la fórmula: 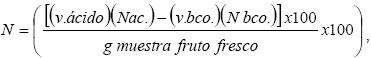 donde, v. ácido es el volumen gastado de H2SO4 (0.1N) para titular la muestra, Nác. y Nbco. es la normalidad del H2SO4 (0.1N), v. bco. es el volumen gastado de H2SO4 (0.1N) para titular el blanco. La determinación de calcio (Ca), sodio (Na), potasio (K), zinc (Zn), cobre (Cu), magnesio (Mg) y de manganeso (Mn) se realizó por el método de absorción atómica (A. O. A. C., 1980), en la cual 1 g de fruto seco y molido se incinera. Un matraz con el incinerado junto con la mezcla perclórica (ácido nítrico y ácido perclórico 3:1 (v/v)) se coloca en una estufa para la digestión hasta obtener un líquido de color claro. El digerido se filtra (con papel filtro 42 sin cenizas) a un matraz y se afora a 100 ml con agua desionizada. Se toma lectura de las muestras en el espectrofotómetro de absorción atómica. El contenido para K, se determinó con la fórmula;
donde, v. ácido es el volumen gastado de H2SO4 (0.1N) para titular la muestra, Nác. y Nbco. es la normalidad del H2SO4 (0.1N), v. bco. es el volumen gastado de H2SO4 (0.1N) para titular el blanco. La determinación de calcio (Ca), sodio (Na), potasio (K), zinc (Zn), cobre (Cu), magnesio (Mg) y de manganeso (Mn) se realizó por el método de absorción atómica (A. O. A. C., 1980), en la cual 1 g de fruto seco y molido se incinera. Un matraz con el incinerado junto con la mezcla perclórica (ácido nítrico y ácido perclórico 3:1 (v/v)) se coloca en una estufa para la digestión hasta obtener un líquido de color claro. El digerido se filtra (con papel filtro 42 sin cenizas) a un matraz y se afora a 100 ml con agua desionizada. Se toma lectura de las muestras en el espectrofotómetro de absorción atómica. El contenido para K, se determinó con la fórmula; 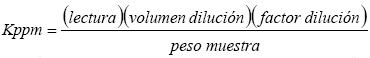 y el resto de los minerales con la fórmula,
y el resto de los minerales con la fórmula, 
RESULTADOS Y DISCUSIÓN
Altura de planta
La Figura 1 muestra que Prohexadiona–Ca originó efectos significativos (P<0.05) en la altura de la planta de tomate de cáscara.

Los resultados ilustran que la aplicación de P–Ca redujo de forma significativa la altura de las plantas al compararse con las del testigo. Este efecto en la altura de la planta, la cual disminuyó hasta un 19.84 %, es consistente con lo reportado por Ramírez et al. (2006) en manzano "Golden Delicious". La reducción de altura de laplanta está directamente relacionado con un bloqueo por parte de P–Ca en la biosíntesis de las giberelinas A1, A4 y A7 biológicamente activas (Evans et al., 1997) como lo ha reportado Ramírez et al. (2008).
Número de frutos por planta
Los efectos de la aplicación de Prohexadiona–Ca en el número de frutos por planta se muestran en el Cuadro 1. Se observó que los tratamientos de 175 y 200 mg·L–1 mostraron un aumento significativo (P<0.05) en frutos por planta, con incrementos del 26 y 60 %, respectivamente. Efectos similares se han observado en manzano (Ramírez et al., 2006) y tomate "Saladette" (Ramírez et al., 2005). En esta investigación se logró obtener tres cortes de tomate con una buena producción y calidad de fruto (Cuadro 1). El retardo temporal de crecimiento en la planta, principal propiedad de la hormona P–Ca, beneficia al proceso de fructificación; así como el aceleramiento y aumento de yemas florales y amarre de fruto, debido a que la modificación en el flujo de fotoasimilados hacia tejidos meristemáticos favorece a esos procesos (Jackson y Looney, 2003; Sánchez, 2003; Pilatti, 1997).
Peso promedio y tamaño de fruto
La aplicación de P–Ca no afectó el peso promedio de fruto. Con frecuencia se observa en manzano una reducción en el peso de frutos cuando se aplican retardantes de crecimiento como el Alar y P–Ca (Basak y Rademacher, 2000). Lo encontrado en este estudio resulta interesante, considerando que el peso del fruto no se afectó (Cuadro 1). Resultados similares han sido reportados por Ramírez et al. (2005) en tomate "Saladette" y por Dayatilake et al. (2005) en manzano.
Prohexadiona–Ca aplicado a plantas de tomate de cáscara no afectó el tamaño de fruto (Cuadro 1). De forma importante se observó un aumentó en el número de frutos por planta sin afectar el tamaño de estos. Efectos similares reporta Dayatilake et al. (2005) en frutos de manzano al aplicar la P–Ca.
El hecho de no haberse reducido el tamaño y peso de los frutos con el P–Ca, representa un importante valor agregado en el producto cosechado. Este resultado se podría explicar con experiencias de otros autores como lo reportado por Ramírez et al. (2005), ellos encontraron que con la aplicación de P–Ca a plantas de tomate se aumentó significativamente el contenido de citocininas, las cuales junto con las auxinas presentes en semillas inmaduras de frutos jóvenes, influyen en la regulación de la división y alargamiento celular, y por lo tanto inductores del crecimiento y peso de fruto, como lo mencionan Raven et al. (1992). En relación a lo anterior, Coleto (1995) menciona que después de la división celular inicia la acumulación de los fotosintatos y con ello el crecimiento y peso del fruto. Por lo tanto, es probable que la aplicación de P–Ca al reducir la demanda de asimilados y otras hormonas en tejidos de crecimiento apical, favoreciera el contenido de citocininas y la acumulación de fotosintatos en fruto, evitando con esto reducción de peso y tamaño en ellos a pesar de un mayor número por planta como resultado de la aplicación del retardante.
Rendimiento por planta
El rendimiento por planta se incrementó con Prohexadiona–Ca (Figura 2). En el tratamiento de 200 mg·L–1 se observó una diferencia significativa (P<0.05) ante el resto de los tratamientos, con un incremento de hasta un 83 % en comparación con el testigo. En las dosis de 125 y 175 mg·L–1, aunque no significativo, se observó un 19 y 20 % de aumento respectivamente. Los incrementos en el rendimiento reflejan principalmente los efectos positivos en el mayor número de frutos con la aplicación de Prohexadiona–Ca (Cuadro 1). Estos resultados se fortalecen con lo reportado en tomate "Saladette" al aplicar 175 y 200 mg·L–1 de P–Ca (Ramírez et al., 2005) y en cultivos como manzano (Ramírez et al., 2006; Dayatilake et al., 2005) y pera (Costa et al., 2004).

Vitamina C
En la Figura 3 se muestran los efectos de los tratamientos en los valores de la concentración de vitamina C en frutos de tomate de cáscara. Los resultados obtenidos muestran que los tratamientos con P–Ca incrementaron el contenido de vitamina C en comparación con el testigo. Frutos tratados con P–Ca a dosis de 175 y 200 mg·L–1 aumentaron de forma significativa (P<0.05) el contenido del antioxidante con un 33 y 45 %, respectivamente. La vitamina C es un antioxidante importante como elemento nutritivo para la salud humana, se le atribuye el fortalecimiento del organismo en defensa a enfermedades cardiovasculares (Carr y Frei, 1999). El aumento significativo de la vitamina C como efecto de la aplicación del retardante de crecimiento vegetativo, se puede derivar de un estímulo en la biosíntesis del ácido ascórbico sintetizado con base en la oxidación a través de la enzima oxidasa de ácido ascórbico como lo han sugerido Chaudhary et al. (2006), y Melo y Cuamatzi (2007). Se han mostrado incrementos de vitamina C con la aplicación de retardantes de crecimiento como el paclobutrazol (Jamalian et al., 2009), pero con la desventaja de que éste persiste en la planta y produce efectos tóxicos en humanos, con lo que se prohibe su uso en cultivos hortícolas (Owens y Stover, 1999). El retardante P–Ca no causa esos efectos adversos (Rademacher, 2004), por lo tanto, el aumento en el contenido de Vitamina C en frutos de tomate de cáscara, confirma la importancia de P–Ca como una alternativa en la tecnología aplicada a la horticultura moderna (Ramírez, 2003).

Proteína
Los resultados obtenidos mostraron que la aplicación de Prohexadiona–ca no afectó el contenido de proteína en frutos. De forma interesante P–Ca mantiene los niveles de proteína (Cuadro 2) con un importante incremento en el número de frutos y rendimiento por planta, por lo que, probablemente el reducir el crecimiento vegetativo con P–Ca se reduce la competencia por el flujo de asimilados entre el proceso de fructificación y el de crecimiento vegetativo, beneficiándose con esto el desarrollo de frutos (Ramírez et al., 2005; Rademacher y Kober, 2003). El retardante aplicado indujo que las plantas respondieran de forma suficiente en el abastecimiento de fotosintatos a frutos, los cuales son básicos en la síntesis de aminoácidos y péptidos utilizados en la síntesis de proteínas (Armstrong y Bennett, 1982). Las proteínas son macromoléculas que realizan una diversidad funcional mucho mayor que otras en el metabolismo de las plantas (Campbell y Farrel, 2004), por lo que, mantener los niveles de proteína resulta de gran importancia para no afectar los procesos metabólicos y con ello la producción y calidad de frutos.

Catalasa
La aplicación de Prohexadiona–Ca a plantas de tomate de cáscara indujo efectos en la actividad enzimática catalasa en frutos en proceso de maduración.
Con la aplicación de 175 mg·L–1 de P–Ca se manifestó un aumento significativo (P<0.05) en la actividad enzimática catalasa. La Figura 4 ilustra que al aumentar la dosis de P–Ca a 200 mg·L–1 el contenido de actividad catalasa no mostró diferencias significativas ante el testigo y es superada por el valor obtenido con la aplicación de 175 mg·L–1. Los resultados podrían reflejar un efecto de P–Ca en la actividad de catalasa como punto máximo de estimulo con la dosis de 175 mg·L–1. Los niveles de catalasa se relacionan con una mayor tolerancia al estrés oxidativo, causado por factores ambientales como el frío (Jiménez et al., 2002; Lafuente et al., 2004) o patógenos (Beltrán et al., 2006). Con base en esas experiencias, es probable que el efecto de P–Ca pueda también mediar a través de una fortaleza adicional en el tejido en donde esté actuando el retardante del crecimiento (Fernando y Jones, 1999; Roemmelt et al., 1999; Costa et al., 2006; Rademacher et al., 2006; Spinelli et al., 2006; Greene, 2008).

Peroxidasa
Los resultados del análisis estadístico a datos del contenido de la actividad enzimática peroxidasa en frutos de tomate de cáscara (Cuadro 2), no mostraron diferencias significativas entre los tratamientos. La actividad de esta enzima está relacionada con la senescencia y daños oxidativos (Zhou et al., 2003; Vicente et al., 2006). La peroxidasa degrada el H2O2 con lo que se le considera una enzima antioxidante (Nakano yAsada, 1981; Asada, 1997). Incrementos en el contenido de peroxidasa se ha asociado al proceso de maduración (Robinson, 1991; Alia et al., 2002). Los frutos analizados en este trabajo se tomaron en un estado de madurez comercial (fisiológicamente verdes), y no fisiológicamente maduros. Lo anterior, con base en el atractivo que tiene esta hortaliza entre el consumidor en ese estadio y que es un fruto de tomate color verde y ácido. Esto, podría explicar la no significancia estadística entre los tratamientos.
Minerales en fruto
No se observaron diferencias significativas en los niveles de nutrientes entre tratamientos. Los resultados obtenidos muestran que la aplicación de prohexadiona–Ca no afectó el contenido mineral en frutos de tomate de cáscara "Rendidora". El flujo de minerales en plantas es variado y depende de la disponibilidad y demanda entre órganos de la planta y la etapa de desarrollo de la misma (Gutiérrez, 1997). Es posible que al reducirse el crecimiento vegetativo por la presencia de P–Ca en el tejido, se modifique el patrón de traslocación de minerales y éstos sean enviados en mayor proporción hacia los frutos en desarrollo y su distribución sea más equitativa entre ellos; reflejándose en lo observado el no afectarse adversamente el contenido y los niveles de los mismos (Rademacher, 2004). Los elementos minerales, son necesarios para el metabolismo de la planta, cumplen funciones como constituyentes de moléculas orgánicas, en la reserva energética, de forma iónica y reacciones redox (Raven et al., 1992). La deficiencia de minerales, se refleja principalmente en frutos, induce desórdenes fisiológicos que se traducen en bajas en el rendimiento, tamaño, peso, color, forma, sabor y calidad nutritiva en fruto (Martínez et al., 2008; Mancera et al., 2007), lo que demuestra la gran importancia de no afectar los niveles en el contenido mineral; a pesar de que son diversos los factores que pudieran influenciar en la absorción, movilidad y asimilación de los elementos minerales en la planta (Shaviv y Mikkelsen, 1993). (Cuadro 3)
CONCLUSIONES
Con base en los resultados obtenidos y bajo las condiciones en que se realizó el trabajo, la aplicación de Prohexadiona–Ca (175 y 200 mg·L–1) a plantas de tomate de cáscara variedad Rendidora, reduce la altura final de la planta e incrementa el rendimiento, sin afectar el tamaño, peso, contenido de proteína, de peroxidasa y minerales en fruto. El contenido de vitamina C y la actividad catalasa se incrementa significativamente con la aplicación del retardante de crecimiento.
LITERATURA CITADA
ALIA, I.; COLINAS, M. T.; MARTÍNEZ, M. T.; SOTO, M. R. 2002. Factores fisiológicos, bioquímicos y de calidad en frutos de zapote Mamey (Pouteria sapota Jacq. H. E. Moore & Stearn) durante poscosecha. Chapingo Horticultura 8(2): 263–281. [ Links ]
A.O.A.C. 1980. Association of Official Agricultural Chemists. Official Methods of Analysis. Washington, D. C. [ Links ]
ARMSTRONG, F. B.; BENNETT, T. P. 1982. Bioquímica. Traducido al español por CUCHILLO, C. Editorial Reverte. Barcelona, España. 522 p. [ Links ]
ASADA, K. 1997. The role of ascorbate peroxidase and monodehydroascorbate reductase in H2O2 scavenging in plants. In: Oxidative stress and molecular biology of antioxidant defenses. SCANDALIOS, J. G. (ed). Cold Spring Harbor Laboratory Press. Plainview, New York, EE. UU. pp. 715–735. [ Links ]
BASAK, A.; RADEMACHER, W. 2000. Growth regulation of pome and stone fruit trees by use of Prohexadione–Ca. Acta Horticulturae 514: 41–50. [ Links ]
BELTRAN, M. J.; OGURA, T.; MANZO, G.; ARIAS, C. 2006. Catalasas de hongos fitopatogenos: ¿factores de virulencia y resistencia a los fungicidas? Revista Mexicana de Fitopatología 24(1): 50–58. [ Links ]
CAMPBELL, M. K.; FARREL, S. O. 2004. Bioquímica. Cuarta edición. Traducido al español por AGUILAR, Q. M. T. Editorial Thomson. D.F., México. 812 p. [ Links ]
CARR, A. C.; FREI, B. 1999. Toward a new recommended dietary allowance for vitamin C based on antioxidant and health effects in humans. American Journal of Clinical Nutrition 69(6): 1086–107. [ Links ]
CHAUDHARY, B. R.; SHARMA, M. D.; SHAKYA, S. M.; GAUTAM, D. M. 2006. Effect of plant growth regulators on growth, yield and quality of chilli (Capsicum annuum L.) at Rampur, Chitwan. Journal of the Institute of Agriculture and Animal Science 27: 65–68. [ Links ]
COLETO, J. M. 1995. Crecimiento y Desarrollo de las Especies Frutales. Segunda edición. Editorial Mundi–Prensa. Madrid, España. 168 p. [ Links ]
COSTA, G.; ANDREOTTI, C.; SPINELLI, F.; RADEMACHER, W. 2006. Prohexadione–Ca: More t·han a growth regulator for pome fruit trees. Acta Horticulturae 727: 107–116. [ Links ]
COSTA, G.; SABATINI, E.; SPINELLI, F.; ANDREOTTI, C.; BOMBEN, C.; VIZZOTO, G. 2004. Two years of application of Prohexadione–ca on apple: Effect on vegetative and cropping performance, fruit quality, return bloom and residual effect. Acta Horticulturae 653: 35–40. [ Links ]
DAYATILAKE, G. A.; WÜNSCHE, J. N.; WOOD, P.; MCARTHEY, S.; MANKTELOW, D.; LO, P.; GURNSEY, S.; TUSTIN, D. S. 2005. The use of prohexadione–ca for improved crop management. Acta Horticulturae 694: 315–319. [ Links ]
EVANS, J. R.; ISHIDA, C. A.; REGUSCI, C. L.; RADEMACHER, W. 1997. Mode of action, metabolism and uptake of BAS–125W, prohexadione–calcium. HortScience 32: 558. [ Links ]
FERNANDO, W. G. D.: JONES, A. L. 1999. Prohexadione calcium – a tool for reducing secondary fire blight infection. Acta Horticulturae 489: 597–600. [ Links ]
GREENE, D. 2008. The effect of repeat annual applications of prohexadione–calcium on fruit set, return bloom and fruit size of apples. HortScience 43: 376–379. [ Links ]
GUTIÉRREZ, M. V. 1997. Nutrición mineral de las plantas: avances y aplicaciones. Agronomía Costarricense 21(1): 127–137. [ Links ]
JACKSON, D. I.; LOONEY, N. E. 2003. Utilización de biorreguladores en fruticultura. In: Producción de frutas de climas templados y subtropicales. JACKSON, D. I. (ed). Ed. Acribia. Zaragoza, España. pp. 119–120. [ Links ]
JAMALIAN, S.; TEHRANIFAR, A.; TAFAZOLI, E.; ESHGHI, S.; DAVARYNEJAD, G. H. 2009. Paclobutrazol can reduce the negative effects of salinity on reproductive growth, yield and fruit quality of strawberry plant. Acta Horticulturae 842: 825–828. [ Links ]
JIMENEZ, A.; CREISSEN, G.; KULAR, B.; FIRMIN, J.; ROBINSON, S.; VERHOEYEN, M.; MULLINEAUX, P. 2002. Changes in oxidative processes and components of the antioxidant system during tomato fruit ripening. Planta 214(5): 751–758. [ Links ]
KAR, M.; MISHRA, D. 1976. Catalase, peroxidase, and polyphenoloxidase activities during rice leaf senescence. Plant Physiology 57(2): 315–319. [ Links ]
LAFUENTE, M. T.; SALA, J. M.; ZACARIAS, L. 2004. Active oxygen detoxifying enzymes and phenylalanine ammonia–lyase in the ethylene–induced chilling tolerance in citrus fruit. Journal Agriculture Food Chemistry 52(11): 3606–3611. [ Links ]
MANCERA, M. M.; SOTO J. M.; SÁNCHEZ, E.; YAÑEZ, R. M.; MONTES, F.; BALANDRAN, R. R. 2007. Caracterización mineral de manzana 'Red Delicious' y 'Golden Delicious' de dos países productores. Tecnociencia Chihuahua 1(2): 6–17. [ Links ]
MARTINEZ, F. E.; SARMIENTO, J.; FISCHER, G.; JIMÉNEZ, F. 2008. Efecto de la deficiencia de N, P, K, Ca, Mg y B en componentes de producción y calidad de la uchuva (Physalis peruviana L.). Agronomía Colombiana 26(3): 389–398. [ Links ]
MASIA, A. 1998. Superoxide dismutase and catalase activities in apple fruit during ripening and post–harvest and with special reference to ethylene. Physiology Plantarum 104(4): 668–672. [ Links ]
MELO, V.; CUAMATZI, O. 2007. Bioquímica de los procesos metabólicos. Segunda edición. Editorial Reverté. D.F., México. 406 p. [ Links ]
NAKANO, Y.; ASADA, K. 1981. Hydrogen peroxide is scavenged by ascorbate–specific peroxidase in spinach chloroplasts. Plant and Cell Physiology 22(5): 867–880. [ Links ]
OWENS, L.; STOVER, E. 1999. Vegetative growth and flowering of young apple trees in response to prohexadiona–calcium. HortSience 34: 1194–1196. [ Links ]
PADAYATT, S. J.; DARUWALA, R.; WANG, Y.; ECK, P. K.; SONG, J.; KOH, W. S.; LEVINE, M. 2001. Vitamin C: from molecular actions to optimum intake. In: Handbook of Antioxidants. CADENAS, E.; PACKER, L. (eds) 2nd edition. CRC Press. Washington DC, EE.UU. pp. 117–145. [ Links ]
PEÑA, A. 2001. Situación actual y perspectivas de la producción y mejoramiento genético de tomate de cáscara (Physalis ixocarpa Brot.) en México. Primer Simposio Nacional. Técnicas modernas de producción de tomate, papa y otras solanáceas. Universidad Autónoma Agraria Antonio Narro. Saltillo, Coahuila. 10 p. [ Links ]
PILATTI, R. A. 1997. Cultivo Bajo Invernaderos. Ed. Hemisferio Sur, S. A. Universidad Nacional del Litoral. Buenos Aires, Argentina. pp. 7–33. [ Links ]
RADEMACHER, W. 2004. Chemical regulation of shoot growth in fruit trees. Acta Horticulturae 653: 29–32. [ Links ]
RADEMACHER, W.; KOBER, L. 2003. Efficient use of prohexadione–ca in pome fruits. European Journal of Horticultural Science 68(3): 107. [ Links ]
RADEMACHER, W.; SPINELLI, F.; COSTA, G. 2006. Prohexadione–ca: modes of action of a multifunctional plant bioregulator for fruit trees. Acta Horticulturae 727: 97–106. [ Links ]
RAMÍREZ, H. 2003. El uso de hormonas en la producción de cultivos hortícolas para exportación. Memoria del Tercer Simposio Nacional de Horticultura. Saltillo, Coahuila, México. pp. 122. [ Links ]
RAMÍREZ, H.; ALONSO, S.; BENAVIDES, A. 2006. Prohexadione–ca modifies growth and endogenous hormones in the shoot apex in apple trees. Acta Horticulturae 727: 117–124. [ Links ]
RAMÍREZ, H.; PERALTA, R.; BENAVIDES, A.; SÁNCHEZ,A.; ROBLEDO, V.; HERNÁNDEZ, J. 2005. Efectos de prohexadiona–ca en tomate y su relación con la variación de la concentración de giberelinas y citocininas. Chapingo Horticultura 11(2): 283–290. [ Links ]
RAMÍREZ, H.; HERRERA, B.; MÉNDEZ, Y. H.; BENAVIDES, A.; DE LA CRUZ, J. A.; ÁLVAREZ, V.; VILLAREAL, J. A. 2008. Prohexadiona de calcio disminuye el contenido de giberelinas endógenas en ápices de tomate Saladette y chile pimiento. Chapingo Horticultura 14(2): 193–198. [ Links ]
RAVEN, P. H.; EVERT, R. F.; EICHHORN, S. E. 1992. Biología de las plantas. Volumen 2. Traducido al español por SANTAMARÍA, S.; LLORET, F.; MAS, M.; CARDONA, M. A. Editorial Reverté. Barcelona, España. 773 p. [ Links ]
ROBINSON, D. S. 1991. Peroxidases and catalases in foods. In: Oxidative enzymes in foods. ROBINSON, D. S.; ESKIN, N. A. (eds). Elsevier Applied Science. LTD. England. pp. 1–9. [ Links ]
ROEMMELT, S.; TREUTTER, D.; SPEAKMAN, J. B.; RADEMACHER, W. 1999. Effects of prohexadione–ca on the flavonoid metabolism of apple with respect to plant resistance against fire blight. Acta Horticultirae 489: 359–364. [ Links ]
ROJAS, M.; RAMÍREZ, H. 1993. Control hormonal del desarrollo de las plantas. Segunda Edición. Editorial Limusa. D.F., México. 239 p. [ Links ]
SANCHEZ, F. 2003. Obtención de plantas ornamentales compactas, mediante la aplicación de paclobutrazol y podas de formación. Resúmenes del X Congreso Nacional de la Sociedad Mexicana de Ciencias Hortícolas, IX Congreso Nacional y II Internacional de Horticultura Ornamental. Chapingo, México. p. 169. [ Links ]
SHAVIV, A.; MIKKELSEN, R. L. 1993. Controlled–release fertilizers to increase efficiency of nutrient use and minimize environmental degradation–a review. Fertilizer Research 35(1–2): 1–12. [ Links ]
SISTEMA DE INFORMACIÓNAGROALIMENTARIA Y PESQUERA(SIAP). 2009. Anuario estadístico de la producción agrícola 2008; Tomate verde. Secretaría de agricultura, ganadería, desarrollo rural, pesca y alimentación. México. Consulta 7 de Septiembre de 2009; www.siap.gob.mx [ Links ]
SPINELLI, F.; COSTA, G.; SPEAKMAN, J. B.; RADEMACHER, W.; HALBWIRTH, H.; STICH, K. 2006. Prohexadione–calcium induces in apple the biosynthesis of luteoforol, a novel flavan 4–ol, which is active against Erwinia amylovora. Acta Horticulturae 704: 239–244. [ Links ]
UNRATH, C. R. 1999. Prohexadione–Ca: A promising chemical for controlling vegetative growth of apples. HortScience 34: 1197–1200. [ Links ]
VICENTE, A.; MARTÍNEZ, G.; CHAVES, A.; CIVELLO, P. 2006. Effect of heat treatment on strawberry fruit damage and oxidative metabolism during storage. Postharvest Biology and Technology 40(2): 116–122. [ Links ]
ZHOU, Y.; DAHLER, J.M.; UNDERHILL, S. J. R.; WILLS, R.B.H. 2003. Enzymes associated with blackheart development in pineapple fruit. Food Chemistry 80(4): 565–572. [ Links ]

