










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista Chapingo. Serie horticultura
versão On-line ISSN 2007-4034versão impressa ISSN 1027-152X
Rev. Chapingo Ser.Hortic vol.16 no.1 Chapingo Jan./Abr. 2010
Variación anatómica del xilema en tallo de cultivares de tomate injertados en un tipo criollo
Variation in stem xylem anatomy of tomato cultivars grafted onto a local race
A. Sory Toure1; R. Nieto-Ángel2*; J. E. Rodríguez-Pérez2; A. F. Barrientos-Priego2; L. A. Ibáñez-Castillo1; E. Romanchik K.1; C. A. Núñez-Colín2
1 Posgrado en Ingeniería Agrícola y Uso Integral del Agua (IAUIA), Universidad Autónoma Chapingo, km 38.5 Carretera México-Texcoco, Chapingo, Estado de México. C. P. 56230. MÉXICO.
2 Instituto de Horticultura, Universidad Autónoma Chapingo, km. 38.5 Carretera México–Texcoco, Chapingo, Estado de México, C. P. 56230. MÉXICO (*Autor responsable).
Recibido: 27 de diciembre, 2007
Aceptado: 25 de febrero, 2009
Resumen
El tomate (Solanum lycopersicum L.) es una de las hortalizas más consumidas, sin embargo, por algunos problemas fitopatológicos se ha visto la necesidad de buscar alternativas, para la producción sin afectar la calidad, tales como el uso de portainjertos resistente a factores ambientales y enfermedades, lo que necesariamente se requiere de estudios para evaluar la compatibilidad vegetativa y la resistencia a dichos factores, y uno de ellos es la evaluación de los sistemas vasculares en los puntos de unión del injerto.
Por lo anterior este estudio tuvo como objetivo analizar la anatomía de los vasos del xilema e índices de conducción de agua del portainjerto criollo de tomate rojo al que se le injertaron tres diferentes cultivares comerciales.
La anatomía del xilema de los cultivares injertados y del portainjerto criollo presentaron cambios con respecto a las plántulas sin injertar. El área, perímetro, longitud del eje mayor y menor de los vasos del xilema de los tallos de plantas, tuvieron valores estadísticamente superiores al realizar los injertos, no así para 'Charleston'/'Criollo de Honduras', 'Sun 7705'/'Criollo de Honduras' y viceversa; con relación a 'Charleston' y 'Sun 7705' sin injertar. 'Reserva F1', en las dos combinaciones ('Reserva F1'/Criollo de Honduras'; 'Criollo de Honduras'/'Reserva F1') y sin injertar, no manifestaron cambios en su anatomía. 'Charleston' presentó en las mismas características los máximos valores con relación a los otros cultivares; además que en las dos combinaciones de 'Charleston' con relación a las combinaciones, 'Charleston'/'Criollo de Honduras' y 'Criollo de Honduras'/'Charleston', también fueron superiores con relación a los otros cultivares.
En general los valores de las variables estudiadas de la anatomía de los vasos del xilema se incrementaron, tanto en los cultivares como en el portainjerto al tener la relación injerto/portainjerto.
Palabras clave: Solanum lycopersicum L., vasos del xilema, compatibilidad del injerto.
Abstract
The tomato (Solanum lycopersicum L.) is one of the most widely consumed vegetables. However, phytopathogical problems have underlined the need to look for alternatives for its cultivation, such as grafting, using rootstock resistant to environmental factors and diseases. For this technique, compatibility between the graft and the rootstock must be taken into account, especially of the water conduction system. The variability in xylem and water conduction indexes in local race rootstock onto which three different commercial cultivars were grafted was studied and compared with non-grafted plants. The anatomy of the xylem of the grafted cultivars and the local race rootstocks exhibited differences with respect to the non-grafted plants. Area, perimeter, and axis length of the largest and smallest axes of stem xylem vessels were statistically superior in grafted plants, but not for 'Charleston'/'Criollo de Honduras', 'Sun 7705'/'Criollo de Honduras', and vice versa, with respect to 'Charleston' and 'Sun 7705' without grafts. 'Reserva F1', in both combinations ('Reserva F1'/Criollo de Honduras'; 'Criollo de Honduras'/'Reserva F1') and without grafts, showed no changes in anatomy. The same characteristics in 'Charleston' had the maximum values with respect to the other cultivars. In general, the values of the xylem vessel anatomy characteristics were increased, of both the cultivars and rootstocks in the scion/rootstock relationship.
Key words: Solanum lycopersicum L. xylem vessels, graft compatibility.
INTRODUCCIÓN
El tomate (Solanum lycopersicum L.), por su valor alimenticio, es una de las hortalizas más importantes en todo el mundo; su preferencia radica en su alto valor nutritivo y económico; se consume en fresco, en ensaladas, cocido o frito en muchos platillos y en menor escala como encurtido. El incremento anual de la producción en algunas regiones del mundo en los últimos años, se relaciona con la generación de nuevas variedades, de técnicas de cultivo más eficientes, del uso de plásticos a campo abierto, de la producción en invernaderos, del uso de la hidroponía en el que se utilizan sustratos inertes y fertilizantes solubles, y lógicamente, al aumento de la superficie cultivada. La producción mundial de tomates para el 2005 fue de 94.11 millones de toneladas (Anónimo, 2005). Los principales países productores de tomate fresco son China, India, Estados Unidos, Turquía e Italia. En México se cultivan alrededor de 20,000 ha, con una producción de 2.8 millones de toneladas (Anónimo, 2005).
Uno de los principales problemas que se han observado en la producción de tomate en todas las regiones del mundo son los fitosanitarios, el del cuello de la planta y la raíz, sobre todo cuando se cultiva en suelos arcillosos en donde generalmente se favorece la presencia de Fusarium, en tanto que en suelos relativamente arenosos, el problema más importante es la presencia de nemátodos; ambas condiciones han requerido la aplicación excesiva de pesticidas.
Una de las prácticas más comunes y funcionales para disminuir los excesos de pesticidas para el control de estas enfermedades es el uso de portainjertos, favoreciendo también la producción y calidad de frutos. Hoolbrook et al. (2002) reportan que la práctica de injertación en tomate, a nivel comercial, se ha utilizado desde 1970; los principales problemas que se han resuelto con el uso de portainjertos en esta especie son la presencia de nemátodos en la raíz, Fusarium en el cuello de la planta, tolerancia a la salinidad de los suelos y del agua de riego, sequía o excesos de humedad en el suelo (Ayre y Turgeon, 2004; Mitidieri et al., 2005, Lifschitz et al., 2006), así como una mayor adaptación a temperaturas y humedad relativa altas (Black et al., 2003).
El uso de combinaciones de tomate 'Monroe'/'Beaufort' tuvo efecto positivo en el rendimiento del fruto, en tanto que en la combinación 'Belle'/'Beaufort' la producción disminuyó en relación con plantas no injertadas; sin embargo, Kacjan y Osvald (2004) mencionan que el uso de potainjertos en hortalizas y en invernadero es una práctica altamente sustentable. Plantas de tomate injertadas a dos tallos tuvieron la misma producción por planta y el tamaño de frutos fue más uniforme en relación a las plantas no injertadas y a un tallo (Peil y Gálvez, 2004). Mitidieri et al. (2005) probaron la combinación de cultivares de tomate 'Superman' y 'Fortaleza', sobre 'Heman', híbrido interespecífico de Lycopersicum hirsutum por L. esculentum, y encontraron que las plantas injertadas mostraron porcentajes significativamente menores de plantas muertas hasta 30 días después del transplante y plantas con menos síntomas aéreos de ataque de nemátodos. Las plantas injertadas mostraron mayor producción y menor cantidad de frutos pequeños.
En injertos, la unión y continuidad de los vasos del xilema es una de las formas más confiables para estimar compatibilidad entre injerto/portainjerto, y conocer así los posibles problemas que pudieran ocurrir en el desarrollo de plantas injertadas (Nieto-Ángel y Borys, 1999). En plántulas de tomate injertadas 'Fanny'/'AR-9704' y evaluadas ocho días después de realizar la injertación por el método de aproximación y con lengüeta, se observó que el xilema y floema en el punto de unión del injerto estaban totalmente fusionados y en plena actividad; además, hubo un incremento de la actividad enzimática del tipo peroxidasa y catalasa, implicadas éstas en la unión del injerto (Fernández-García et al., 2004).
De los tejidos de conducción en los tallos para cualesquier tipo de planta, el tamaño de los vasos del xilema, el porcentaje del xilema y del floema, y la relación entre xilema y floema, son caracteres anatómicos que definen la capacidad de transporte de agua en las plantas, observándose en especies arbóreas que, a medida que el porcentaje de los tejidos vasculares aumenta y el diámetro de los vasos es menor, la cantidad de agua transportada es mayor, y este carácter puede ser un indicador de mayor adaptación de las plantas a condiciones de baja humedad en el suelo (Vasconcellos y Castle, 1994; Reyes-Santamaría et al., 2002).
Se considera que las características anatómicas de los tallos en la unión del injerto son muy importantes para conocer el grado de compatibilidad vegetativa, la capacidad de transporte de agua, la tasa de desarrollo y la capacidad de producción de las plantas en la relación injerto/ portainjerto.
Por lo anterior, y considerando la importancia del uso de portainjertos en tomate, la presente investigación tuvo como objetivo analizar la variación anatómica alrededor del punto de unión del injerto de los tallos de la combinación injerto/portainjertos y viceversa a nivel de plántula, de tres cultivares comerciales y un portainjerto criollo de tomate, como una alternativa para evaluar la compatibilidad vegetativa y su efecto en el transporte de agua y desarrollo de las variedades injertadas.
MATERIALES YMÉTODOS
El estudio se realizó en invernaderos de la Universidad Autónoma Chapingo, en Chapingo, Estado de México (19° 29' N; 98° 53' O), a 2,240 m de altitud.
Se evaluaron tres cultivares comerciales de tomate: 'Reserva F1', 'Charleston F1' y 'Sun 7705', y una población silvestre proveniente de Tegucigalpa, Honduras, que se utilizó como portainjerto. Las plántulas, tanto de los cultivares como del portainjerto, fueron propagadas por semilla, sembrando 15 días antes las semillas del portainjerto con relación a los cultivares.
Se germinaron 40 semillas de cada cultivar, y a los 30 días después de poner las semillas a germinación, las plántulas tuvieron casi 7 cm de altura cuando se utilizaron para la injertación por medio del método de púa en hendidura; del total de plántulas, sólo 30 de ellas se injertaron para evaluar la compatibilidad de injertos, y 10 plántulas no se injertaron y se utilizaron como testigo. La evaluación de la anatomía de los tejidos vasculares alrededor de la unión de los injertos se realizó 15 días después de la injertación, y se utilizaron sólo cinco plántulas de cada cultivar, una por cada repetición; de cada cultivar injertado se obtuvo un tramo de 5 mm de largo de tallo de la unión del injerto, siendo el mismo tamaño para la plántulas no injertadas a la altura de las plántula injertadas, los cuales se fijaron en una solución formaldehído-alchol-ácido acético (50 % etanol 96° + 5 % ácido acético glacial + 10 % formaldehído de 40 % de pureza + 35 % agua); después, se procesaron dichas muestras en un procesador automático de tejidos (Tissuematon Fisher) con etilenglicol monoetil éter (Cellosolve) y xileno, para transferirse a parafina (55 °C) y se sometieron 72 horas dentro de una estufa. Se elaboró el taquete y pirámide de parafina de acuerdo con Sass (1968) y en un micrótomo rotatorio (American Optical modelo 820) se realizaron cortes transversales con grosor de 10 μm, procurando realizar los cortes 2 mm abajo de la unión del injerto, en la unión, y 2 mm arriba de la unión del injerto, colocando las muestras en portaobjetos y cubriéndolos con cubreobjetos, utilizando el adhesivo Haupt y formol al 10 %; su tinción se realizó con safranina y verde fijo para después ser montados (Sass, 1968).
Los cortes anatómicos de tallos fijados en los portaobjetos, se analizaron mediante la utilización de la técnica de Sperry y Saliendra (1994), la que consistió en evaluar cuatro campos planta realizando conteos cada 90° y con una cámara Leica modelo ET-8800N, adaptada a un microscopio también marca Leica modelo Galen III, grabando las imágenes en digital a 40x de objetivo y 10x de ocular; los caracteres evaluados en el punto de unión del injerto fueron: número, tamaño, forma y densidad de células de los tejidos vasculares por campo, y con el programa Image Tool versión 3 (Wilcox et al., 1995), se obtuvieron los datos; así mismo se calcularon el número de vasos por milímetro cuadrado, el diámetro tangencial del vaso (10-2mm), diámetro mayor (10-2 mm), área (10-2 mm-2), perímetro (10-2 mm), índice de redondez [ (4 · π · área)/ perímetro2 ] y el diámetro Feret de los vasos 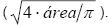 También se calculó la variabilidad intra-grupo
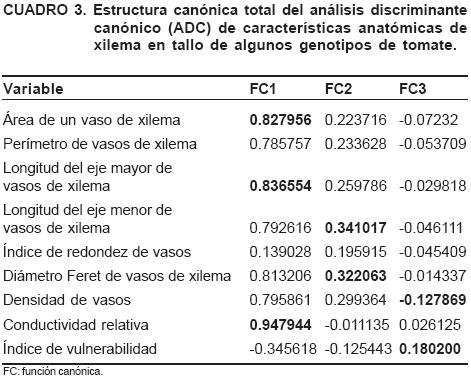
También se calculó la variabilidad intra-grupo 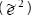 propuesta por Núñez-Colín y Barrientos-Priego (2006) para las variables anatómicas: área, perímetro, longitud de los ejes mayor y menor, diámetro Feret de un vaso del xilema, índice de redondez y densidad vasos del xilema, así como la conductividad relativa e índice de vulnerabilidad del mismo tejido de conducción.
propuesta por Núñez-Colín y Barrientos-Priego (2006) para las variables anatómicas: área, perímetro, longitud de los ejes mayor y menor, diámetro Feret de un vaso del xilema, índice de redondez y densidad vasos del xilema, así como la conductividad relativa e índice de vulnerabilidad del mismo tejido de conducción.
Se obtuvo también la conductividad relativa del brote, mediante la fórmula de Poiseuille modificada por Fahn et al. (1986), CR =(R4 · FRE), donde CR se refiere a la conductividad relativa; R, al radio del vaso y FRE es el número de vasos por milímetro cuadrado. También se calculó el índice de vulnerabilidad de Carlquist (1977),  , donde V se refiere a la vulnerabilidad; DV, al diámetro del vaso y FRE es el número de vasos por milímetro cuadrado. De los datos obtenidos se calcularon los estadísticos básicos para analizar el comportamiento de cada variable dentro de cada grupo derivado de un análisis de componentes principales (ACP), y con esto se analizó la variabilidad intra-grupo , determinando también el centroide del grupo (coordenadas medias de los componentes principales), así como la distancia euclidiana cuadrada de cada individuo con respecto a su centroide, y calcular así el promedio de estas distancias, su error estándar (+ES) con los demás grupos (Núñez-Colín y Barrientos-Priego, 2006).
, donde V se refiere a la vulnerabilidad; DV, al diámetro del vaso y FRE es el número de vasos por milímetro cuadrado. De los datos obtenidos se calcularon los estadísticos básicos para analizar el comportamiento de cada variable dentro de cada grupo derivado de un análisis de componentes principales (ACP), y con esto se analizó la variabilidad intra-grupo , determinando también el centroide del grupo (coordenadas medias de los componentes principales), así como la distancia euclidiana cuadrada de cada individuo con respecto a su centroide, y calcular así el promedio de estas distancias, su error estándar (+ES) con los demás grupos (Núñez-Colín y Barrientos-Priego, 2006).
También se realizó un análisis discriminante canónico (ADC) para comparar los grupos derivados de la función canónica e identificar las variables que explican la variabilidad entre grupos; ésto porque, al utilizar tres cultivares de tomate como injertos y por la reciprocidad entre ellos, el efecto de las variables que conforman cada grupo seguramente será distinto; en el mismo sentido, se calculó la distancia de Mahalanobis para determinar si existen diferencias entre estos grupos (Johnson, 1998). Estos análisis fueron probados en diversos trabajos sobre caracterización de poblaciones vegetales y, en consecuencia, de los caracteres que los determinan, con excelentes resultados (Núñez-Colín et al., 2006; Reyes Santamaría et al., 2002).
RESULTADOS YDISCUSIÓN
Caracteres anatómicos de los vasos del xilema de plántulas injertadas
Considerando que todos los cortes transversales de los tallos de las plántulas se realizaron a los 15 días después de realizar los injertos y a los 5 mm arriba de la unión del injerto (Figura 1), se encontró que el promedio del área, perímetro, longitud de los ejes mayor y menor de los vasos del xilema, los valores fueron superiores (P≤0.05) 'Charleston'/'Criollo de Honduras', 'Sun 7705'/'Criollo de Honduras' y viceversa (Cuadro 1), con relación a 'Charleston' y 'Sun 7705' sin injertar; sin embargo, para el caso del 'Reserva F1' tanto en las dos combinaciones y sin injertar, no semanifestó algún cambio y los valores de estos caracteres fueron iguales (P≤0.05). Es importante indicar que los valores de estos mismos caracteres también fueron superiores (P≤0.05) en las dos combinaciones de 'Charleston' con relación a las de los otros cultivares, coincidiendo en el que también las dos combinaciones de 'Charleston'/'Criollo de Honduras' y 'Criollo de Honduras'/ 'Charleston', presentaron los máximos valores en conductividad relativa, lo que pudiera indicar que este cultivar posee una mayor capacidad de conducción de agua y elementos minerales para el abastecimiento a los diferentes órganos de la planta, como lo citan Vasconcellos y Castle (1994) y Reyes-Santamaría et al. (2002), y a medida que se incremente el porcentaje de los tejidos del xilema se incrementa también la cantidad de agua transportada, por lo tanto existe una mayor adaptación de las plantas a condiciones de baja humedad en el sustrato. Hargrave et al. (1994) encontraron, en plantas de Salvia mellifera, una alta correlación positiva entre el mayor diámetro de los vasos del xilema y la conductividad hidráulica específica del tallo, en tanto que Yang y Tyree (1993) observaron que la conductividad hidráulica absoluta en plantas de maple (Acer saccharum) presentaron una alta correlación con el diámetro basal de la rama en las plantas.
Así mismo, la disminución de la conductividad hidráulica está íntimamente ligada a un mayor estrés hídrico de la planta, lo que supone que ésta debería desarrollar áreas foliares con potenciales hídricos más negativos para lograr la absorción de agua, lo que sería una ventaja que las plantas presentaran diámetros de los tejidos del xilema mayores y, con ésto, favorecer la hidratación de las mismas. El carácter índice de vulnerabilidad, en el que 'Reserva F1' fue más uniforme en tanto que en 'Charleston' y 'Sun 7705', los datos no tuvieron una tendencia clara.
Lo anterior pudiera explicarse por que al realizar los cortes para la injertación, tanto de las células como de los tejidos, al verse interrumpidos los conductos vasculares como efecto de los cortes, tienden a desarrollarse más haciendo que incrementen su tamaño, situación que puede ser favorecido para la reactivación fisiológica normal de la planta, siendo un aspecto fisiológico positivo en el proceso de reconocimiento celular para el "encallamiento" de las células. También es probable que al interrumpirse la continuidad de los vasos del xilema, la traslocación de carbohidratos producidos por las hojas, al no lograr pasar en la unión del injerto, éstos favorecieron el desarrollo mayor de los vasos del xilema; sin embargo, estos datos no son suficientes para saber si este comportamiento anatómico de los vasos se conserven a lo largo del tallo principal de la parte injertada al portainjerto de tomate.
Motisi et al. (2004) observaron que en injertos de plantas de nectarina (Prunus persica) sobre portainjertos de bajo vigor de la misma especie, ocurrió una reducción en cuanto a la conductividad del agua; sin embargo, cuando los portainjertos fueron vigorosos la conductividad se mantuvo constante y similar entre el portainjerto y el injerto; al parecer, esto mismo ocurrió en el presente estudio, en el que seguramente el portainjerto indujo una mayor área de los tejidos de conducción, debido probablemente a que las plantas de dicho portainjerto era más vigoroso.
Variación interna de los vasos del xilema
De las mediciones de los vasos del xilema, primero se realizó el análisis de componentes principales (ACP), en el que se utilizaron sólo los tres primeros componentes, mismos que explican 68.24, 23.22 y 6.38 %, respectivamente, acumulando el 97.84 % de la variación total.
Se tuvieron los siguientes resultados (Figura 2). El cv. Sun 7705, al utilizarlo como injerto, fue el que presentó la mayor varianza interna, lo que significa que presenta una alta heterogeneidad en su sistema de conducción, por lo que las plantas pueden presentar características favorables a excesos de agua y otras a sequía (Reyes-Santamaría et al., 2002; Núñez-Colín et al., 2006), por lo menos con los cultivares aquí evaluados. Caso contrario es el cv. Reserva F1, el que presentó mayor homogeneidad, lo que significa que este cultivar es el que más fácilmente puede adaptarse en cuanto a requerimientos hídricos, puesto que la mayoría de las plantas de este genotipo presentan las mismas características.
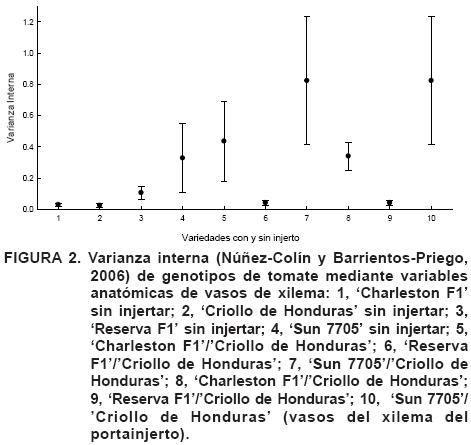
Otro aspecto importante es que criollos sin injertar, criollos como portainjerto de 'Sun 7705' y 'Reserva F1', presentan una menor heterogeneidad que el que fue utilizado como portainjerto de 'Charleston', pudiendo significar que al funcionar como portainjerto este cultivar presenta una alta plasticidad genética que permite adaptarse a los cultivares injertados. El único caso en el que su comportamiento fue distinto, es 'Sun 7705', puesto que su portainjerto presentó una alta homogeneidad y el injerto, alta heterogeneidad, en los otros dos casos los valores de (variabilidad intra-grupo) fueron similares.
Análisis Discriminante Canónico
En este análisis se utilizaron las tres primeras funciones o raíces canónicas que explican el 89.78 % de la variación total (Cuadro 2).
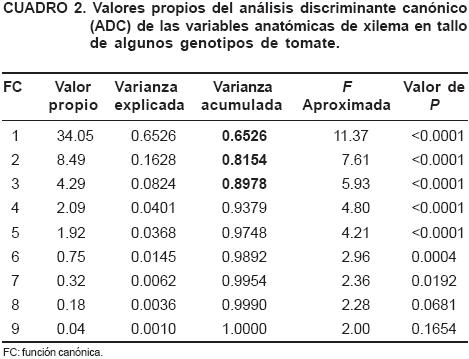
La primera función canónica explica el 65.26 % de la variabilidad total y está asociada a la longitud del eje mayor promedio, al área promedio de los vasos de xilema y a la conductividad relativa, lo que significa que posiblemente la variabilidad concentrada en esta primera función puede estar asociada a la cantidad de agua que se pudiera transportar por el tamaño de vasos, ya que presentan el mismo signo en los coeficientes en la estructura canónica y están altamente correlacionadas (Cuadro 3); a mayor tamaño de vasos, mayor conductividad relativa, ambas variables pueden dar una aproximación del agua que requiere cada cultivar. La segunda función canónica explica el 16.28 % de la variabilidad total, estando asociada al diámetro Feret y a la longitud del eje menor de los vasos del xilema, no encontrando alguna relación biológica lógica con el comportamiento de estas variables. La tercera función explica sólo el 8.24 % de la variabilidad total, y está asociada a la densidad de vasos y al índice de vulnerabilidad, y generalmente plantas con un mayor número de vasos relativamente pequeños, están asociados a genotipos resistentes a sequía (Núñez-Colín et al., 2006; Reyes-Santamaría et al., 2002; Vascocellos y Castle, 1994).
En la proyección tridimensional (Figura 3) se observó que el genotipo criollo, utilizado como portainjerto en cada uno de los cultivares y sin injertar, presentó características muy diferentes, lo que explicaría que este genotipo presenta una amplia plasticidad anatómica derivado de su carácter genético, ya que al ser un genotipo que no ha sufrido una presión de selección, la amplitud de adaptación a las condiciones ambientales también es mayor (Dovzhansky, 1970).
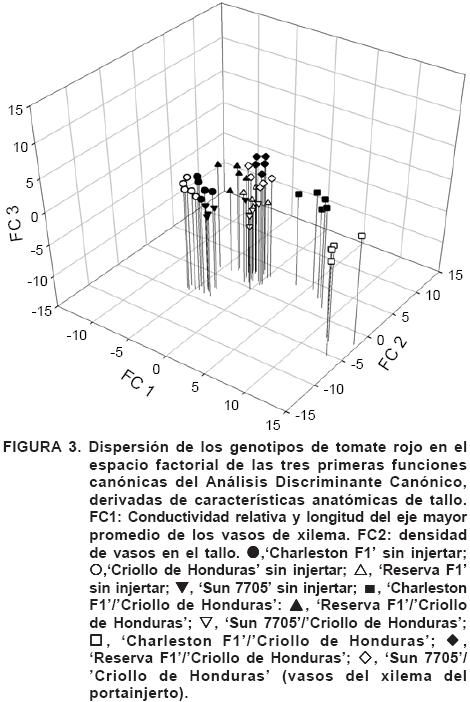
El 'Criollo de Honduras' expande su tamaño y densidad de vasos de acuerdo con el cultivar que se le injerte; así el 'Criollo de Honduras' que fue injertado con 'Charleston F1', presentó mayor tamaño de vasos, pero menor densidad. 'Sun 7705' y su portainjerto presenta diámetros de vasos similares, aunque el cultivar, en este caso, presentó una mayor densidad de los mismos. Para el caso de 'Reserva F1' y su portainjerto, el cultivar presentó un menor tamaño de vasos y una menor densidad con relación a su portainjerto, y estas muestras también divergen del material criollo sin injertar, por lo que se comprueba la plasticidad genética que presenta dicho material (Dovzhansky, 1970).
Características de los vasos del xilema de las plántulas injertadas versus plántulas no injertadas
Los valores de las mismas variables analizadas anteriormente, al comparar las plántulas injertadas contra las no injertadas, indican que cuando los valores de t son positivos (Cuadro 4), significa que los valores son mayores para los tratamientos sin injertar, en tanto que si los valores son negativos, entonces los valores son mejores para los tratamientos en donde se realizó el injerto. Por lo anterior, los datos mostraron que para las variables área, perímetro, longitud del eje mayor y menor de vasos del xilema y variables derivadas de éstas, se encontró que fue significativamente mayor el tamaño de los vasos del xilema al injertarlas con el portainjerto 'Criollo de Honduras', lo que significa que fue mejor realizar los injertos en 'Charleston F1' y 'Sun 7705', no así para 'Reseva F1', que sus valores no presentaron un nivel estadísticamente significativo, es decir, sin cambios al realizar los injertos. También se incrementaron los valores de las mismas variables en los portainjertos de las variedades injertadas, y altamente significativos en 'Charleston F1' y 'Reserva F1', y sólo significativamente mejor en 'Sun 7705'.
No hubo diferencias significativas entre los valores de las variedades sin injertar versus portainjertos sin injertar, y sí mayor efecto en los portainjertos injertados en relación a los portainjertos no injertados (Cuadro 4).
En general podemos indicar que los valores se incrementaron, tanto en las variedades cultivadas como en los portainjertos al tener la relación injerto/portainjerto.
CONCLUSIONES
La anatomía de los vasos del xilema de los cultivares de tomate injertados y de portainjerto criollo, presentaron cambios con respecto al testigo sin injertar.
El área, perímetro, longitud del eje mayor y menor de los vasos del xilema, fueron estadísticamente superiores al realizar los injertos, en 'Charleston F1' y 'Sun 7705' sin injertar.
En el caso del cv. Reserva F1, tanto en las dos combinaciones y sin injertar, no manifestó algún cambio y los valores de estos caracteres fueron estadísticamente iguales.
El cv. Charleston F1 presentó, en los mismos caracteres referidos, los máximos valores en relación a los otros cultivares, además de que en las dos combinaciones del cv. Charleston F1 en relación a las combinaciones, 'Charleston F1'/'Criollo de Honduras' y 'Criollo de Honduras'/ 'Charleston F1', también fueron superiores a los otros cultivares.
En las dos combinaciones de injertos del cv. Charleston F1 se presentaron los máximos valores en el carácter conductividad relativa en relación a los otros cultivares.
Los valores de las variables: área, perímetro, longitud del eje mayor y menor de los vasos del xilema y las variables derivadas de éstas, se encontró que fue significativamente mayor el tamaño de los vasos del xilema al injertarlos con el portainjerto 'Criollo de Honduras', lo que significa que fue mejor realizar los injertos en 'Charleston F1' y 'Sun 7705', no así para 'Reseva F1'.
El cv. Sun 7705 al utilizarlo como injerto fue el que presentó la mayor varianza interna, y esta heterogeneidad en su sistema de conducción significan características favorables a excesos de agua y otras a sequía, caso contrario es el cv. Reserva F1, que presentó mayor homogeneidad lo que significa que este cultivar es el que más fácilmente puede adaptarse en cuanto a requerimientos hídricos, puesto que la mayoría de las plantas de este genotipo presentan las mismas características, corroborándose esto con los datos de las funciones canónicas en el que el área y la longitud del vaso del xilema, así como la conductividad relativa son los caracteres con mayor tamaño, lo que pudiera deducirse que exista una mayor posibilidad de transporte de agua; en el mismo sentido se observó que el genotipo criollo como portainjerto presentó características de mayor plasticidad anatómica en los vasos del xilema, lo que pudiera proponerse que el criollo pudiera presentar una mayor amplitud de adaptación a diversas condiciones ambientales.
Al comparar los valores, área, perímetro, longitud del eje mayor y menor de los vasos del xilema y las otras variables derivadas de éstas, se observa que es significativamente mayor en las variables del portainjerto 'Criollo de Honduras' en relación a los cultivares sin injertar, y sólo significativamente en los portainjertos de los cultivares injertados; es decir, los valores se incrementaron, tanto en los cultivares como en los portainjertos al tener la relación injerto/portainjerto.
LITERATURA CITADA
ANÓNIMO. 2005. Servicio de Información Agroalimentaria y Pesquera (SIAP) de la Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación (SAGARPA). [ Links ]
ANÓNIMO. 1998. El jitomate, la hortaliza de excelencia en exportación. Claridades Agropecuarias 62: 3-18. [ Links ]
AYRE, B. G.; TURGEON, R. 2004. Graft transmission of a floral stimulant derived from constans. Plant Physiology 135: 2271-2278. [ Links ]
BLACK, L. L.; WU, D. L.; WANG, J. F.; KALB, T.; ABBASS, D.; CHEN, J. H. 2003. Grafting Tomatoes for Production in the Hot-Wet Season. Asian Vegetable Research & Development Center. AVRDC Publication Num. 03-551. 6 p. [ Links ]
CARDOSO, S. C.; FERMINO SOARES, A. C.; DOS SANTOS BRITO, A.; ARAUJO DE CARBALHO, L.; DA SILVA LEDO, C. A. 2006. Viabilidade de uso do híbrido Hawai 7996 como portaenxerto de cultivares comerciais de tomate. Bragantia (São Paulo), Campinas 65(1): 89-96. [ Links ]
CARLQUIST, S. 1977. Ecological factors in wood evolution: a floristic approach. American Journal of Botany 64: 887-896. [ Links ]
DOBZHANSKY, T. 1970 Genetics of the evolutionary process. Columbia University Press. Columbia, USA. 505 p. [ Links ]
FAHN, A.; WERKER, E.; BAAS, P. 1986. Wood anatomy and identification of trees and shrubs from Israel and adjacent regions. Academy of Sciences and Humanities. Jerusalem, Israel 221 p. [ Links ]
FERNÁNDEZ-GARCÍA, N.; CARVAJAL, M.; OLMOS, E. 2004. Graft union formation in tomato plantas: peroxidase and catalase involvent. Annals of Botany 93: 53-60. [ Links ]
HARGRAVE, K. R.; KOLB, K. J.; EWERS, F. W.; DAVIS, S. D. 1994. Conduit diameter and drought-induced embolism in Salvia mellifera Greene (Labiatae). New Phytologist 126: 695 705. [ Links ]
HOLBROOK, N. M.; SHASHIDHAR, V. R.; JAMES, R. A.; MUNNS, R. 2002. Stomatal control in tomato with ABA-deficient roots: response of grafted plants to soil drying. Journal of Experimental Botany 53(373): 1503-1514. [ Links ]
JOHNSON, D. E. 1998. Métodos Multivariados Aplicados al Análisis de Datos. Traducido por H. Pérez Castellanos. International Thomson Editores. D. F. , México. 566 p. [ Links ]
KACJAN MARŠI', N.; OSVALD, J. 2004. The influence of grafting on yield of two tomato cultivars (Lycopersicon esculentum Mill.) grow in a plastic house. Acta Agriculturae Slovenica 83(2): 243-249. [ Links ]
LIFSCHITZ, E.; EVIATAR, T; ROZMAN, A.; SHALIT,A.; GOLDSHMIDT, A.; AMSELLEM, Z.; ALVAREZ, J. P.; ESHED, Y. 2006. Substitute for diverse environmental stimuli ortholog triggers systemic signals that regulate growth and flowering and FT The tomato. Proceedings of the National Academy of Sciences 103(16): 6398-6403. [ Links ]
MITIDIERI, M. S.; BRAMBILA, M. V.; PIRIS. M.; MALDONADO, L. 2005. El uso de portainjertos resistentes en cultivo de tomate bajo cubierta: resultados sobre la sanidad y rendimiento del cultivo. Instituto Nacional de Tecnología Agropecuaria, Centro Regional Buenos Aires Norte, Estación Experimental Agropecuaria San Pedro. Buenos Aires Argentina. pp. 1-8. [ Links ]
MOTISI, A.; CARUSO, T.; GRUTTA, I.; MARRA, F. P.; PERNICE, F. 2004. Canopy architecture appraisal by fractal dimension of 'flordastar' peach grafted onto different rootstocks. Acta Horticulturae 732: 383-389. [ Links ]
NIETO-ÁNGEL, R.; BORYS, M. W. 1999. Relaciones fisiológicas y morfológicas de injertos de frutales sobre tejocote (Crataegus spp.) como portainjerto. Revista Chapingo Serie Horticultura 5(2): 137-150. [ Links ]
NÚÑEZ-COLÍN, C. A.; BARRIENTOS-PRIEGO, A. F. 2006. Estimación de la variabilidad interna de muestras poblacionales, mediante el análisis de componentes principales. Interciencia 31(11): 802-806. [ Links ]
NÚÑEZ-COLÍN, C. A.; BARRIENTOS-PRIEGO, A. F.; RODRÍGUEZ-PÉREZ, J. E.; NIETO-ÁNGEL, R. 2006. Variabilidad anatómica de los sistemas de conducción y estomático de genotipos de Prunus spp. de diferentes orígenes. Pesquisa Agropecuaria Brasileira 41(2): 233-241. [ Links ]
PARKINSON, M.; YEOMAN, M. M. 1982. Graft formation in cultured, explanted internodes. New Phytologist 91: 711-719. [ Links ]
PEIL, R. M. N.; GÁLVEZ, J. L. 2004. Rendimiento de plantas de tomate injertadas y efecto de la densidad de tallas en el sistema hidropónico. Hort. Bras. 22 (2): 265-270. [ Links ]
REYES-SANTAMARÍA, I.; TERRAZAS, T.; BARRIENTOS-PRIEGO, A. F.; TREJO, C. 2002 Xylem conductivity and vulnerability in cultivars and races of avocado. Scientia Horticulturae 92: 97-105. [ Links ]
RUIZ, J. M.; BLASCO, B.; RIVERO, R.; ROMERO, L. 2005. Nicotine-free and salt-tolerant tobacco plants obtained by grafting to salinity-resistant rootstocks of tomato. Physiologia Plantarum 124: 465–475. [ Links ]
SASS, J. E. 1968.Botanical Microtechnique. 3rd Edition. The Iowa State University Press. Ames, Iowa, USA. 227 p. [ Links ]
SPERRY, J. S.; SALIENDRA, N. Z. 1994. Intra and inter-plant variation in xylem cavitation in Betula occidentalis. Plant Cell and Environment 17: 1233-1241. [ Links ]
VASCONCELLOS, L. A. B. C.; CASTLE, W. S. 1994. Trunk xylem anatomy of mature healthy and blighted grapefruit trees on several rootstocks. Journal of American Society for Horticultural Science 119(2): 185-194. [ Links ]
WILCOX, D; DOVE, B.; MC.DAVID, D.; GREER, D. 1995. Image Tool. Version 3 Users guide.University of Texas, Health Science Center, San Antonio, USA. [ Links ]
YANG, S.; TYREE, M. T. 1993. Hydraulic resistance in Acer saccharum shoots and its influence on leaf water potential and transpiration. Tree Physiology 12(3): 231-242. [ Links ]

