










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Chapingo. Serie horticultura
On-line version ISSN 2007-4034Print version ISSN 1027-152X
Rev. Chapingo Ser.Hortic vol.15 n.3 Chapingo Sep./Dec. 2009
Densidad de población en cultivo hidropónico para la producción de tubérculo-semilla de papa (Solanum tuberosum L.)
Population density in hydroponics culture for potato tuber-seed production (Solanum tuberosum L.)
R. Flores-López¶; F. Sánchez-del Castillo; J. E. Rodríguez-Pérez; M. T. Colinas-León; R. Mora-Aguilar; H. Lozoya-Saldaña
Instituto de Horticultura. Departamento de Fitotecnia. Universidad Autónoma Chapingo. Km. 38.5 Carretera México-Texcoco. Chapingo, Estado México. C. P. 56230. MÉXICO. Correo-e: floresrmx@yahoo.com.mx (¶Autor responsable).
Recibido: 19 de septiembre, 2008.
Aceptado: 7 de julio, 2009.
Resumen
El propósito del presente estudio fue determinar la influencia de la densidad de siembra (45, 25, 16 y 8 plantas·m-2), sobre la producción de minitubérculos mayores de 10 mm de diámetro e indicadores fisiológicos del crecimiento en condiciones de invernadero e hidroponia en la variedad Gigant. En plantas crecidas a mayor densidad de población (45 plantas·m2) el índice de área foliar fue de 13.4 a los 75 días después de la emergencia (DDE) y la altura de la planta se incrementó 45 %, respecto a la menor densidad (8 plantas·m2), con esta última, la biomasa total (67.5 g), el número (16.5) y biomasa (52.5 g) de tubérculos por planta fueron significativamente mayores a los 75 DDE, en comparación con la densidad de 45 plantas·m2, con 44.5 g, 10.2 g y 36.8 g, respectivamente. La tasa absoluta de crecimiento (TAC) de la planta completa varió de 0.4 a 1.7 g·d-1 dependiendo de la densidad y de la etapa de desarrollo del cultivo, mientras la TAC de tubérculo a menor densidad fue 45 a 59 % superior respecto al tratamiento de 45 plantas·m-2, la eficiencia fotosintética y la partición de carbohidratos al tubérculo, medidos como tasa de asimilación neta (TAN) y tasa de asimilación económica neta (TAEN), fueron 118 a 160 % mayores en la densidad de 8 plantas·m2 respecto a 45 plantas·m2, sin embargo, la tasa de crecimiento del cultivo (TCC) a la menor densidad representó el 12 % del valor del tratamiento de mayor densidad (64.4 g·m-2·d-1) a los 75 DDE, lo que se reflejó, en esta última, en el número y biomasa de tubérculos mayores de 10 mm de diámetro, con 470 unidades y 1,658 g·m-2 respectivamente.
Palabras clave adicionales: Solanum tuberosum, tubérculo-semilla, invernadero, hidroponia.
Abstract
The aim of this work was to study the influence of plant density (45, 25, 16 and 8 plants·m-2) on hydroponic greenhouse production of Gigant variety potato minitubers larger than 10 mm in diameter and on physiological growth indicators. With high population density (45 plants·m-2) the leaf area index was 13.4 at 75 days after emergence and plant height was 45 % higher than plants grown at 8 plants·m-2. At this density, the total plant biomass (67.5 g), tuber number (16.5) and tuber biomass (52.5 g) were significantly higher than at 45 plants·m-2, with 44.5 g, 10.2 and 36.8 g, respectively.
The absolute growth rate (AGR) of whole plant varied from 0.4 to 0.7 g·d-1 depending on plant density and the growth stage, and the absolute tuber growth rate (TGR) in the lower density treatment was 45 to 59 % higher than the TGR of plants growing at 45 plants·m-2, Photosynthetic efficiency and carbohydrates diverted to the tuber, estimated as net assimilation rate (NAR) and economic assimilation rate (EAR), with 8 plants·m-2 were 118 to 160 % higher than the NAR and EAR with 45 plants·m-2. However, the crop growth rate (CGR) with 8 plants·m-2 was only 12 % of the value obtained with 45 plants·m-2 (64.4 gm-2·d-1) at 75 DAE. This was also manifested in a larger number of tubers over 10 mm in diameter and tuber biomass (470 units and 1,658 g·m2, respectively).
Additional keywords: Solanum tuberosum, tuber-seed, hydroponics, greenhouse.
INTRODUCCIÓN
La producción en invernadero de semilla básica de papa (Solanum tuberosum L.) se realiza con el propósito de obtener semilla libre de enfermedades, que muestre todo su potencial de producción en campo, tanto para semilla, como para la producción comercial de este tubérculo, por ello se requieren tubérculos mayores de 10 mm de diámetro que ya puedan ser cultivados en campo, aseguren la emergencia y mayor cantidad de tallos por metro lineal para así lograr el establecimiento adecuado del cultivo y la mayor productividad del mismo.
El proceso de tuberización es influenciado por factores ambientales e intrínsecos; entre los que destacan el genotipo, la concentración interna de AG3, fotoperiodo, temperatura y radiación interceptada por la planta. Algunos de los factores que pueden ser manipulados para incrementar la producción de minitubérculos en invernadero son: momento de inicio de fertilización, solución nutritiva y su composición, densidad de plantación, arreglo espacial de las plantas y diámetro mínimo de los tubérculos a ser cosechados (Struik y Wiersema, 1999).
El número de tubérculos producidos por planta decrece gradualmente al aumentar la densidad de plantación (Rubio et al., 2000), lo que conduce a bajas tasas de multiplicación, por lo que el número de tubérculos producidos varía de dos a diez, en función de la técnica de producción empleada (Ranally, 1997). La densidad óptima de plantación depende de la variedad, del número deseado de minitubérculos y de los costos de producción, entre los que destacan el costo del material inicial; minitubérculos o plantas in vitro, espacio de invernadero y labores culturales (Struik y Wiersema, 1999).
En cultivos hidropónicos con el uso de esferas de arcilla expandida Rolot y Seutin, (1999) indican produjeron más de 400 tubérculos·m-2, tanto con 59 como con 200 plantas·m-2, debido a la disposición continúa de nutrientes. Por otro lado, en aeroponia 60 plantas·m-2 produjeron 802 tubérculos·m-2, el doble de los obtenidos con 100 plantas·m-2 (Farran y Mingo-Castel, 2006). En sustratos como tierra de monte, mezclas en diferentes proporciones de fibra de coco:perlita y turba:perlita y fertilización de N, P y K granulado se recomiendan densidades de población en invernadero de entre 44 y 100 plantas·m-2 (Rubio et al., 2000).
Existe escasa información disponible referente a la influencia de la densidad de población en la producción de minitubérculos en condiciones de cultivo hidropónico; conocimiento requerido en sistemas de producción donde no hay restricción de nutrientes y el ambiente subterráneo se puede controlar a favor del desarrollo de la planta. Estudios de acumulación de materia seca (Mora et al., 2005) y el uso de indicadores fisiológicos permiten describir cuantitativamente el crecimiento mediante muestreos periódicos de biomasa de la planta completa, sus partes, y del área foliar; han mostrado su eficacia para entender la influencia de factores intrínsecos y extrínsecos involucrados en la producción de papa (Basu et al., 1999); así, se han hecho estudios sobre: evaluación de genotipos (Aguilar et al., 2006; Mora et al., 2006), humedad del suelo (Jefferies y Mackerron, 1989) y determinación de la eficiencia de nutrientes (McCollum, 1978).
El propósito de la presente investigación fue estudiar la influencia de la densidad de siembra en cultivo hidropónico con perlita bajo invernadero sobre indicadores fisiológicos del crecimiento y el rendimiento de minitubérculos.
MATERIALES Y MÉTODOS
El experimento se estableció en Chapingo, Estado de México, de mayo a agosto de 2006 en un invernadero de cristal con las siguientes condiciones ambientales: humedad relativa máxima de 88%, mínima de 36 % y media de 62 %; temperatura media de 21 °C y una mínima nocturna de 7 °C, la radiación varió a lo largo del día de acuerdo con las condiciones ambientales, con una lectura máxima de 2000 i moles·m-2·s-1.
Se empleó la variedad Gigant, liberada en Holanda en 1987, actualmente cultivada en México, de ciclo precoz (80 a 90 días), piel blanca, ojos superficiales, pulpa crema, forma oblonga-aplanada, susceptible al tizón tardío de la papa (Phytophthora infestans Mont. de Bary) y con calidad para consumo sin procesar (Rubio et al., 2000). Los minitubérculos utilizados fueron de 15 a 17 mm de diámetro, libres de enfermedades de virus, hongos y bacterias.
El experimento se realizó en condiciones hidropónicas en macetas de plástico negro con volumen de un litro rellenas con perlita grado hortícola como sustrato. En cada maceta se depositó un minitubérculo a 10 cm de profundidad y las plantas se condujeron a un solo tallo. La solución nutritiva empleada tuvo la siguiente concentración de nutrimentos en mg·litro-1: N =160, P = 80, K = 250, Ca = 250, Mg = 75, Fe =4, Mn = 0.5, Cu = 0.5, Zn = 0.5, B = 0.5, con pH de 6.0 y conductividad eléctrica final de 2.0 dS·m-1. El gasto diario de la solución nutritiva suministrada mediante riego por goteo fue de 500 ml por planta, dosificada en cuatro riegos (9:30, 12:30, 14:30 y 16:30 h). A lo largo del ciclo se hicieron aplicaciones preventivas con Mancozeb a razón de 2 g·litro-1 y Clorotalonil 1.5 ml·litro-1 para el control de tizón tardío de la papa, se alternaron aplicaciones de Metamidofos, Endosulfan y Ometoato una vez por semana, para el control de mosquita blanca (Bemisia tabaci Genn).
Se utilizó el diseño experimental de bloques completos al azar, los tratamientos aplicados consistieron en cuatro densidades de siembra: 45, 25, 16 y 8 plantas·m-2 con cuatro repeticiones, la unidad experimental consistió de 25 macetas por tratamiento.
Los caracteres medidos a los 30, 40, 50, 60, 70 y 75 días después de la emergencia fueron: altura de la planta (cm), peso seco de estolones (g), peso seco de tubérculos (g), peso seco de vástago (g), peso seco de la planta completa (g), número de tubérculos y área foliar (cm2) medida con un integrador LI-COR, modelo LI-3100. El peso seco de los distintos órganos se obtuvo al secar en estufa a 70 °C durante 72 horas y los tubérculos se seccionaron y se secaron por 96 horas.
El análisis de crecimiento de las plantas se realizó con los datos de materia seca de tubérculo, vástago y planta completa en los muestreos realizados en las fechas indicadas, los datos fueron ajustados mediante regresión no lineal, con el modelo logístico: 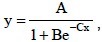 donde: Y fue la variable respuesta; X = la variable independiente; A = la asíntota, valor de Y cuando X tiende a infinito; B fue el parámetro relacionado con la ordenada al origen; C = el valor de X para el punto de inflexión; e (base de los logaritmos neperianos) = 2.718281828.
donde: Y fue la variable respuesta; X = la variable independiente; A = la asíntota, valor de Y cuando X tiende a infinito; B fue el parámetro relacionado con la ordenada al origen; C = el valor de X para el punto de inflexión; e (base de los logaritmos neperianos) = 2.718281828.
Se calculó la tasa absoluta de crecimiento (TAC), que corresponde a la primera derivada del modelo logístico (Rodríguez et al., 2006).
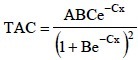
Ésta se define como la ganancia en gramos de materia seca por día, en este caso del vástago, tubérculos y de la planta completa.
La tasa de asimilación neta (TAN) que representa la cantidad de biomasa acumulada por unidad de área foliar y unidad de tiempo (g·m-2·d-1), se calculó de acuerdo con la fórmula siguiente:

Donde: TAC es la tasa absoluta de crecimiento de la planta; af = área foliar.
La tasa de asimilación económica neta (TAEN) representa la cantidad de biomasa del órgano de interés por unidad de área foliar y unidad de tiempo (g·m-2·d-1). Es análoga a la TAN, sólo que se toma en cuenta el peso seco de tubérculo (McCollum, 1978).

Donde TCT = tasa absoluta de crecimiento de tubérculo.
La tasa de crecimiento del cultivo (TCC) se expresa en términos de peso por unidad de superficie y tiempo, se obtuvo al multiplicar el TCC = IAF.TAN.
TCC = IAF.TAN
Donde, IAF = índice de área foliar; TAN = tasa de asimilación neta.
El cálculo del índice de área foliar (IAF) se hizo al dividir el área foliar total de la planta entre la superficie de suelo ocupada por la misma, considerando los pasillos.
El índice de cosecha (IC) se obtuvo al dividir la biomasa de los tubérculos entre la biomasa total acumulada por la planta hasta el momento de la cosecha.
RESULTADOS Y DISCUSIÓN
Al incrementar la densidad de población, aumentó la altura de la planta (Cuadro 1), con diferencia (P = 0.05) entre tratamientos desde los 30 DDE y hasta los 75 DDE. Plantas del tratamiento de menor densidad (8 plantas·m-2) crecieron 31 % menos que las plantas del tratamiento de 45 plantas·m-2; lo anterior fue ocasionado posiblemente por la interacción hormonal, ya que en condiciones de sombra el ácido indolacético se incrementa y actúa sinérgicamente con las giberelinas, lo que provoca elongación celular (Gardner et al., 1990). En hojas de plantas expuestas a sombreo Woolley y Waering (1972) detectaron alta concentración de giberelinas, y se ha comprobado alta concentración endógena de éstas en tallos en condiciones no favorables para la tuberización (Menzel, 1985; Vreugdenhil y Sergeeva, 1999).

A los 30 DDE el área foliar por planta fue mayor significativamente en los tratamientos de mayor densidad (25 y 45 plantas·m-2) en comparación con los tratamientos de 16 y 8 plantas·m-2. El IAF fue superior (P=0.05) en el dosel de plantas de la mayor densidad con 7.4 (Cuadro 1), en comparación con el resto de los tratamientos, y esta diferencia se mantuvo durante todo el ciclo de cultivo. El incremento del índice de área foliar maximiza la intercepción de radiación en el dosel en los primeros días, hasta alcanzar índices de área foliar donde hay sombreo entre las plantas, de tal manera que un IAF de 4.0 intercepta aproximadamente 95 % de la radiación incidente (Boyd et al., 2002).
Para la determinación de la eficiencia fotosintética de las plantas a través del análisis de crecimiento, los valores de acumulación de materia seca de tubérculo, vástago y total fueron linearizados mediante el modelo logístico con R2 mayor de 0.8; los parámetros A, B y C (Cuadro 2) se emplearon para calcular los indicadores fisiológicos de crecimiento: TAC, TAN, TAEN y TCC discutidos a continuación.
La tasa absoluta de crecimiento del vástago (Figura 1) desde los 30 a los 50 DDE en las plantas de mayor densidad descendió, mientras que en el resto de los tratamientos se incrementó ligeramente, manteniendo este comportamiento hasta los 75 DDE, en donde los máximos valores correspondieron a la densidad de 16 plantas·m-2 (0.6 g·d-1) y el menor se obtuvo con 45 plantas·m-2 (0.06 g·d-1).
Lo anterior indica que las plantas crecidas bajo 45 plantas·m-2 compiten más e interceptan menos radiación, por lo que la planta crece en longitud del tallo pero con poca acumulación de materia seca. En los cuatro tratamientos la TAC de tubérculo se incrementó hasta los 50 DDE donde las plantas crecidas a menor densidad presentaron un crecimiento exponencial de tubérculo por día con máxima acumulación de materia seca de los tubérculos de 1.66 g·d-1, también las plantas de los tratamientos de 16 y 25 plantas·m-2 tuvieron su máxima acumulación de materia seca a los 50 DDE, con valores de 0.8 y 0.9 g·d-1 de máxima ganancia, respectivamente, lo que representa el 50 % respecto al tratamiento de 8 plantas·m-2. En contraste el tratamiento de 45 plantas·m-2 presentó su máxima tasa de acumulación de materia seca en tubérculo a los 75 DDE, lo que indica que el ciclo de cultivo se alargó debido posiblemente a las condiciones no inductivas de tuberización ocasionada por la menor captación de radiación por planta.
Lo anterior sugiere que con altas densidades de población la disposición de radiación por planta disminuye, lo que ocasiona menor acumulación de materia seca en el órgano de interés; situación que indica alteración de las relaciones fuente-demanda, ya que se promueve el crecimiento de la parte aérea a expensas del crecimiento de los tubérculos (Gawronska et al., 1990).
A los 50 DDE sólo el tratamiento de 8 plantas·m-2 alcanzó su máxima TAC total por planta (1.7 g·d-1), mientras que para densidades de 16 y 25 plantas·m-2 los valores máximos se presentaron de los 57 a los 65 DDE (1.3 y 1.1 g·d-1, respectivamente). Por su parte, plantas del tratamiento de 45 plantas·m-2, alcanzaron su máxima acumulación de materia seca a los 75 DDE (1.4 g·d-1). El comportamiento de la TAC de la planta completa está influenciado por la TAC de los tubérculos en todos los tratamientos, lo que concuerda con lo detectado por Mora et al. (2006), al estudiar la cinética de acumulación de biomasa de cinco genotipos de papa.
A los 30 DDE la TAN de plantas crecidas a menor densidad ya era de 6.1 g·m-2d-1; 160 % superior que la TAN con 45 plantas·m-2 y 35 % respecto a la de plantas de 16 y 25 plantas·m-2. De los 40 a 50 DDE la TAN de plantas a menor densidad llegó a su valor más alto (8.6 g·m-2·d-1); superior 82, 93 y 118 % respecto a la TAN de las plantas con 16, 25 y 45 plantas·m-2. Después de los 50 DDE y hasta los 75 DDE la TAN de las plantas desarrolladas a densidades de población de 8, 16 y 25 plantas·m-2 descendió hasta llegar a 2.9, 3.6 y 2.2 g·m-2·d-1, respectivamente; en contraste con las plantas crecidas a mayor densidad de población (45 plantas·m-2), donde la TAN se incrementó y hasta los 75 DDE alcanzó la mayor ganancia de biomasa seca por unidad de área foliar por día (3.9 g·m-2·d-1). Lo anterior sugiere que la reducción de la radiación provocada por la alta densidad de población en plantas de papa de la variedad Gigant conduce al alargamiento del ciclo de cultivo y confirma que las plantas con mayor disponibilidad de luz fijan mayor cantidad de CO2 por unidad de tiempo (Figura 1).
Por otro lado, la TAN máxima en plantas de cada tratamiento coincide con el periodo de mayor TAC de tubérculo, lo que sugiere que el incremento en la demanda de asimilados en los tubérculos estimula la tasa de fotosíntesis (Aguilar et al., 2006; Mora et al., 2006).
Así mismo, la TAEN presentó un comportamiento similar a la TAN, la mayor tasa de acumulación de biomasa de tubérculos por planta y más rápido en el ciclo fue en plantas a menor densidad, lo que ocurrió también con 25 y 16 plantas·m-2 con el máximo a los 50 días para descender a menos de 1 g·m-2·d-1 a los 75 DDE, mientras a mayor densidad la eficiencia de asimilación económica se incrementó lentamente de 1.2 a los 30 DDE a 3.1 g·m-2·d-1 a los 75 DDE (Figura 1). Lo anterior indica que a menor densidad, la planta acorta su ciclo de cultivo y produce más asimilados que son destinados al tubérculo.
A los 75 DDE la biomasa de estolones por planta con 45 plantas·m-2 disminuyó con respecto al resto de las densidades empleadas (P=0.05); sin embargo, el número y biomasa de tubérculos por planta (Cuadro 3), fue igual con respecto a los tratamientos de 16 y 25 plantas·m-2, pero no en comparación con plantas del tratamiento de 8 plantas·m-2, las cuales presentaron los valores más altos de biomasa y número de tubérculos (26 y 37 %, respectivamente). Esta menor acumulación de biomasa y rendimiento de tubérculos en los tratamientos de mayor densidad posiblemente se debió al autosombreo de las plantas. Se ha demostrado que bajos niveles de radiación disminuyen la asimilación de CO2, biomasa total y rendimiento en tubérculos (Gawronska et al., 1990; Wurr et al., 1997).
Sin embargo, a los 75 DDE el índice de cosecha fue igual estadísticamente (P=0.05) en tres de las cuatro densidades (16, 25 y 45 plantas·m-2) lo que implica que al final del ciclo de cultivo, independientemente de estas tres densidades empleadas, las plantas destinaron la misma proporción de asimilados al tubérculo; en contraste, las plantas del tratamiento de menor densidad tuvieron el índice de cosecha significativamente mayor que el de los tres tratamientos anteriores, lo que confirma que a mayor intercepción de radiación fotosintéticamente activa por planta no sólo se producen mayor cantidad de asimilados, sino que también se destina mayor proporción de carbohidratos al tubérculo, y la diferencia encontrada en biomasa de tubérculo es debida tanto a la mayor cantidad de asimilados producidos como a la partición de mayor proporción de los mismos hacia los tubérculos en plantas del tratamiento de menor densidad. Estos resultados concuerdan con lo mencionado por Muro et al. (1997) quienes informan que a mayor densidad de población no incrementó el número de tubérculos de menor diámetro en detrimento de los tubérculos de mayor diámetro, lo que puede ser debido a que las densidades empleadas (20, 41 y 83 plantas·m-2) fueron menores que el máximo potencial que puede soportar el cultivo hidropónico.
El análisis de los resultados por unidad de superficie de invernadero, muestran resultados opuestos a los obtenidos por planta. La tasa de crecimiento del cultivo (TCC) a los 30 DDE fue más baja en el tratamiento de 8 plantas·m-2 (6.1 g·m-2·d-1) que representó el 31, 39 y 61 % de la TCC de los tratamientos de 45, 25 y 16 plantas·m-2 (Figura 1). En los cuatro tratamientos la TCC se incrementó conforme se desarrolló el cultivo y a los 50 DDE la producción de materia seca por unidad de superficie de plantas crecidas a menor densidad de población representó el 40, 49 y 78 % de aquellas de 45, 25 y 16 plantas·m-2, respectivamente. Y a los 75 DDE la producción de materia seca por unidad de superficie y tiempo en el tratamiento de 8 plantas·m-2 fue de 8.1 g·m-2·d-1, que representó sólo 12 % de la TCC del tratamiento de mayor densidad de población con 64.4 g·m 2·d-1 y el 38 y 35 % respecto a 16 y 25 plantas·m-2 con 21.3 y 23.2 g·m-2·d-1, respectivamente. Lo anterior indica que a mayor densidad de población la fijación de CO2 por unidad de superficie de invernadero se incrementó, debido al desarrollo más rápido de área foliar por el dosel (Boyd et al., 2002). El mayor índice de área foliar lo tuvo el tratamiento de 45 plantas·m-2 desde los 30 DDE y hasta los 75 DDE. Lo anterior fue confirmado con los resultados obtenidos al multiplicar los valores medios de número y biomasa de tubérculos mayores de 10 mm por planta por el número de plantas por metro cuadrado presentes en cada tratamiento. El número total de minitubérculos en el tratamiento de 45 plantas·m-2 fue de 470 unidades, similar a lo indicado por Rolot et al. (1999) quienes empleando esferas de arcilla y 200 plantas·m-2 obtuvieron más de 400 tubérculos·m-2, mientras que el menor número de tubérculos·m-2 (132) fue detectado en las plantas del tratamiento de menor densidad (8 plantas·m-2). Respecto a la materia seca acumulada en los tubérculos se obtuvieron 420 g·m-2 con 8 plantas·m-2, en contraste con 1,658 g·m-2 para 45 plantas·m-2, casi cuatro veces más que en la menor densidad de población empleada.
CONCLUSIONES
La alta densidad de población (45 plantas·m-2) en cultivo hidropónico en invernadero con la variedad Gigant, disminuyó la acumulación de biomasa total, biomasa y número de tubérculos mayores de 10 mm de diámetro por planta, la tasa absoluta de crecimiento, la tasa de asimilación neta y la tasa económica de asimilación neta por planta, en contraste incrementó la tasa de crecimiento del cultivo, así como la biomasa y número de tubérculos mayores de 10 mm, por unidad de superficie de invernadero respecto al tratamiento de menor densidad de población (8 plantas·m-2).
LITERATURA CITADA
AGUILAR, L. M. G.; CARRILLO, S. A. J.; RIVERA, P. A.; GONZÁLEZ, H. A. V. 2006. Análisis de crecimiento y de relaciones fuente-demanda en dos variedades de papa (Solanum tuberosum L.). Rev. Fitotecnia Mexicana 29(2): 145-156. [ Links ]
BOYD, N. S.; GORDON, R.; MARTIN, R. C. 2002. Relationship between leaf area index and ground cover in potato under different management conditions. Potato Research 45: 117-129. [ Links ]
BASU, P. S.; ASHOO, S.; GARG, I. D.; SUKUMARAN, N. P. 1999. Tuber sink modifies photosynthetic response in potato under water stress. Enviromental and Experimental Botany 42(1): 25-39. [ Links ]
FARRAN, I.; MINGO-CASTEL, M. 2006. Potato minituber production using aeroponics: Effect of plant density and harvesting intervals. American Journal of Potato Research 83: 47-53. [ Links ]
GARDNER, P. F.; PEARCE, B. R.; MITCHELL, L. R. 1990. Physiology of Crop Plants. Iowa State University Press: AMES. USA. 327 p. [ Links ]
GAWRONSKA, H.; DWELLE, B. R.: PAVEK, J. J. 1990 Partitioning og photoassimilates by potato plants (Solanum tuberosum L.) as influenced by irradiance: II. Partitioning patterns by four clones grown under high and low irradiation. American Potato Journal 67: 163-176. [ Links ]
JEFFERIES, A. R.; MACKERRON, L. K. D. 1989. Radiation interception and growth of irrigated and droughted potato (Solanum tuberosum). Field Crops Research 22: 101-112. [ Links ]
MCCOLLUM, E. R. 1978. Analysis of potato growth under differing P regimes. II. Time by P-status interactions for growth and leaf efficiency. Agronomy Journal 70: 58-67. [ Links ]
MENZEL, C. M. 1985. Tuberization in potato at high temperatures: Interaction between temperature and irradiance. Annals of Botany 55: 35-39. [ Links ]
MIDMORE, D. J.; PRANGE, R. K. 1992. Growth responses of two Solanum species to contrasting temperatures and irradiance levels: relations to photosynthesis, dark respiration and chlorophyll fluorescence. Annals of Botany 69(1): 13-20. [ Links ]
MORA, A. R.; ORTIZ, C. J.; RIVERA, P. A.; MENDOZA, C. C. A.; COLINAS, L. M. T.; LOZOYA, S. H.; RODRÍGUEZ, P. J. E. 2005. Comportamiento de la acumulación y distribución de biomasa. Revista Chapingo Serie Horticultura 11(1): 135-142. [ Links ]
MORA, A. R.; ORTIZ, C. J.; RIVERA, P. A.; MENDOZA, C. C. A.; COLINAS, L. M. T.; LOZOYA, S.H. 2006. Índices de eficiencia de genotipos de papa establecidos en condiciones de secano. Revista Chapingo Serie Horticultura 12(1): 85-94. [ Links ]
MURO, J.; DÍAZ, V.; GOÑI, J. L.; LAMSFUS, C. 1997. Comparison of hydroponic culture and culture in peat/sand mixture and the influence of nutrient solution and plant density on seed potato yields. Potato Research 40: 431-438. [ Links ]
RANALLI, P. 1997. Innovative propagation methods in seed tuber multiplication programmes. Potato Research 40: 439-453. [ Links ]
RODRÍGUEZ, P. J. E.; SAHAGÚN, C. J.; GASGA, P.; MORA, A. R. 2006. Programas de cómputo para la estimación de parámetros de regresión no lineal útiles en el análisis de crecimiento de cultivos. 52da Reunión Anual de la Sociedad Interamericana de Horticultura Tropical. Sep. 24-30. San Juan, Puerto Rico. [ Links ]
ROLOT, L. J.; SEUTIN, H. 1999. Soiless production of potato minitubers using a hydroponic technique. Potato Research 42: 457-459. [ Links ]
RUBIO, C. O. A.; RANGEL G. J.; FLORES, L. R.; MAGALLANES, G. J. V.; DÍAZ, H. C.; ZAVALA, Q. T.; RIVERA P. A.; CADENA, H. M.; ROCHA, R. R.; ORTIZ, T. C.; LÓPEZ, D. H.; DÍAZ, V. M.; PAREDES, T. A. 2000. Manual para la producción de papa en las sierras y valles altos del centro de México. SAGAR. INIFAP. CIRCE. Campo Experimental Valle de Toluca. Libro Técnico No. 1. Zinacantepec, Estado de México.79 p. [ Links ]
STRUIK, C. P.; WIERSEMA, G. S. 1999. Seed Potato Technology. Ed. Wageningen Pers, Wageningen, The Netherlands 383. [ Links ]
WOOLEY D. J.; WAEREING, P. F. 1972. Environmental effects on endogenous cytokinins and gibberellin levels in Solanum tuberosum. New Phytology 71: 1015-1025. [ Links ]
WURR, D. C. E.; HOLE, C. C.; FELLOWS, J. R.; MILLING, J.; LYNN, J. R.; O'BRIEN, P. J. 1997. The effect of some enviromental factors on potato tuber numbers. Potato Research 40: 297-306. [ Links ]
VREUGDENHIL, D.; SERGEEVA, I. L. 1999. Gibberellins and tuberization in potato. Potato Research 42: 471-481. [ Links ]

