










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Chapingo. Serie horticultura
On-line version ISSN 2007-4034Print version ISSN 1027-152X
Rev. Chapingo Ser.Hortic vol.15 spe Chapingo Jan. 2009
Identificación de algunas especies de microorganismos benéficos en la rizosfera de gerbera y su efecto en la productividad
Inoculation responses with benefical microorganisms in gerbera taking into account rizosphera caracterization
M. R. Soroa–Bell1*, A. Hernández–Fernández2, F. Soto–Carreño1 y E. Terry–Alfonso1
1Departamento de Fitotecnia, Instituto Nacional de Ciencias Agrícolas. Carretera Tapaste. Km 3½. San José de las Lajas. La Habana, Cuba. C. P. 32700. Correo–e: resobell@inca.edu.cu (*Autor responsable).
2Departamento de Microbiología, Universidad de La Habana. Calle 25 entre H e I. Vedado. Ciudad de La Habana, Cuba.
Recibido: 29 de abril, 2009
Aceptado: 21 de septiembre, 2009
Resumen
Se identificaron las especies de hongos micorrízicos arbusculares y los géneros de bacterias promotoras del crecimiento vegetal atraídos por los exudados radicales de Gerbera jamesonii cv. Bolus. Para el aislamiento de las bacterias promotoras del crecimiento vegetal, se utilizó el modelo espermosférico, los tubos colectores y la cámara de quimioatracción modificada. Para la caracterización de las cepas, se realizaron determinaciones micromorfológicas (movilidad, presencia de endosporas, entre otras) y culturales (forma, tamaño, opacidad, entre otras) en los medios estudiados. Se realizaron cinco repeticiones por tratamiento y se replicaron estas determinaciones tres veces. Se encontraron cinco especies de hongos micorrízicos arbusculares y cuatro géneros de bacterias promotoras del crecimiento vegetal atraídos por los exudados de gerbera. La especie Glomus hoi like, tuvo 10 esporas·g–1, mientras que por las bacterias promotoras del crecimiento vegetal. el género Pseudomonas representó el 30% de los microorganismos presentes en la rizosfera. A partir de lo obtenido se inocularon plantas adultas establecidas en recipientes de cinco litros de capacidad de suelo ferralítico rojo más estiércol vacuno a una proporción de 2:1. Se evaluó el rendimiento y diámetro del capítulo floral, encontrándose que las plantas inoculadas con Glomus hoi like obtuvieron mayor ocupación fúngica en la raíz (1.91%), mayores poblaciones de hongos (5.6 esporas·ml–1) y bacterias (8.2 log UFC·ml–1) y mayor número de flores (35).
Palabras clave: Gerbera jamesonii cv Bolus, microorganismos, hongos micorrízicos arbusculares, bacterias promotoras del crecimiento vegetal, biofertilización, rizosfera.
Abstract
We identify the mycorrizal fungi (HMA) species living in the rhizosphere of Gerbera jamesonii cv. Bolus and the plant growth promoting bacterial (PGBP) genera attracted by those radical exudates. The chemoattraction of some plant growth promoting bacterial toward root exudates of this crop was also evaluated. The espermospher model was used for bacterial isolation and Collector tubes and the chemotaxis camera modified for the evaluation of the chemotactic effect of those radical exudates. To the bacterial characterization, was carried out micromorphological determinations (mobility, endosporas presence and others) and cultural determinations (forms, size, opacity and others). Five replications per treatment were used three times. Five micorrizal fungi species and four plant growth promoting bacterial genera attracted by gerbera radical exudates were found, almost Glomus genus were predominant, being Glomus hoi like specie the principal one. Pseudomonas (30%) was the predominant genus among bacterials. Taking into account these results, adult plants were established in recipients of five litres of capacity, were inoculated using red ferralitic soil mixed with cow dung in a proportion of 2:1. The evaluated variables were yield and floral capitulum diameter and root colonization, finding that inoculated plants with Glomus hoi like had more root occupation (1.91%), more mycorrizal fungus espores (5.6 espores·ml–1) and bacterial (8.2 log UFC·ml–1) in the rhizosphere and more flowers (35).
Key words: Gerbera jamesonii cv. Bolus, mycorryzal fungus, promoting growth plant bacterial, biofertilization, rhizosphere and microorganisms.
INTRODUCCIÓN
Un número amplio de microorganismos se encuentran en el suelo. La diversidad y el número de los mismos dependen en gran medida de la composición y concentración de los nutrientes exudados por las raíces de las plantas (Wamberg et al., 2003). La interacción entre los microorganismos y los cultivos puede ser beneficiosa, dañina o neutral, en ocasiones esto puede variar en función de las condiciones del suelo (Lynch, 1990).
Conocer los integrantes de la comunidad microbiana asociada al cultivo de interés es un aspecto de particular atención para desarrollar una floricultura ecológica ya que es posible favorecer la aplicación de inoculantes sin perjudicar el equilibrio biológico de los suelos. También, el análisis del comportamiento de los hongos y bacterias ante los exudados vegetales, constituye uno de los principios básicos de la interacción planta – microorganismo. Los exudados radicales son utilizados por los microorganismos como fuente nutritiva, influyendo indirectamente en las interrelaciones entre los microorganismos colonizadores a través de la acción selectiva que ejercen sobre especies o grupos particulares.
Pocos han sido los estudios encaminados a identificar los microorganismos atraídos por los exudados radicales en especies de flor de corte, la mayoría de los reportes relacionados con estos estudios han sido realizados en gramíneas, fundamentalmente en cereales y los microorganismos encontrados en dichos cultivos difieren a partir de la zona geográfica, tipo de suelo y cultivo, entre otros aspectos (Hernández et al., 2008).
La gerbera (Gerbera jamesonii cv Bolus), es cultivada en Cuba gracias a su plasticidad en cuanto a tipo de suelo y régimen térmico. Es una especie vistosa que cuenta con una alta demanda internacional, pero durante su cultivo se aplican excesivamente insumos químicos que alteran la comunidad microbiana y disminuye la presencia de insectos benéficos (Altieri, 2007).
La agricultura actual propone el uso de biofertilizantes (Terry, 2006) que, en los sistemas productivos es una alternativa para lograr un desarrollo agrícola ecológicamente sostenible. Fernández et al. (2006) plantean adoptar una estrategia de suministro de nutrientes a las plantas mediante la combinación de fertilizantes minerales con abonos orgánicos y biofertilizantes, poniendo énfasis en los últimos por su bajo costo, no contaminan el ambiente y mantienen la fertilidad y biodiversidad; por lo que, integrar el estudio de la biofertilización es de vital importancia en el cultivo de gerbera y constituye una alternativa de manejo en el entorno urbano y periurbano.
Este trabajo tuvo como objetivo la identificación y evaluación de los hongos micorrízicos arbusculares y bacterias promotoras del crecimiento vegetal asociadas a la rizosfera de Gerbera jamesonii cv Bolus, para introducir la biofertilización y determinar su repercusión en la productividad del cultivo.
MATERIALES Y MÉTODOS
Para el desarrollo de la fase experimental se concibieron dos etapas: la primera en la que se identifican los hongos micorrízicos y las bacterias promotoras del crecimiento vegetal asociados a la rizosfera de Gerbera; la segunda donde se inoculan las plantas con los microorganismos mencionados y se evalúa el rendimiento y diámetro de las inflorescencias.
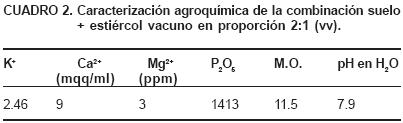
El suelo sobre el cual se establecieron las plantas de gerbera (Gerbera jamesonii cv Bolus), fue Ferralítico Rojo lixiviado éutrico (Cuba, MINAG, 1999). El Cuadro 1 muestra las características del mismo. Las plantas para el estudio de caracterización fueron fertilizadas durante todo su cultivo con estiércol vacuno.
Se identificó la comunidad bacteriana asociada al cultivo, para lo que se utilizaron semillas botánicas obtenidas por productores especializados en la reproducción de esta especie. Las mismas fueron previamente desinfectadas con solución de hipoclorito de calcio al 10% y procesadas, según metodología descrita por Hernández et al. (2008) y se colocaron en cámaras húmedas, hasta su germinación. Posteriormente, estas semillas se colocaron en tubos espermosféricos sin fuente de carbono y nitrógeno con el objetivo de aislar las bacterias con la capacidad de vivir a expensas de los exudados radiculares del cultivo. Finalmente, cada uno de los tubos espermosféricos fue inoculado con diluciones seriadas (10–1 a 10–6) de las diferentes muestras de suelo rizosférico procedente de las plantas en estudio. Se establecieron cinco tubos por dilución y una planta por cada tubo. Se incubaron en un fotoperíodo de 16 horas luz – ocho horas oscuridad a 27 °C, durante siete días.
Las raíces de las plantas crecidas en cada tubo se extrajeron y maceraron. Se realizaron diluciones seriadas (10–1 a 10–6) en solución salina compuesta por NaCl, KCL, KH2PO4 y Na2HPO4 y se sembraron en medios selectivos para los géneros microbianos en estudio (Hernández y Escalona, 2003). Para la identificación de las cepas, se realizaron determinaciones micromorfológicas (reacción de gram, movilidad, presencia de endosporas, cápsulas, flagelos, etc.) y culturales (forma, tamaño, opacidad, elevación, superficie, borde, consistencia, etc.) en los medios estudiados. Se realizaron cinco réplicas por tratamientos y se replicaron estas determinaciones tres veces para autentificar las cepas encontradas.
A partir de las especies detectadas en la rizosfera de gerbera, se seleccionaron los hongos micorrízicos: Glomus hoi like y G. moseeae y las bacterias: Psedomonas fluorescens y Burkholderia cepacia, los cuales constituyen inóculos de probada infectividad y efectividad en el manejo de diferentes cultivos agrícolas (Rivera et al., 2003). Estas especies sirvieron, para evaluar el comportamiento de Gerbera jamesonii cv. Bolus al ser inoculada con los referidos microorganismos en su medio rizosférico. En este experimento se utilizaron plantas adultas de gerbera que fueron multiplicadas agámicamente y que no habían sido fertilizadas con anterioridad. Se establecieron las plantas en recipientes de cinco litros de capacidad y el sustrato natural empleado fue suelo más estiércol vacuno composteado (Cuadro 2), a una proporción de 2:1 (v:v).
El efecto de la inoculación simple con HMA y PGPB sobre el desarrollo y productividad de Gerbera se valoró a partir del empleo de los siguientes tratamientos:
1. Burkholderia cepacia (Cepa: 0057– Título 3.24x109 ufc·g de turba–1).
2. Pseudomonas fluorescens (Cepa: J–143 – Título 2.3x109 ufc g·de turba–1).
3. Glomus hoi like (Cepa: INCAM–2 – Título 250 esporas·g de suelo–1). Especie abundante, muy parecida a Glomus hoi Berch &Trappe.
4. Azospirillum brasilense (Cepa: Sp–7 – Título 3.55x109 ufc·g de turba–1)
5. Glomus mosseae (Cepa: INCAM–4 – Título 250 esporas·g de suelo–1)
6. Testigo de producción (Fertilizante mineral, formulación 9–13–17)
Se empleó como soporte sólido la turba molinada, tamizada y estéril. La inoculación se realizó por el método de recubrimiento en el momento de la plantación (Gómez et al., 1996 citado por Fernández et al., 2006). Para todos los casos se mezcló un kilogramo del inóculo más 600 ml de agua con lo que se obtuvo una mezcla pastosa que sirvió para recubrir la zona radicular de las plantas una vez podadas las raíces y un tercio de las hojas.
Diseño experimental y análisis estadístico:
Para determinar en la primera etapa la presencia de hongo micorrízico, número de esporas y otros grupos microbianos se tomaron 50 g de suelo rizosférico en 10 plantas establecidas, seleccionadas al azar y cultivadas sin biofertilizantes ni fertilización mineral, a partir de las cuales se conformó y procesó la muestra de rizosfera. El conteo de hongos se realizó por el método de disolución–suspensión de suelo, con siembras superficiales en placas petri (Morell et al., 2006).
Para determinar el efecto de la inoculación simple con HMA y PGPB sobre el desarrollo y productividad de gerbera, se empleó un diseño Completamente Aleatorizado con seis tratamientos. Las evaluaciones se realizaron a los 60 días de establecidas las plantas, a razón de 10 plantas por tratamientos. Se midieron las siguientes variables: diámetro de flores (cm), número de flores por plantas y colonización de hongos micorrízicos (Phyllips y Hayman, 1970).
Para el procesamiento de datos se empleó un Análisis de Varianza de Clasificación Simple y se utilizó la Prueba de Rangos Múltiples de Duncan, cuando existieron diferencias entre los tratamientos, utilizando el programa estadístico STATGRAPHICS Versión 5.1 Plus para Windows.
RESULTADOS Y DISCUSIÓN
• Hongos micorrízicos arbusculares (HMA) en la rizosfera.
Se encontraron cinco especies predominantes en la rizosfera de gerbera que fueron: Glomus hoi like, G. intraradices like, G . sp., G. mosseae like y Scutellospora sp.
De estas especies asociadas a la rizosfera, G. hoi like fue la que predominó pues se observó un total de 10.47 esporas·g–1 (Cuadro 3). Este promedio difirió significativamente del promedio de las otras especies, quienes tuvieron un menor número de esporas por gramo de suelo. Cuatro de las cinco especies detectadas pertenecen el género Glomus que, como se sabe (Rivera y Fernández, 2003), son altamente infectivos, por lo tanto el manejo de ellos se puede comenzar con cualquiera de los propágulos infectivos del hongo micorrizógeno, ya sea esporas, raicillas infectadas o fragmentos de hifas; y estos inóculos, aún sin esporas mantienen un elevado potencial infectivo (Rivera y Fernández, 2003). Generalmente las especies de este género han sido reportadas como eficientes en diferentes tipos de suelos, mientras que, en el caso de Scutellospora sp., sólo las esporas parecen ser los propágulos infectivos, como en las especies de Gigaspora.

La cuantificación de especies en suelo ferralítico rojo lixiviado con diferentes niveles de explotación, indicaron que el género Glomus predomina en la rizosfera de cultivos permanentes como mango, ficus y en áreas de explotación intensiva (Morell et al., 2006).
La comunidad de especies de HMA es específica de un sitio y depende a su vez de las prácticas agronómicas (Sieverding, 1991), por lo que el conocimiento de las especies de HMA presentes y su manejo, ya sea elevando las concentraciones de los propágulos encontrados o cambiando en el tiempo la composición de las mismas, es importante para un adecuado manejo del agroecosistema.
Los HMA son considerados insumos biológicos de enorme potencial en la agricultura, debido a sus efectos positivos sobre la adaptación y el crecimiento de una gran variedad de cultivos. Además, estos microorganismos son componentes clave para el desarrollo de la biota del suelo por su gran capacidad de interacción con diferentes especies microbianas, a la vez que, pueden modificar muchos aspectos de las propiedades físicas en la zona rizosférica (Riera et al., 2005). Todos esos efectos modifican los patrones de colonización de la raíz micorrizada, donde se desarrollan procesos biológicos que mejoran las condiciones de los suelos para el desarrollo de las plantas, aspectos muy importantes para el establecimiento de una agricultura sostenible y el funcionamiento del ecosistema.
Muchas interacciones tienen lugar entre los propios hongos micorrízicos y los demás microorganismos de la rizosfera y las respuestas de las plantas a la micorrización involucran no sólo al hongo, sino a todos los micro y macroorganismos presentes por ello es importante conocer además los géneros de bacterias promotoras del crecimiento vegetal presentes.
• Bacterias promotoras del crecimiento vegetal presentes en la rizosfera de Gerbera
La composición de la comunidad bacteriana de Gerbera (Figura 1), evidenció que Pseudomonas constituye una población dominante con relación a Azospirillum, Bacillus y Streptomyces; alcanzando el 30% del total de esporas.
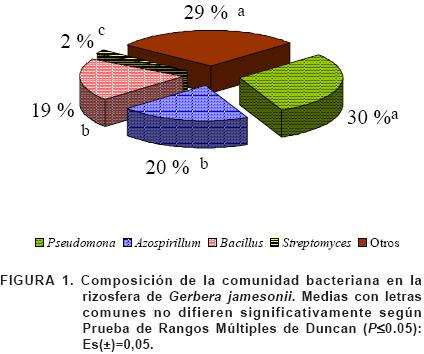
Pseudomonas constituye un género microbiano abundante en la rizosfera de cultivos de interés agrícola, fundamentalmente por su capacidad de metabolizar una amplia gama de sustancias carbonadas exudadas por las raíces de las plantas, además de su metabolismo versátil, corto período de latencia y rápida velocidad de crecimiento (Loredo et al., 2004 y Hernández et al., 2008). Asimismo, Hernández et al. (2008), al caracterizar la rizosfera del cultivo del maíz var. Francisco mejorado en tres localidades edafoclimáticas diferentes, reportó que Pseudomonas constituye un género dominante.
Otros de los géneros observados aunque en menor cuantía fueron Azospirillum y Bacillus. Bacterias del género Azospirillum han sido aisladas de distintas regiones geográficas a partir de una gran variedad de plantas pertenecientes a diferentes familias botánicas. Se ha encontrado dominando la rizosfera del cultivo del arroz en suelo ferralítico rojo lixiviado y ferralítico amarillento (Rives et al., 2007).
En cuanto al género Bacillus, Hernández et al. (2004) coinciden en afirmar que es un género predominante en las condiciones estudiadas. Las poblaciones de Streptomyces se encuentran alrededor del 2%.
Este estudio identificó los grupos microbianos benéficos existentes en la rizosfera de gerbera e indica la posibilidad de trabajar con inoculantes que incrementen las poblaciones de los mismos. De esta manera se evitaría la introducción de géneros exógenos en los ecosistemas naturales.
• Efecto de inóculos microbianos en la población de microorganismos de la rizosfera de Gerbera jamesonii
Las concentraciones de hongos y bacterias encontradas en la rizosfera de Gerbera (Figura 2) indican que en esta especie hay una actividad microbiana pues en todas las muestras analizadas, ambos grupos se encontraron presentes. Estos resultados reafirman que bacterias y hongos son los principales constituyentes de la biomasa microbiana del suelo (Baath et al., 2003). La presencia de PGPB puede influir en el crecimiento de las hifas extrarradiculares del HMA y en la colonización de las raíces de las plantas, de ahí la importancia de este estudio.
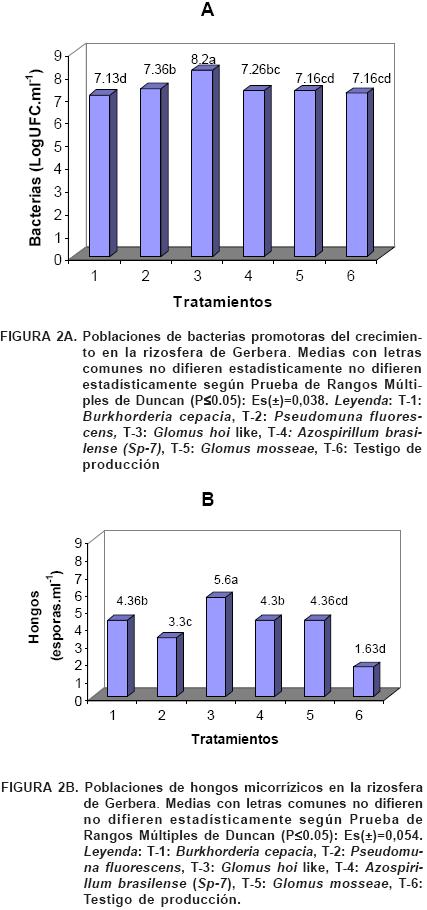
Las plantas inoculadas con Glomus hoi like (Tratamiento 3) presentaron mayor concentración de bacterias (8.2 LogUFC·ml–1) y hongos (5.6 esporas·ml–1) en la rizosfera, lo que remarca la importancia del empleo de los biofertilizantes como una vía para incrementar las poblaciones dominantes en los cultivos agrícolas (Riera y Medina, 2005).
Existen trabajos que estudian las relaciones existentes entre los microorganismos promotores del crecimiento vegetal (hongos micorrízicos arbusculares y las bacterias promotoras del crecimiento vegetal), los cuales desempeñan un papel clave en la toma de nutrientes, la tolerancia al estrés ambiental y, en general, el mantenimiento de la salud radical, favoreciendo así el aumento del rendimiento de los cultivos (Matiru y Dakora, 2004). La micorriza tiene efectos inhibitorios en organismos del suelo patógenos de raíces, dicha inhibición pudieran ser causada por la producción de metabolitos secundarios por los HMA que afectan selectivamente a otros organismos, pero la investigación en este sentido es aún limitada (Wamberg et al., 2003).
Somers y Vanderleyden (2004), clasificaron a los HMA y las PGPB como benéficas ya que tienen la capacidad de colonizar las raíces de las plantas y estimular el crecimiento vegetal, sin provocar afectaciones al ambiente rizosférico.
• Análisis de la colonización radical en plantas inoculadas
Se registró que los HMA tuvieron una colonización por encima del 50% en las raíces de Gerbera (Cuadro 4). Los tratamientos Glomus hoi like y Glomus mosseae alcanzaron una colonización radical del 58%, difiriendo significativamente del resto de los tratamientos. Resultados similares fueron obtenidos por Terry y Leyva (2006) en tomate, donde obtuvo porcentajes de colonización superiores al 50%.
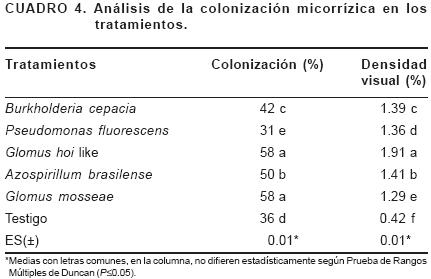
Se encontró que el tratamiento inoculado con G. hoi like ocupa una mayor área en la raíz de la planta y difiere significamente de los tratamientos, alcanzando una densidad visual del 1.9%, lo que pudiera emplearse como criterio para afirmar que este inóculo tuvo una mayor colonización de manera general.
Los valores de colonización encontrados en los tratamientos inoculados con PGPB sugieren que los microorganismos no viven de manera aislada en la rizosfera de las plantas. El tratamiento inoculado con A. brasilense mostró altos valores de colonización; al respecto Mirabal et al. (2002) detectaron bacterias diazotróficas pertenecientes al género Azospirillum en micelios de HMA en suelos tropicales y, podría suponerse que se estableció una relación mutualista positiva entre A. brasilense y los HMA presentes en la rizosfera de esta especie; lo que constituye una herramienta potencial para el desarrollo de una agricultura en armonía con el ambiente.
• Influencia de la inoculación con PGPB y HMA en el diámetro de la inflorescencia y el rendimiento.
La identificación de algunos microorganismos asociados a la rizosfera de Gerbera jamesonii, fue el primer paso para explorar el manejo del cultivo por la escasez de información que existía al respecto. A partir de las especies seleccionadas, se analizó la influencia de la inoculación en el diámetro de la inflorescencia y el rendimiento.
Al estudiar la influencia que ejerció la inoculación con PGPB y HMA en el diámetro promedio de flores (Figura 3), se observó que los tratamientos con G. hoi like y A. brasilense presentaron los valores más altos de este indicador. Sin embargo, no difieren significativamente del tratamiento testigo.

Los resultados indican que, a pesar de que el género Glomus predomina en la rizosfera no todas las especies responden igual. G. hoi like resultó beneficioso para producir diámetros superiores (8.7 cm), sin diferir significativamente de Azospirillum brasilense (8.5 cm), lo que se corresponde, con los resultados obtenidos por Wang et al. (1993) al inocular con HMA diferentes especies ornamentales, entre las cuales se destacaba Gerbera.
Respecto al comportamiento de Azospirillum, Bashan y Bashan (2005) plantearon que se manifiesta como un microorganismo endógeno pero, además, se encontró representando en un 20% a la comunidad bacteriana asociada a Gerbera jamesonii cv Bolus, por lo que al potenciar dicho microorganismo con el inóculo eficiente, se lograron diámetros superiores de inflorescencia.
A pesar de que Pseudomonas es un género fuertemente atraído por los exudados radiculares de Gerbera, los tratamientos con B. cepacia y P. fluorescens no manifestaron los mayores incrementos en el diámetro de las inflorescencias, lo que podría sugerir que no son las especies predominante en gerbera, pudiendo tratarse de P. aureginosa (especie tipo) o P. putida. Por otro lado, la cepa P. fluorescens (J–104), es autóctona de gramíneas y no del cultivo estudiado, por tanto, podría suponerse que los exudados radiculares no ejercieron una quimioatracción activa sobre la misma y no lograra colonizar eficientemente el cultivo. Esto puede haber provocado que en la competencia microbiana, la cepa aplicada como inoculante, no lograra establecerse y colonizar eficientemente el cultivo; de ahí la importancia de trabajar con cepas autóctonas de los cultivos, para garantizar la factibilidad ecológica de los productos que se apliquen.
Por otro lado, las micorrizas son endosimbiontes obligados de las plantas y dependen del carbono que liberan las células radiculares. Dicha presencia altera la exudación radicular y por lo tanto influye en las poblaciones microbianas presentes (Wamberg et al., 2003).
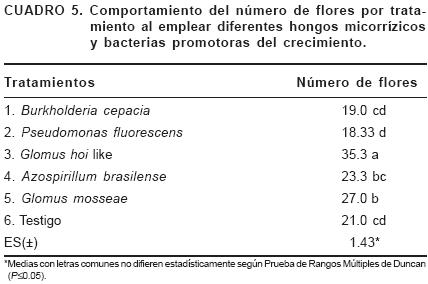
En cuanto al rendimiento por plantas (Cuadro 5), el tratamiento inoculado con G. hoi like aportó la mayor cantidad de flores (35), difiriendo significativamente de los restantes tratamientos. Estos valores corresponden a la producción acumulada hasta los 60 días de efectuada la plantación. González y Ferrera (2001) al inocular con HMA diferentes especies de plantas ornamentales, encontraron los mayores rendimientos en los tratamientos micorrizados.
CONCLUSIONES
Los géneros Glomus y Pseudomonas predominan en la rizosfera de este cultivo; sin embargo, al inocular con Burkholderia cepacia y Pseudomonas fluorescens no se obtuvieron incrementos de las variables diámetro de la inflorescencia y número de flores. El empleo de G. hoi like, A. brasilense y el testigo incrementó el diámetro de las inflorescencias, pero la inoculación con G. hoi like, proporcionó las mayores concentraciones de hongos y bacterias en la rizosfera, mayor infección micorrízica y un rendimiento superior con relación a los restantes inóculos empleados.
LITERATURA CITADA
ALTIERI, M. 2007. La agroecología como alternativa sostenible frente al modelo de agricultura industrial. Agroecología y agricultura ecológica. 229: 75–93. [ Links ]
BAATH, E.; ANDERSON, T. H. 2003. Comparision of soil fungal/baterial ratios in a pH gradient using physiological and PLFA based techniques. Soil Biology and Biochemistry 35: 955–963. [ Links ]
BASHAN, Y.; BASHAN L. E. 2005. Bacterial Plant growth – promotion. Encyclopedia of Soil in the Environment. D. Hillet (Ed.) Elsevier, Oxford, UK. vol 1. 2200 p. [ Links ]
CUBA, MINAG. INSTITUTO DE SUELOS. 1999. Nueva versión de clasificación genética de los suelos de Cuba. La Habana: Agrinfor. 64 p. [ Links ]
FERNÁNDEZ, F.; DELL'AMICO, J. M; RODRÍGUEZ, P. 2006. Efectividad de algunos tipos de inoculantes micorrízicos a base de Glomus hoi–like en el cultivo del tomate (Lycopersicon esculentum Mill. var Amalia). Cultivos Tropicales 27(3): 25–30. [ Links ]
GONZÁLEZ, M. C.; FERRERA, C. 2001. Influencia de cinco hongos endomicorrízicos en el desarrollo del Anthurium. Revista Chapingo Serie Horticultura 1(1): 119–123. [ Links ]
HERNÁNDEZ, A.; HEYDRICH M.; ACEBO, Y.: VELÁSQUEZ, M. G.; HERNÁNDEZ , A. N. 2008. Antagonistic activity of Cuban native rhizobacteria against Fusarium verticillioides (Sacc) Nirenb. in maize (Zea mays L.). Applied Soil Ecology 39: 180–186 [ Links ]
HERNÁNDEZ, A.; RIVES, N.; HEYDRICH, M. 2004. Caracterización de la comunidad microbiana y endófita asociada al cultivo del arroz variedad J–104. En: Congreso Científico del INCA (14:2004, La Habana). Memorias [CD–ROM].Instituto Nacional de Ciencias Agrícolas. [ Links ]
HERNÁNDEZ, L. G.; ESCALONA, M. A. 2003. Microorganismos que benefician a las plantas: las bacterias PGPR. La Ciencia y el Hombre 16: 19–25. [ Links ]
LOREDO, O.; LÓPEZ, L.; ESPÌNOSA, D. 2004. Bacterias promotoras del crecimiento vegetal asociadas con gramínea: Una revisión. Terra Latinoaméricana 22: 225–239. [ Links ]
LYNCH, J. M. 1990. The rhizosfere. London: Mc Millans, 366 p. [ Links ]
MATIRU, V. N.; DAKORA, F. 2004. Potencial use of rhizobial bacteria as promoters of plant growth for increased yield in landraces of African cereal crops. African Journal of Biotechnology 3: 1–7. [ Links ]
MIRABAL, L; ORTEGA, E.; RODES, R.; FERNÁDEZ, F. 2002. Método efectivo para la desinfección total de esporas de hongos micorrizógenos arbustivos (HMA): Aislamiento y caracterización de bacterias endospóricas en Glomus clarum. Cultivos Tropicales 23(1): 21–24. [ Links ]
MORELL, F.; HERNÁNDEZ, A.; FERNÁNDEZ, F.; TOLEDO, Y. 2006, Caracterización agrobiológica de los suelos ferralíticos rojos lixiviados de la región de San José de las Lajas, en relación con el cambio en el manejo agrícola. Cultivos Tropicales 27(4): 13–18. [ Links ]
PHYLLIPS, D. M.; HAYMAN, D. S. 1970. Improved procedures for clearing roots and staining parasites and vesicular arbuscular mycorrizal fungi for rapid assessment for infection. Trans. British Mycol. Soc. (Londres) 55: 101–188. [ Links ]
RIERA, M.; MEDINA, N. 2005. Influencia de las micorrizas sobre las poblaciones bacterianas y su efecto sobre los rendimientos en secuencias de cultivo. Cultivos Tropicales 6(4): 21–27. [ Links ]
RIVERA, R.; FERNÁNDEZ, K. 2003. Bases Científico–Técnicas para el manejo de los sistemas agrícolas micorrizados eficientemente. In: RIVERA R.; FERNÁNDEZ F.; HERNÁNDEZ, A.; MARTÍN, J. R.; FERNÁNDEZ, K. El manejo efectivo de la simbiosis micorrízica, una vía hacia la agricultura sostenible. Estudio de caso: El Caribe. Editorial Agustín García Marrero. La Habana. 166 p. [ Links ]
RIVES, N.; ACEBO, Y.; HERNÁNDEZ, A. 2007. Bacterias promotoras del crecimiento vegetal en el cultivo del arroz (Oryza sativa L.). Perspectivas de su uso en Cuba. Cultivos Tropicales 28 (2): 29–38. [ Links ]
SIEVERDING, E. 1991. Vesicular Arbuscular Mycorrhiza in Tropical Agrosystem. Deutsch Gesellsschaft fur techniische (GTZ) GMBH, Federal Republic of Germany. 371p. [ Links ]
SOMERS, E.; VANDERLEYDEN, J. 2004. Rhizosphere bacterial signalling; a love parade beneath our feet. Critical reviews of Microbiology 30: 205–240. [ Links ]
TERRY, E.; LEYVA, A. 2006. Evaluación agrobiológica de la inoculación micorrizas–rizobacterias en tomate. Agronomía Costarricense 30(1): 65–73. [ Links ]
WAMBERG, C.; CHRISTENSEN, S.; JAKOBSEN, I.; MULLER, A. K.; SOROSEN, S. J. 2003. The mycorrhizal fungus (Glomus intraradices) affects microbial activity in the rhizosphere of pea plants (Pisum sativum). Soil Biology and Biochemestry 35: 1349–1357. [ Links ]
WANG, H.; PARENT, S.; GOSELIN, A.; DESJARDINS, Y. 1993. Vesicular arbuscular mycorrizal peat–based susbtrates enhance symbiosis establishment and growth of three micropropagated. Journal of America Society 118: 6, 896–901. [ Links ]

