



![Almacenamiento de frutos de zapote mamey [Pouteria sapota (Jacq.) H. E. Moore & Stearn] en atmósfera modificada](/img/es/prev.gif)






Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Chapingo. Serie horticultura
versión On-line ISSN 2007-4034versión impresa ISSN 1027-152X
Rev. Chapingo Ser.Hortic vol.15 no.1 Chapingo ene./abr. 2009
Influencia de la radiación solar en la producción de semilla–tubérculo de papa bajo cultivo sin suelo
Effect of solar radiation in the production of seed potato in soiless cultivation
R. Flores–López*, F. Sánchez–del Castillo, J. E. Rodríguez–Pérez, R. Mora–Aguilar, M. T. Colinas–León y H. Lozoya–Saldaña
Instituto de Horticultura. Departamento de Fitotecnia. Universidad Autónoma Chapingo. Km. 38.5 Carretera México–Texcoco. Chapingo, Estado de México. C. P. 56230. México. Correo–e: floresrmx@yahoo.com.mx (*Autor responsable).
Recibido: 24 de marzo, 2008
Aceptado: 2 de septiembre, 2008
Resumen
El propósito del presente estudio fue determinar la influencia de cuatro niveles de radiación solar (100, 70, 50 y 40%) en invernadero, controlada mediante el uso de mallas plásticas (30, 50 y 60% de sombra y un testigo sin malla), sobre el rendimiento de papa variedad Gigant, cultivada con tubérculos de 15 a 17 mm en perlita. A los 75 días después de la emergencia (DDE) las plantas expuestas a 100 y 70% de radiación tuvieron mayor número (25 a 35%) y cantidad de biomasa de tubérculos (50%), biomasa total, fotosíntesis neta, tasa absoluta de crecimiento, tasa de asimilación neta y tasa de asimilación económica. El índice de cosecha fluctuó entre 0.51 y 0.71 en las plantas cultivadas bajo 40 y 100% de radiación, respectivamente. En términos generales, este comportamiento fue también observado en muestreos a los 40, 50, 60 y 70 DDE, es decir, a menor radiación se disminuyó la eficiencia fotosintética de la planta, lo que repercutió directamente en la disminución de biomasa del vástago y tubérculo; así como en los índices de eficiencia medidos mediante análisis de crecimiento.
Palabras clave: Solanum tuberosum, tubérculo–semilla, invernadero, sustrato, hidroponía.
Abstract
The aim of this research was to determinate the effect of four–radiation levels (100, 70, 50 and 0%) provided by plastic mesh (30, 50 and 60% shade, and a control without mesh) on tuber biomass accumulation in exposed greenhouse plants. Giant variety tubers 15 to 17 mm in diameter were grown in perlite. Seventy–five days after emergence plants under 100 and 70% radiation had a higher number of tubers tuber (25 a 35%) and total and tuber biomass (50%), net photosynthesis, absolute growth rate, net assimilation rate and net economic rate than those grown under 50 and 40% radiation. The harvest index was 0.71 at 100% vs. 0.51 at 40% radiation. This general behavior was observed in samples 40, 50, 60 and 70 days after emergence. Lower radiation affects photosynthetic efficiency of potato plants, decreasing tuber and total biomass, as well as the efficiency indexes of the growth analysis.
Key words: Solanum tuberosum L, hydroponics, seed–tuber, substrate, greenhouse.
INTRODUCCIÓN
El cultivo de papa (Solanum tuberosum L.) en México está limitado por la poca disponibilidad de semilla certificada; sólo 13% (8,000 ha) de 62,000 ha cultivadas con este tubérculo es sembrada con ésta; no obstante que existe la infraestructura necesaria de laboratorios e invernaderos para su producción. El proceso de producción de semilla certificada en México incluye las fases de laboratorio (semilla prebásica I), invernadero (básica II) y cinco ciclos en campo (NOM–041–FITO–2002, Diario Oficial de la Federación, 2003).
En la producción de tubérculo–semilla de papa en invernadero se emplean diversas cubiertas plásticas que difieren en la transmisión de radiación: desde 50 hasta 85% de la radiación total incidente, situación que repercute en rendimientos variables por planta, por lo que es necesario determinar el nivel de radiación óptimo para la producción de minitubérculos, hacer más eficiente este sistema de producción e incrementar la disponibilidad de semilla básica.
El proceso de tuberización en papa es influenciado por varios factores: como el balance hormonal (Struick y Wiersema, 1999), concentración interna de Ag3 (Vreugdenhil y Sergeeva, 1999), fotoperíodo (Martínez–García et al., 2002), radiación (Jackson, 1999), concentración de CO2 (Lawson et al., 2002), nutrición mineral (Struik y Wiersema, 1999), temperatura (Thornton et al., 1996) y del genotipo (Struik y Wiersema, 1999).
Más del 80% del peso seco de una planta se origina del carbono fijado por la fotosíntesis; sin embargo, no se puede asegurar que a mayor intercepción de radiación solar, mayor sea la producción del órgano de interés debido a que la hoja de papa se satura a 1,200 ì mol·m–2·s–1 (Dwelle, 1985). La radiación necesaria para saturar las hojas de una planta completa en un dosel debe ser más alta, por lo que, tanto el tipo de cubierta, como la densidad de plantación son importantes en el rendimiento del cultivo bajo invernadero. En cultivo in vitro, Niu et al., (1996) mostraron que al incrementar la irradiación aumentó la masa seca. En condiciones controladas, bajo sombra, decreció sustancialmente el número de células del tubérculo tanto al inicio de la tuberización como en la etapa de llenado, lo que afectó el rendimiento de la planta (Chen y Setter, 2003).
El empleo de indicadores fisiológicos, como la tasa absoluta de crecimiento (TAC), tasa relativa de crecimiento (TRC), tasa de asimilación neta (TAN), razón de área foliar (RAF), índice de área foliar (IAF) e índice de cosecha, han sido empleados para entender el efecto de diversos factores bióticos y abióticos sobre el cultivo de la papa (McCollum, 1978; Mora et al., 2006), en la evaluación de genotipos (Aguilar et al., 2006), sobre el efecto de diferentes condiciones de humedad del suelo y determinación de la eficiencia de fósforo (McCollum, 1978).
Por otro lado, el cultivo hidropónico, ofrece como principal ventaja la disposición continua de nutrientes a la planta y se emplean diversos sustratos como: turba, fibra de coco, perlita, mezclas físicas de turba o fibra de coco con perlita (Struik y Wiersema, 1999; Ritter et al., 2001).
El propósito de la presente investigación fue determinar la influencia de cuatro niveles de radiación, provocados por malla de sombra, sobre la tuberización en plantas de papa cultivadas en invernadero y en hidroponia, en términos de acumulación de biomasa y número de tubérculos para su uso como semilla en campo.
MATERIALES Y MÉTODOS
El experimento se estableció en Chapingo, Estado de México, de mayo a agosto de 2006 en un invernadero de vidrio; las condiciones ambientales dentro del invernadero fueron: humedad relativa media 62%, máxima 88% y mínima 36%; temperatura media diaria 21 °C, mínima nocturna 10 °C y máxima diurna 35 °C, la cual fue tomada cada media hora con dos registradores de datos Hanna. La radiación incidente al interior del invernadero varió a lo largo del día, de acuerdo con la época del año, a la hora del día y el grado de nubosidad presente, registrándose una lectura máxima de hasta 2000 ì moles·m–2·s–1 en la condición de 100% de radiación.
Se empleó la variedad Gigant, liberada en Holanda en 1987, precoz (80 a 90 días), piel blanca, ojos superficiales, pulpa crema, forma oblonga–aplanada, susceptible al tizón tardío de la papa (Phytophthora infestans Mont. De Bary). Actualmente se cultiva en México y es una de las principales variedades para consumo en fresco (Rubio et al., 2000). Los minitubérculos utilizados fueron de 15 a 17 mm de diámetro, libres de enfermedades de virus, hongos y bacterias, provenientes de plántulas in vitro, desarrolladas en invernadero con certificación para la producción de semilla–tubérculo de papa.
Los tratamientos aplicados consistieron en cuatro niveles de radiación incidente: 100, 70, 50 y 40%, proporcionados mediante el uso de mallas plásticas negras con diferente densidad de filamentos, estos porcentajes de sombra fueron determinados con mediciones hechas por debajo y por encima de cada una de las mallas, con un ceptómetro lineal (PAR Sunfleck, Decagon Devices). Al nivel de 100% no se aplicó sombra y se consideró como testigo. El experimento se realizó en condiciones hidropónicas en bolsa de polietileno bicolor (negro por dentro y blanco por fuera) con 25 litros de perlita grado hortícola (1 a 4 mm de diámetro); cada bolsa se sembró con un minitubérculo a 12 cm de profundidad. La unidad experimental consistió de 11 macetas con separación de 50 cm entre ellas, lo que correspondió a una densidad de 4 plantas·m–2 útil. Se utilizó un diseño experimental de bloques completos al azar, con cuatro repeticiones. Las plantas se condujeron a un tallo y la solución nutritiva empleada tuvo la siguiente concentración de nutrimentos en mg·litro–1: N 160, P 80, K 250, Ca 250, Mg 75, Fe 4, Mn 0.5, Cu 0.5, Zn 0.5, B 0.5, con pH de 6.0 y conductividad eléctrica final de 2.0 dS·m–1. El gasto diario de la solución nutritiva suministrada mediante riego por goteo fue en promedio de 500 ml por planta, distribuida en cuatro riegos (9:30, 12:30, 14:30 y 16:30 h).
Se hicieron aplicaciones preventivas con Mancozeb a razón de 2 g·litro–1 y Clorotalonil 1.5 ml·litro–1 para el control de Phytophthora infestans, así mismo, se alternaron aplicaciones de Metamidofos, Endosulfan y Ometoato una vez por semana, para el control de mosquita blanca (Bemisia tabaci Genn.).
Se hicieron muestreos destructivos de cuatro plantas por tratamiento (una por repetición) a los 30, 40, 50, 60, y 75 días después de la emergencia, la planta se separó en sus diferentes órganos y se contaron número de tubérculos mayores de 10 mm, altura de la planta (cm) y área foliar (cm2); esta última medida con un integrador LI–COR, modelo LI–3100. El peso seco de estolones (g), tubérculos (g), raíz (g), hojas y tallos (g), se obtuvo al secar en estufa a 70 °C durante 72 a 96 h de acuerdo al tipo de órgano.
En la quinta hoja a partir del ápice, que se encontraba totalmente expandida se midió la fotosíntesis neta en cuatro ocasiones a los 40, 50, 60 y 70 DDE con un analizador de gases de infrarrojo marca LI–COR, modelo LI–6250.
El análisis de crecimiento se realizó con los datos de masa seca de tubérculo, vástago y planta completa, los cuales fueron ajustados mediante regresión no lineal, con el
modelo logístico:  donde: Y fue la variable respuesta; x la variable independiente; A la asintota, valor de y cuando x tiende a infinito; B el parámetro relacionado con la ordenada al origen; C el valor de x para el punto de inflexión, velocidad de crecimiento; e= 2.718281828. Posteriormente se calculó la tasa absoluta de crecimiento (TAC), que corresponde a la primera derivada del modelo
donde: Y fue la variable respuesta; x la variable independiente; A la asintota, valor de y cuando x tiende a infinito; B el parámetro relacionado con la ordenada al origen; C el valor de x para el punto de inflexión, velocidad de crecimiento; e= 2.718281828. Posteriormente se calculó la tasa absoluta de crecimiento (TAC), que corresponde a la primera derivada del modelo
logístico: (Rodríguez et al., 2006), y 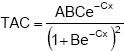 que se define como la ganancia en gramos de materia seca por día. En este caso del vástago, tubérculos y de la planta completa.
que se define como la ganancia en gramos de materia seca por día. En este caso del vástago, tubérculos y de la planta completa.
La tasa de asimilación neta (TAN) que representa la cantidad de biomasa acumulada por unidad de área foliar y unidad de tiempo (g·m–2·d–1), se calculó de acuerdo con la fórmula siguiente:
 donde: TAC fue la tasa absoluta de crecimiento; af el área foliar.
donde: TAC fue la tasa absoluta de crecimiento; af el área foliar.
La tasa de asimilación económica neta (TAEN) representa la cantidad de biomasa del órgano de interés por unidad de área foliar y unidad de tiempo (g·m–2·d–1). Es análoga a la TAN, sólo que se toma en cuenta el peso seco de tubérculo (McCollum, 1978). El índice de cosecha (IC) se obtuvo al dividir la biomasa de los tubérculos entre la biomasa total acumulada por la planta hasta el momento de la cosecha.
Se efectuó la prueba de correlación de Pearson, el análisis de varianza correspondiente al diseño experimental empleado y las pruebas de comparación de medias de Tukey en cada variable en estudio en cada muestreo. Se utilizó el programa Statistical Analysis System (SAS) versión ocho.
RESULTADOS Y DISCUSIÓN
La altura de las plantas desde los 50 DDE fue significativamente mayor en los tratamientos donde disminuyó la radiación incidente (Cuadro 1). A los 75 DDE las plantas del tratamiento testigo sin sombra tuvieron en promedio 33 cm menos altura que plantas sometidas a 60% de reducción de la radiación incidente, esto pudo deberse a que la baja intensidad de luz provocó la elongación del tallo por el alargamiento de las células, posiblemente por mayor acumulación de auxinas (Gardner et al., 1985), aunque también se han demostrado incrementos en los niveles de giberelinas en hojas bajo sombra (Woolley y Waering, 1972).

El área foliar por planta fue igual en todos los tratamientos de luz (P≤0.05) a los 50 y 60 DDE, pero a los 75 DDE aumentó significativamente en plantas cultivadas bajo 40% de radiación con respecto a los demás tratamientos que fueron estadísticamente iguales. Los tratamientos de 70 y 50% de radiación presentaron área foliar intermedia entre el testigo y 40%. El incremento del área foliar con menor disposición de radiación posiblemente es debido a que las plantas de papa en esas condiciones requieren superar las deficiencias en fotosíntesis y destinan mayor proporción de los carbohidratos que producen hacia la formación de hojas (Gawronska et al., 1990).
El crecimiento de la raíz tuvo fuerte influencia de la radiación, ya que la acumulación de masa seca de este órgano en plantas con 100 y 70% de radiación fue superior (P≤0.05) al mostrado con 50 y 40% desde los 50 DDE (Cuadro 1). El mayor crecimiento de la raíz favoreció a la planta al explorar más área del sustrato y tener mayor disponibilidad de nutrientes, lo que también pudo influir en el rendimiento biológico y económico de las plantas.
En el Cuadro 2 se presenta la fotosíntesis neta, medida en los diferentes muestreos. Las plantas expuestas a menor radiación tuvieron tasas fotosintéticas de 30 a 50% de las que alcanzaron las plantas expuestas a 100 y 70% de radiación; sin embargo, sólo se detectaron diferencias significativas (P≤0.05) en los muestreos de los 50, 60 y 70 DDE.
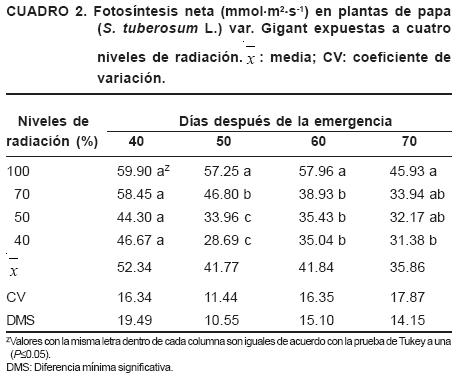
Contrario a lo que indica Dwelle, (1985) la fotosíntesis neta de la hoja no se incrementó durante el llenado de tubérculos. Las plantas del tratamiento de 40% de radiación presentaron una disminución significativa de la tasa de fotosíntesis neta entre 50 a 70 DDE con respecto al testigo.
También se observa que a los 70 DDE bajó la fotosíntesis neta en las plantas de todos los tratamientos, posiblemente debido a la senescencia general de las hojas.
Los valores de acumulación de masa seca de tubérculo, vástago y total fueron linearizados mediante el modelo logístico, los parámetros A, B y C (Cuadro 3) fueron empleados para el cálculo de los parámetros del análisis de crecimiento, TAC, TAN y TAEN, que se discuten a continuación.
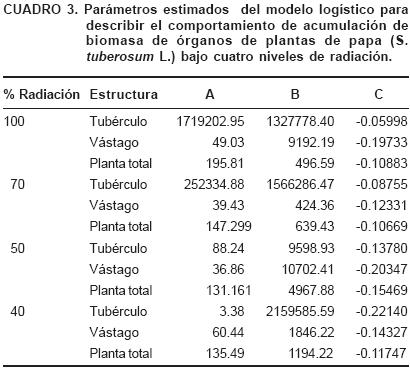
La Figura 1 presenta la acumulación de biomasa y tasas absolutas de crecimiento (TAC) de vástago, tubérculos y de la planta completa. En el caso de tubérculos la TAC se incrementó gradualmente de los 40 a los 60 DDE y de manera exponencial de los 60 hasta los 75 DDE en condiciones de 100 y 70% de radiación, con valores de 6.98 y 10.01 g·d–1 respectivamente a los 75 DDE, en contraste con plantas de los tratamientos de 50 y 40% con valores de 2.2 y 2.1 g·d–1, respectivamente, siendo de tres veces a cinco mayor.
La TAC del vástago se incrementó de los 30 a los 50 DDE disminuyendo posteriormente, lo que indica que la planta, en este período, destinó carbohidratos tanto a los tubérculos como a la formación de hojas y tallos; posteriormente disminuyó la acumulación de asimilados en la formación del vástago y se incrementó su acumulación en tubérculos. Lo que sugiere que las plantas en condiciones de baja luminosidad invierten mayor proporción de asimilados en la generación de hojas y crecimiento del tallo en detrimento del llenado de los tubérculos (Gawronska et al., 1990).
Sin embargo, la TAC de la planta completa indica que entre los 60 y 75 DDE la ganancia de masa seca por día fue menor que la ganancia en tubérculos, lo que sugiere, que la mayor acumulación de biomasa después de los 70 días se debería a la remobilización de carbohidratos de otras estructuras de la planta (hojas, tallo) hacia los tubérculos, lo que ocasiona la senescencia precoz del follaje (Mora et al., 2006).
Lo anterior puede confirmase al atender el comportamiento de la acumulación de masa seca (Figura 1) donde se tiene un incrementó sobresaliente a los 70 a los 75 DDE en tubérculos. Así mismo, se aprecia que el comportamiento de acumulación de materia seca total está determinado por la acumulación de ésta en tubérculos. Las plantas de los tratamientos de 50 y 40% de radiación tuvieron un comportamiento similar con menor acumulación de biomasa durante todo el ciclo de cultivo. Lo anterior se explica al observar que hay menor fotosíntesis neta en plantas con tratamientos en menor radiación (Cuadro 2), lo que condujo a menor acumulación de biomasa causado por la menor intercepción de radiación (Gawronska et al., 1990).
En la Figura 2 se presenta la tasa de asimilación neta, las plantas en 100% de radiación mostraron valores altos durante todo el ciclo que a menor radiación, con la TAN superior a los 50 a 55 DDE de 10.05 y 6.49 g·m–2·d–1 para el testigo y 70% de radiación, respectivamente y de 8.30 y 5.64 g·m–2·d–1 para 50 y 40%, lo que sugiere mayor eficiencia fotosintética por unidad de superficie foliar a mayor radiación, en comparación con plantas expuestas a menor radiación, no obstante tener una área foliar igual o mayor (Cuadro 1). A 40% de radiación las plantas produjeron un 55% menos de materia seca por unidad de superficie foliar por día a los 50 DDE y un 48 al 50% menos a los 60, 70 y 75 DDE que el testigo. También se aprecia que la TAN después de los 40 días disminuyó en todos los tratamientos debido posiblemente al autosombreo de las hojas al aumentar el follaje.
La tasa de asimilación económica neta fue mayor en el testigo que los tratamientos de sombra (Figura 2), lo que indica que la planta destina mayor cantidad de asimilados al tubérculo en condiciones de alta radiación por unidad de área foliar y tiempo.
Al usar este parámetro junto con la TAN se puede aseverar que en condiciones de alta radiación no sólo se produce mayor biomasa total sino también hay mayor partición de asimilados al tubérculo, por lo que la TAEN puede ser usada como un coeficiente de partición (McCollum, 1978).
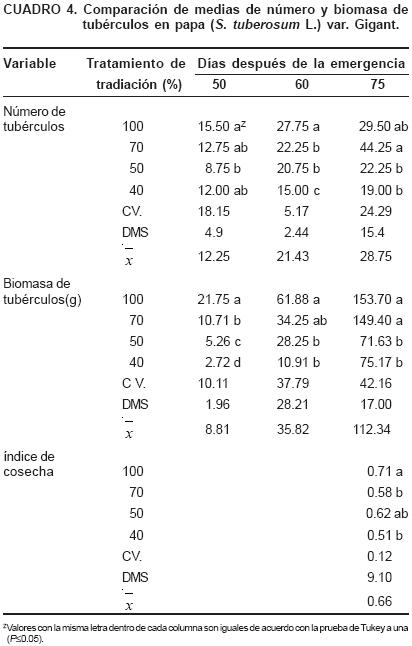
Se obtuvo mayor rendimiento en peso seco de tubérculos en la condición de 100% de radiación (Cuadro 4), Similarmente el número de tubérculos a los 75 DDE fue mayor (P≤0.05) en los tratamientos que recibieron más radiación con ausencia de diferencia estadística entre los tratamientos de 100 y 70% de radiación en relación al rendimiento y número de tubérculos, lo que sugiere que en localidades con radiación solar similar a la de Chapingo, México, aún con 30% de sombra es posible producir minitubérculos de papa sin disminución significativa en el rendimiento. Es decir, las plantas sometidas a mayor radiación destinaron mayor cantidad y proporción de asimilados al llenado de tubérculos con un índice de cosecha a los 75 días de 0.71 contra 0.51 en el tratamiento de 40% de radiación, donde no sólo hay una reducción en la biomasa total de la planta, sino también en el índice de cosecha (Cuadro 4), lo anterior coincide con lo mencionado por Gawronska et al. (1990) que plantas de papa en alta intensidad de luz invierten del 60 al 74% de la biomasa total hacia los tubérculos.
CONCLUSIONES
En las condiciones en las que se realizó el estudio se puede concluir que reducir la radiación solar incidente en el invernadero hasta un 30% no repercute en una disminución significativa de biomasa total y en la partición de asimilados al órgano de interés, lo que ocurre si la tasa de sombra es superior del 50%.
LITERATURA CITADA
AGUILAR, L. M. G.; CARRILLO, S. A. J.; RIVERA, P. A.; GONZÁLEZ, H. A. V. 2006. Análisis de crecimiento y de relaciones fuente–demanda en dos variedades de papa (Solanum tuberosum L.). Revista Fitotecnia Mexicana 29(2): 145–156. [ Links ]
CHEN, C. T.; SETTER, T. L. 2003. Response of potato tuber cell division and growth to shade and elevated CO2. Annals of Botany 91(3): 373–381. [ Links ]
DWELLE, B. R. 1985. Photosynthesis and photoassimilate partitioning. pp. 35–58. In: Li, H. P. (Ed), Potato Physiology. Academic Press, INC. Orlando, Florida. [ Links ]
GARDNER, P. F.; PEARCE, B. R.; MITCHEL, L. R. 1989. Physiology of Crop Plants. Iowa State University Press: AMES. USA. [ Links ]
GAWRONSKA, H.; DWELLE, B. R.; PAVEK, J. J. 1990. Partitioning of photoassimilates by potato plants (Solanum tuberosum L.) as influenced by irradiance: II. Partitioning patterns by four clones grown under high and low irradiation. American Potato Journal 67: 163–176. [ Links ]
JACKSON, D. S. 1999. Multiple signaling pathways control tuber induction in potato. Plant Physiology 119: 1–8. [ Links ]
LAWSON, T.; CRAIGON, J.; BLACK, R. C.; COLLS, J. J.; LANDON, G.; WEYERS, B. D. 2002. Impact of elevated CO2 and O3 on gas exchange parameters and epidermal characteristics in potato (Solanum tuberosum L.). Journal of Experimental Botany 53(369): 737–746. [ Links ]
MARTÍNEZ–GARCÍA, F. J.; GARCÍA–MARTÍNEZ, L. J.; BOU, J.; PRAT, S. 2002. The interaction of gibberellins and photoperiod in the control of potato tuberization. J. Plant Growth Regulation. 20: 377–386. [ Links ]
McCOLLUM, E. R. 1978. Analysis of potato growth under differing P regimes. II. Time by P–status interactions for growth and leaf efficiency. Agronomy Journal 70: 58–67. [ Links ]
MORA, A. R.; ORTIZ, C. J.; RIVERA, P. A.; MENDOZA, C. C. A.; COLINAS, L. M. T.; LOZOYA, S. H. 2006. Índices de eficiencia de genotipos de papa establecidos en condiciones de secano. Revista Chapingo Serie Horticultura 12(1): 85–94. [ Links ]
NIU, G.; KOZAI, T.; MIKAMI, H. 1996. Simulation of the effects of photoperiod and light intensitiy on the growth of potato plantlets cultured photoautotrophically in vitro. Acta Horticulturae 440: 622–627. [ Links ]
RITTER, E.; ANGULO, B.; RIGA, P.; HERRÁN, C.; RELLOSO, J.; JOSÉ, S. M. 2001. Comparison of hydroponic and aeroponic cultivation systems for the production of potato minitubers. Potato Research 44: 127–135. [ Links ]
RODRÍGUEZ, P. J. E.; SAHAGÚN, C. R.; GASGA, P.; MORA, A. R. 2006. Programas de cómputo para la estimación de parámetros de regresión no lineal útiles en el análisis de crecimiento de cultivos. 52da Reunión anual de la Sociedad Interamericana de Horticultura Tropical. Sep. 24–30. San Juan, Puerto Rico. [ Links ]
RUBIO, C. O. A.; RANGEL, G. J.; FLORES, L. R.; MAGALLANES, G. J. V.; DÍAZ, H. C.; ZAVALA, Q. T.; RIVERA, P. A.; CADENA, H. M.; ROCHA, R. R.; ORTÍZ, T. C.; LÓPEZ, D. H.; DÍAZ, V. M.; PAREDES, T. A. 2000. Manual para la producción de papa en las sierras y valles altos del centro de México. SAGAR–INIFAP–CIRCE. Campo Experimental Valle de Toluca. Libro Técnico Núm. 1. Zinacantepec, Estado de México.79. [ Links ]
STRUIK, C. P.; WIERSEMA, G. S. 1999. Seed Potato Technology. Ed. Wageningen Pers, Wageningen, The Netherlands 383. [ Links ]
THORNTON, K. M.; MALIK, J. N.; DWELLE, B. R. 1996. Relationship between leaf gas exchange characteristics and productivity of potato clones grown at different temperatures. American Potato Journal 73: 63–77. [ Links ]
VREUGDENHIL, D.; SERGEEVA, I. L. 1999. Gibberellins and tuberization in potato. Potato Research 42: 471–481. [ Links ]
WOOLEY D. J.; WAEREING, P. F. 1972. Environmental effects on endogenous cytokinins and gibberellin levels in Solanum tuberosum. New Phytology 71: 1015–1025. [ Links ]

